亚细胞特定区域化线粒体含量是否与肌纤维类型、饮食干预有关?
2021-06-02耿国强梁祎珩RasmusJensenJoachimNielsenNielsrtenblad邱俊强
耿国强,梁祎珩,Rasmus Jensen,Joachim Nielsen,Niels Ørtenblad,邱俊强*
(1.北京体育大学,北京 100084;2.南丹麦大学,丹麦欧登塞,5230)
线粒体作为能为机体供给能源物质并支持人体运动功能的重要细胞器,在体育科学界始终受到广泛的关注。有研究表明,线粒体含量是影响线粒体功能的重要因素(Groennebaek et al.,2020)。在电镜观察下,基于肌纤维肌节的结构特性,线粒体在骨骼肌中并非均匀分布(Boncompagni et al.,2009;Elander et al.,1985;Nielsen et al.,2017)。根据所分布的不同区域划分线粒体亚型的概念最早被Palmer等(1977)提出,他们从大鼠心肌中提取出两种不同类型的线粒体,并证明了肌膜下线粒体对耐力训练具有一定的适应性。在随后的研究中,根据结构和形态学上的差异,将位于肌细胞中不同特定区域的线粒体,称为亚细胞特定区域化线粒体,有两种类型,分别为肌原纤维间线粒体(intermyofibrillar mitochondria,IMF线粒体)和肌膜下线粒体(subsarcolemmal mitochondria,SS线粒体)(Krieger et al.,1980;Takahashi et al.,1996)。近年来,大量研究证实,肌细胞内的线粒体不均匀分布具备功能表达特异性,例如,SS和IMF线粒体对蛋白质合成和代谢水平的作用有明显差异,且通过不同的通路进行调控(Cogswell et al.,1993;Ferreira et al.,2010;Groennebaek et al.,2020)。但是,关于人体肌细胞特定区域线粒体的含量及其在快慢肌中分布特性的研究较少。虽然线粒体总含量的多少是区分Ⅰ/Ⅱ型肌纤维的直接证据之一(Ogata et al.,1985),但是使用光学显微镜量化线粒体含量以判定肌纤维类型的方法,实用性不强且时间和经济成本较高。有研究发现,肌纤维的z线宽度与肌纤维类型有较强的关联性,因此认为其可能可以作为电子显微镜下有效预测肌纤维类型的指标之一(Sjöström et al.,1982)。但是,时至今日仍然没有足够的数据证明z线宽度和特定区域线粒体含量的直接相关关系,因此并未被广泛使用。
短期饮食干预可影响耐力运动表现,相比混合饮食或高脂低碳水饮食干预,高糖膳食后具有更高肌糖原储备的受试者进行耐力运动的力竭时间明显更长(Bergström et al.,1967;Jeukendrup,2011),提示肌糖原含量与运动表现密切相关。作为肌糖原有氧氧化的主要代谢场所,线粒体被认为是氧化磷酸化供能过程中最重要的细胞器之一。与线粒体在骨骼肌中的分布特点类似,研究已证实肌糖原在骨骼肌中也符合非均匀分布,即分布具有区域化特性,依据肌节结构特性将肌糖原储存于独立的糖原“贮存池”(Han et al.,1992)。有研究在静坐少动人群中证实了这一观点,结果表明,SS肌糖原的储存敏感度和优先级都高于IMF区域肌糖原,糖原的储存存在区域差异性(Nielsen et al.,2009)。也有研究报道了短期饮食干预的肌糖原含量储存具有肌纤维类型和亚细胞区域化特异性,分布于肌纤维内部的糖原储备含量和中高强度耐力运动表现有潜在的相关性(Fisher,2020)。随着人体有氧运动时间的累积,线粒体作为糖原有氧氧化的主要代谢场所,在肌糖原非线性的氧化供能过程中起到重要的调控作用(Irrcher et al.,2003)。而以耐力运动中能源底物(糖原)与线粒体代谢为切入点,大量研究报道了能量代谢过程中的线粒体区域化特性,Chomentowski等(2011)的研究表明,两种不同区域化分布的线粒体具有各自独立的线粒体调控模式,IMF线粒体含量受胰岛素活性的直接影响,而SS线粒体含量却保持不变。还有研究报道,骨骼肌细胞中特定区域化线粒体的呼吸作用有明显的差异,IMF线粒体第Ⅲ阶段的呼吸作用水平是SS线粒体的2.3~2.8倍(Adhihetty et al.,2005)。然而,关于短期饮食干预对肌糖原的影响是否进而对相应区域的线粒体含量产生影响则尚未可知。因此,本研究在不同饮食干预下,尝试探究不同运动表现的差异是否与骨骼肌线粒体含量的区域化分布特性以及不同肌纤维类型有关,为特定区域化线粒体特性的后续研究提供更多的参考依据。
1 研究对象与方法
1.1 研究对象
招募11名无专业运动经历的男性运动爱好者作为研究对象(基本情况:年龄25±4岁,身高183±6 cm,体重77±10 kg,BMI23±2 kg/m2,体脂率14±3%,最大摄氧量4.54±0.58 L/min)。实验前,由实验人员告知受试者实验过程中可能存在的风险、益处以及详细实验流程和时间进度,受试者知悉实验信息后签署了知情同意书。本研究得到南丹麦健康研究伦理区域委员会(S-20170198)的批准,并符合《赫尔辛基宣言》中所述的标准。
1.2 运动测试方案
在实验正式开始两周前,对所有受试者进行基线测试。使用DXA双能射线骨密度测试仪(Lunar Prodigy,GE Healthcare,丹麦邦比)对受试者体成分进行测量。使用功率车递增运动负荷方案测定受试者的最大摄氧量(O2max),使用间接测热法通过气体代谢仪(AMIS Sport,Innovision Aps,Glamsbjerg,丹麦)采集气体代谢数据,实验前使用16.0%O2和4.0%CO2的高精度两组分气体混合物进行校准,并使用3 L空气注射器在低、中、高流速下对流量计(Hans Rudolph,Kansas美国密苏里州市)进行校准。随后依据O2max测试的结果,确定受试者在测试周进行正式实验的运动强度,为75%O2max功率自行车强度(Hvid et al.,2013;Loon et al.,2004)。
正式实验包括3次耐力力竭测试,每2次测试需间隔3天,间隔期间进行不同的短期饮食干预。受试者在实验室完成75%O2max恒定强度的功率自行车力竭测试。恒定负荷运动期间,当受试者的运动强度在5 min内连续两次低于既定强度(75%O2max强度)的90%,或者未能在30 s内恢复到既定运动强度时,则认为受试者达到力竭无法继续运动,即刻停止实验并进行肌肉活检。
在每次耐力力竭测试开始前,实验人员需在每位受试者下肢的股外侧肌部位采集肌肉活检样本,随后开始运动,运动中每隔60 min在相同的部位取肌肉活检,直至力竭,力竭后即刻再取一次肌肉活检。由于3种不同饮食干预下的测试平均力竭时间并不一致,因此,3次测试(依次为混合饮食干预、高脂肪+高蛋白质饮食干预和高碳水化合物饮食干预)分别采集3条、2条、4条肌肉纤维,每个受试者共取9个肌肉活检位点。
1.3 饮食干预方案
为了探究不同的短期饮食干预是否会影响骨骼肌线粒体含量,本研究采用了由Bergström等(1967)设计的测试方案,该方案被认为是经典的饮食和耐力运动干预方案,所有受试者在完成3次不同的短期饮食干预(3天)后进行耐力力竭测试(图1)。在实验周开始前,所有受试者可进行自选混合饮食(self-chosen mix-diet),在第一个测试日前一天(周一)受试者进行为期一天的给定混合饮食干预(1 400 kcal碳水化合物+850 kcal蛋白质+550 kcal脂肪),并于周二进行耐力力竭测试。在第1次测试结束后,饮食干预方案为3天高脂肪+高蛋白质饮食(1 500 kcal脂肪+1 300 kcal蛋白质),直至第2测试日(周五)。测试结束后饮食干预方案为3天高碳水化合物饮食(2 300 kcal碳水化合物+500 kcal蛋白质),直至第3测试日(下周一)。上述饮食方案除自由混合饮食外,其他所有方案均由实验人员提供所需的食材和食谱,且受试者须严格按照食谱要求在家烹饪和进食。
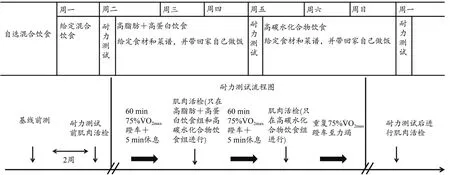
图1 实验流程图Figure 1.Experimental Protocol
1.4 肌纤维类型挑选方法
活检的每条肌肉中提取10条肌纤维,并以肌纤维的z线宽度为肌纤维类型的鉴别指标。为了定义两种主要的肌纤维类型(Ⅰ型肌纤维和Ⅱ型肌纤维),只挑选出明显的Ⅰ型和Ⅱ型肌纤维各3条,为了对比两种类型肌纤维的线粒体含量的明显差异,剔除了Ⅱa型肌纤维的线粒体含量干扰因素,研究人员在10条肌纤维中挑选z线宽度最粗的3条肌纤维并定义为Ⅰ型肌纤维,相反的z线宽度最细的3条被定义为Ⅱ型肌纤维(Nielsen et al.,2010)。由于在测试过程中,有两名受试者分别只接受了8次肌肉活检,实验人员依据上述方法共提取出582条肌纤维用以之后的电镜切片材料。
1.5 电镜切片制作
将肌肉活检取出的肌肉样本用2.5%戊二醛在0.1 M、pH 7.3的二甲胂酸钠缓冲液中固定24 h,随后继续在0.1 M、pH 7.3的二甲胂酸钠缓冲液中冲洗4次。冲洗后,将纤维样本用1%四氧化锇和1.5%亚铁氰化钾在4℃、0.1 M二甲胂酸钠缓冲液中再固定90 min,之后继续在4℃、0.1 M二甲胂酸钠缓冲液中冲洗2次。之后通过一系列分级的4℃~20℃酒精脱水,和20℃氧化丙烯和环氧树脂分级混合物的浸润后,包埋在100%、30℃环氧树脂下。上述工序处理之后,使用切片机(Ultracut UCT超薄切片机,莱卡微观系统,德国)将肌纤维样本切成3个深度(相隔150 μm)的60 nm超薄切片,并与乙酸铀酰和柠檬酸铅对比。最后在预先校准过的CM100透射电子显微镜(飞利浦,荷兰)和Olympus Veleta镜头(Olympus图像软件,德国)下检查切片并拍摄照片。将制备好的切片放大13 500倍,并以随机系统化的原则分别拍摄12张IMF区域的图片和12张SS区域的图片(图2)。

图2 肌纤维电镜切片示意图Figure 2.The Schematic Diagram of Fiber Sections with TSM
1.6 体视学方法
在形态学和线粒体含量的检测阶段为双盲操作。2名研究人员使用标准的体视学方法盲审计算亚细胞特定区域化线粒体体积密度(图3)。2名研究人员对不同组别下电镜切片的分析工作量是相同的。使用点计数法(Gundersen et al.,1988)测量所有线粒体的体积密度(volume density,VV),公式为VV=AA,AA是通过点计数法测量出的线粒体面积分数,点计数网格尺寸为60 nm×60 nm。由于线粒体所在区域不同,IMF线粒体含量表达为相应肌原纤维的体积分数的形式(单位μ3·μ-3),SS线粒体含量表达为线粒体体积除以肌纤维表面积的形式(单位μ3·μ-2);总线粒体的体积密度根据上述两种类型线粒体含量重新计算得出,遵循公式总线粒体(μm3·μm-3)= IMF线粒体(μm3·μm-3)+ SS线粒体(μm3·μm-2)/20,公式中 SS线粒体含量做次分数运算得出相应的体积密度数值,根据前人研究,理想假设肌纤维是半径为40 μm的圆柱体(Nielsen et al.,2010)。

图3 体视学方法标记线粒体示意图Figure 3.The Schematic Diagram of Counting Mitochondria by Using Stereological Method
1.7 统计学方法
使用SPSS 27.0软件进行统计学数据分析,所有数据通过了正态分布和方差齐性检验,采用平均数±标准差的形式呈现。线粒体含量与z线宽度的相关关系采用皮尔逊相关系数的方法处理,皮尔逊相关系数R取值-1到+1,绝对值越大表示相关程度越大。使用单因素方差分析的方法测试3种不同短期饮食干预下耐力力竭测试的运动表现的差异性。使用混合效应模型分析交互作用和主效应来比较肌纤维线粒体含量在不同类型肌纤维和不同短期饮食干预下的差异,其中肌纤维类型和饮食干预类型为固定因素,受试者基础信息作为随机因素。首先确定两个固定变量的交互作用,判定两者是否存在互相之间的影响。若两变量存在交互作用,则通过简单效应检验数据结果;如无交互作用,则直接检验主效应结果。P<0.05被认为有显著性差异。
2 研究结果
2.1 不同饮食干预下耐力测试力竭时间的差异
根据表1可知,3种短期饮食干预下耐力测试力竭时间有显著性差异,高碳水化合物饮食组的力竭时间相比其他两组显著更长(P<0.001)。

表1 3种饮食干预下耐力测试力竭时间Table 1 Exhaustion Time of Endurance Tests under 3 Different Diets
2.2 亚细胞特定区域化线粒体和z线宽度的相关性研究
根据表2可知,IMF、SS和总线粒体含量都和其对应的z线宽度具有显著中等正相关性(P<0.001,RTotal=0.44,RIMF=0.42,RSS=0.37,图 4)。
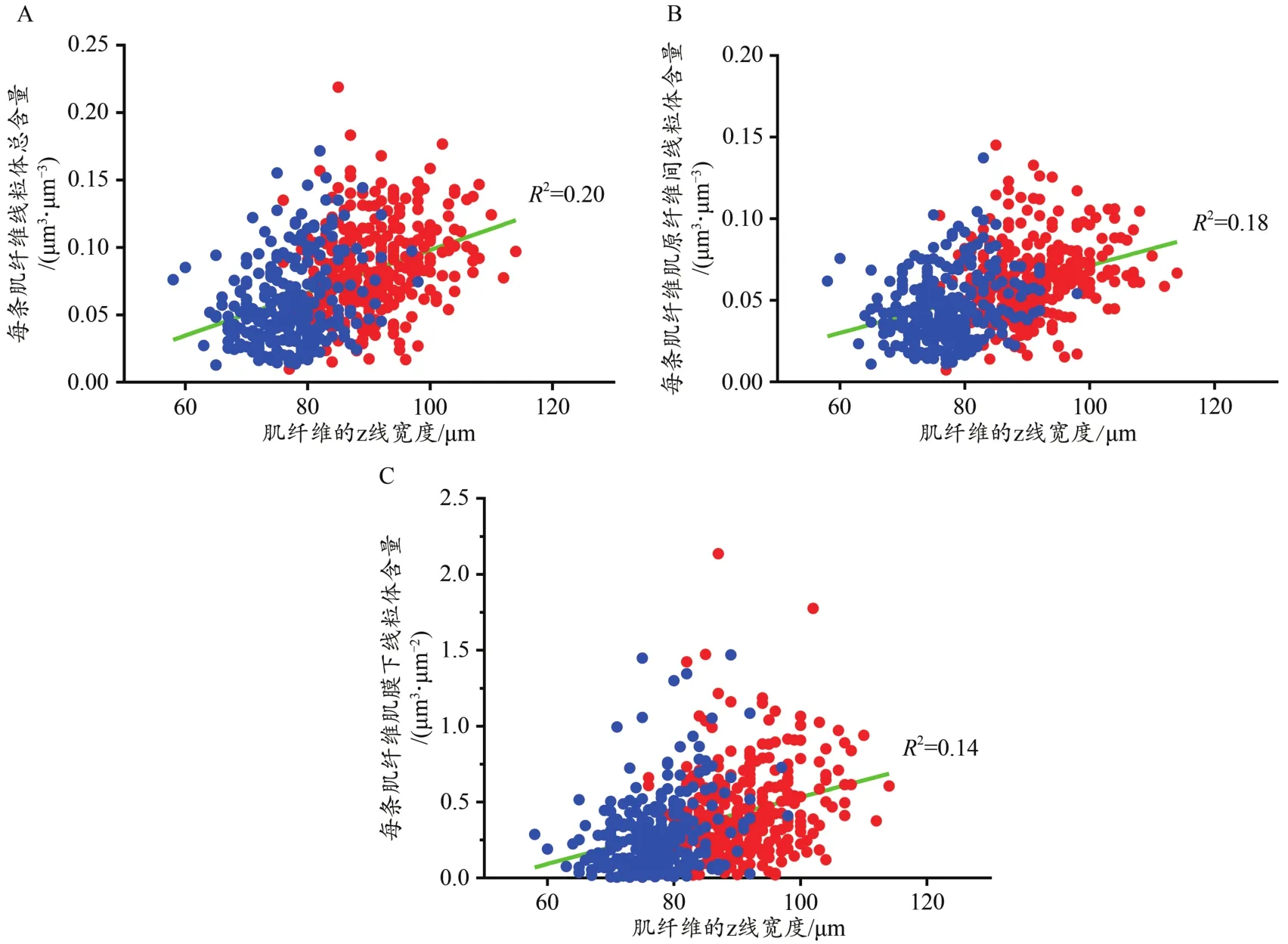
图4 线粒体含量和z线宽度的皮尔逊相关性分析散点图Figure 4.The Pearson’s Correlation Results between the Mitochondrial Contents and z-line Width

表2 不同类型特定亚细胞定位的线粒体含量和其z线宽度的相关性Table 2 The Correlation between the Subcellular Localization Specific Mitochondrial Contents and z-line Width
2.3 不同肌纤维类型和饮食干预下线粒体含量的差异
结果显示,不同类型肌纤维所含线粒体含量有明显差异,Ⅰ型肌纤维的总线粒体IMF和SS线粒体含量均非常显著地高于Ⅱ型肌纤维(P<0.001,图5)。相较于对应的Ⅱ型肌纤维,Ⅰ型肌纤维的平均总线粒体含量高47%;平均IMF线粒体含量,Ⅰ型肌纤维比Ⅱ型肌纤维高41%;平均SS线粒体含量,Ⅰ型肌纤维比Ⅱ型肌纤维高67%。此外,对肌纤维类型和饮食干预这两个组间变量来说,不存在显著性交互作用(P>0.05),故可以直接比较主效应。无论哪种类型的线粒体(总线粒体、IMF线粒体、SS线粒体),Ⅰ型肌纤维的线粒体含量都非常显著地多于Ⅱ型肌纤维(P<0.001)。而且,任何一种短期饮食干预之间都不存在线粒体含量的显著性差异(P>0.05)。由表3可知,在3种不同的饮食干预下,不同肌纤维类型间特定区域化线粒体含量的比较也有明显差异,Ⅰ型肌纤维IMF线粒体含量和体积占比显著高于Ⅱ型肌纤维(P<0.001),Ⅰ型肌纤维SS线粒体含量和体积占比显著高于Ⅱ型肌纤维(P<0.001);各饮食干预组之间的线粒体含量及肌纤维体积占比无明显差异(P>0.05)。

表3 3种短期饮食干预下Ⅰ型、Ⅱ型肌纤维中亚细胞特定区域化线粒体含量均值Table 3 Subcellular Localization Specific Mitochondrial Contents(Mean±SD)in Type Ⅰ and Ⅱ Muscle Fibers under Different Diet Intervention

图5 不同肌纤维类型和3种饮食干预的线粒体含量差异柱状图Figure 5.The Mitochondrial Contents in Three Different Diet Interventions
3 分析与讨论
3.1 特定区域化线粒体含量与肌纤维类型
早期研究证实,Ⅰ型和Ⅱ型肌纤维的结构和代谢特性存在明显差异(Hoppeler et al.,2003)。体育科学界基本达成了“Ⅰ型肌纤维主导有氧代谢”的共识(Ørtenblad et al.,2018;Schiaffino et al.,2011)。据此,线粒体含量也被证实其与肌纤维类型的高度依赖性,即相比于Ⅱ型,Ⅰ型肌纤维中的线粒体总含量更多,但不同肌纤维类型中不同区域分布下的线粒体含量特征并不清楚。已有研究结果表明,性别、身体状况等因素不会影响普通人群的线粒体总量,普通人群的肌纤维线粒体总量均随着耐力运动训练有所提升,且具有区域化特异性:相较IMF线粒体,SS线粒体含量对耐力运动适应的敏感度更高(Devries et al.,2013;Hoppeler et al.,1985)。关于不同肌纤维类型的线粒体含量对比,Howald等(1985)的研究显示,普通人群的总线粒体的体积密度分数Ⅰ型肌纤维约为6%,Ⅱ型肌纤维约为4.5%,但是此实验并没有比较不同肌纤维类型下的区域化线粒体含量特性。本研究中,Ⅰ型肌纤维的总线粒体的体积密度分数为8.7%,Ⅱ型的肌纤维总线粒体的体积密度分数为5.9%,数值均高于上述研究中普通人群的相关指标。由于本研究中受试者为耐力项目的业余爱好者,最大摄氧量水平均超过45 ml/(min·kg-1),他们的有氧能力较普通人群更好,这可能是本研究线粒体含量数值整体水平偏高的原因之一。本研究在此基础上对不同肌纤维类型中特定区域化分布的线粒体含量进行了对比,结果显示,除了Ⅰ和Ⅱ型肌纤维总线粒体含量上的明显差异外,不同亚细胞区域定位下(IMF,SS),Ⅰ型肌纤维的平均线粒体含量全部显著高于Ⅱ型肌纤维的平均线粒体含量。本研究结果表明,Ⅰ型肌纤维的平均总线粒体含量比与Ⅱ型高47%;Ⅰ型肌纤维的平均IMF线粒体含量比Ⅱ型肌纤维高41%;而相比于Ⅱ型肌纤维的平均SS线粒体含量,Ⅰ型肌纤维高67%。这个比例的差异显示SS线粒体含量对Ⅰ型肌纤维的依赖度更高,该研究结果与前人研究一致(Nielsen et al.,2010),再次验证SS线粒体对耐力训练的敏感性更高。
关于肌纤维类型的判定标准,所有人群不同类型肌纤维的z线宽度均呈现两极分化(Prince et al.,1981),且z线宽度不易受任何外界干预或者是长期运动训练的影响,是肌纤维超微结构下最不具备可塑性特征的指标之一,因此z线宽度被认为是直观判断肌纤维类型的潜在指标。Sjöström等(1982)据此思路量化了不同肌纤维类型下的z线宽度,研究结果显示,Ⅰ型肌纤维z线的平均宽度最大,为125±11 nm;平均宽度最小的则是Ⅱ型肌纤维,Ⅱb肌纤维z线宽度均值为86±8 nm。此研究团队通过判别分析的统计学方法证明,仅通过z线宽度判定肌纤维类型的准备率高达83%,因此得出结论,z线宽度是判定骨骼肌纤维类型的较优指标之一。然而,使用z线宽度作为肌纤维类型判定的方法近年来仍然没有被广泛使用,该方法未被广泛使用的一个重要原因在于,没有足够多或者数据量足够多的研究作为数据支撑。近年来的研究指出,不同肌纤维类型中肌球蛋白ATP酶活性会影响z线宽度和肌纤维类型的关系(Yan et al.,2010)。部分研究根据肌球蛋白重链收缩特性差异,通过肌纤维银染分析的方法(Sjöström et al.,1982;Talmadge et al.,1993)进行肌纤维类型判定,但操作复杂程度大大增加。本研究通过大数据量特定区域化线粒体含量和z线宽度的相关性分析,验证了z线宽度作为肌纤维类型判定依据的有效性。结果显示,肌纤维特定区域化线粒体含量和其z线宽度呈显著正相关关系。这也为前人研究(Sjöström et al.,1982)中提到的使用z线宽度作为判定肌纤维类型的直接指标,提供了更充足的证据支持。
3.2 短期不同饮食干预对肌纤维线粒体含量的影响
1967年,Bergstrom等(1967)的实验结果首次证明,短期高碳水化合物饮食干预对比混合饮食或者高脂饮食,能明显提高受试者耐力力竭测试运动表现,力竭时间的差异与测试前的糖原储备有高度相关性。本研究采用相似的实验方案,受试者在正式实验前两周开始适应混合饮食的基础上,先后进行为期3天的高脂肪+高蛋白质饮食以及高碳水化合物的饮食干预方案,并比较不同短期饮食干预下亚极量(标准75%O2max强度的运动模型)运动表现的差异。本研究结果表明,相比于其他两种营养干预方案,3天高碳水化合物饮食干预下的亚极量运动测试力竭时间显著更长,且高脂脂肪+高蛋白质饮食方案下的力竭时间在3种方案中最短,此实验方案直接印证了机体的肌糖原储备对中高强度的运动表现的增益作用。此外有研究发现,耐力运动会引发肌纤维中肌糖原区域化的特异性表达。在耐力运动后48 h内,人体肌纤维中均呈现SS肌糖原含量消耗最少,但此区域的肌糖原后续恢复较慢的特征(Marchand et al.,2007)。后续研究显示,短期饮食干预会造成不同区域下肌糖原的特异性表达,SS肌糖原对于碳水化合物的额外补充敏感性最强。在短期的高碳水化合物补充的条件下,肌糖原总量提升,且SS的肌糖原储量相比分布于肌原纤维区域的肌糖原含量明显更高,并显示出更强的有氧运动能力(Jensen et al.,2020)。与此相关联的发现是区域化线粒体含量也会随之产生变化,作为机体向骨骼肌提供ATP的代谢通路中非常关键的转运蛋白,葡萄糖转运体4(glucose transporter type 4,GLUT-4)在人体进行中高强度耐力运动时显著增加,且在SS区域表达更明显(Lauritzen et al.,2008)。而GLUT-4也会通过同一条代谢通路调节线粒体的特定表达,这种长期的运动训练适应会导致肌纤维中线粒体总量的增加(Holloszy,2011;Chomentowski et al.,2011)。但是上述提到的肌糖原特异性与耐力运动表现的紧密联系是否会引起不同区域分布的线粒体发生特定的变化,则鲜见报道。本研究据此思路,针对不同糖原储备的短期饮食干预下的区域化线粒体含量进行比较。
相比于其他两种营养干预方案,3天高碳水化合物饮食干预下的亚极量运动测试力竭时间显著更长,且高脂肪+高蛋白质方案下的力竭时间最短,此实验方案直接印证了机体的肌糖原储备对中高强度的运动表现的增益作用。本实验室的前期研究(Jensen et al.,2020)使用相同的运动模型证明,糖原的机体储备以及长时间亚极量运动中的糖原消耗均具有亚细胞区域化的特异性:在3天的不同饮食干预下,高糖膳食组的Ⅰ型肌纤维IMF肌糖原储备显著高于高脂低糖组或者混合饮食组,SS肌糖原含量则在3组之间无显著性差异。而高糖膳食组Ⅱ型肌纤维的IMF肌糖原含量仅显著高于高脂低糖组,其他各个组别之间则无显著性差异。然而实验并未进一步探究“糖原池”的结构特异性是否会引起相应的特定区域分布的线粒体含量的适应。本研究结果显示,不同能源底物配比(高碳水化合物饮食或高脂肪+高蛋白质饮食)的短期饮食干预下,骨骼肌细胞特定区域化线粒体含量无明显差异;3次不同的饮食干预下,人体肌纤维中均维持了相同的线粒体含量比例,且在Ⅰ、Ⅱ型肌纤维中均呈现IMF线粒体体积占比更多的情况。结果表明,短期饮食干预对亚极量耐力性力竭实验运动表现的显著影响与线粒体含量的区域化分布无关。大量研究证明,长期的耐力训练适应会增加人体肌纤维的区域化线粒体含量,且SS区域线粒体含量的增加更加明显(Bizeau et al.,1998;Nielsen et al.,2010)。因此,本研究并未发现线粒体含量显著变化的原因可能是区域化线粒体含量的适应性改变与长期的运动训练关系更大,而短期的高碳水化合物饮食干预引发的运动表现差异并非通过影响线粒体含量的区域化分布而实现。另一方面,本研究饮食干预时间较短,且目前鲜有关于长期高碳水化合物饮食干预对特定区域化线粒体含量的直接影响的实验。因此,关于特定区域分布的糖原和线粒体之间的特定通路对中高强度耐力项目的运动训练指导意义,需要进一步的研究探讨和证实。
4 结论
Ⅰ型肌纤维中线粒体总量多于Ⅱ型肌纤维,且在肌细胞微观特性视角下,亚细胞不同区域下分布的线粒体(IMF和SS线粒体),均呈现Ⅰ型肌纤维中的分布含量多于Ⅱ型肌纤维的状态;不同肌纤维的总线粒体含量为z线宽度作为区分快慢肌的判断方法提供了更直接的数据支撑;不同糖原储备的短期饮食干预下长时间亚极量运动至力竭时间有显著性差异,但是该运动表现的差异与亚细胞特定区域化线粒体含量无关。
