灰毡毛忍冬Lm4CL基因克隆及表达分析
2021-06-01乔中全王晓明曾慧杰刘思思彭继庆黄益智李永欣
乔中全,王晓明,曾慧杰,刘思思,蔡 能,彭继庆,黄益智,李永欣
(1.湖南省林业科学院,湖南 长沙 410004;2.湖南省林下特色生物资源培育与利用工程技术研究中心,湖南 长沙 410004;3.中南林业科技大学 生命科学与技术学院,湖南 长沙 410004)
灰毡毛忍冬Lonicera macranthoidesHand-Mazz为忍冬科Caprifoliaceae忍冬属Lonicera植物,是中药材山银花的主要来源植物之一,具有清热解毒、疏散风热功能,用于治疗痈肿疔疮、喉痹、丹毒、热毒血痢、风热感冒、温病发热[1]。根据功能成分、药理药效等方面的研究,灰毡毛忍冬的主要活性成分是绿原酸(Chlorogenic acid,CGA)[2-3],且含量较高,是绿原酸提取的主要原料[4-5],已应用于制药、保健品和化妆品等,市场开发潜力大。灰毡毛忍冬的绿原酸含量与生物合成途径相关基因的表达量关系紧密,随着分子技术的快速发展,药用植物功能基因研究已成为当前的研究热点。
4-香豆酸辅酶A连接酶(4CL)是苯丙烷代谢途径转向下游分支途径的最后一个关键酶,以肉桂酸、4-香豆酸、咖啡酸、芥子酸、阿魏酸和5-羟基阿魏酸等为底物,生成相应的酰基辅酶A酯,然后在羟基肉桂酰辅酶奎宁酸羟基肉桂酰转移酶(HQT)的作用下生成绿原酸[6-9]。1987年,研究者第一次在欧芹Petroselinum hortense中克隆出2个4CL基因[10],之后在多种植物中克隆出4CL基因,其存在于一个含有2~5名成员的基因家族中[11]。目前有研究者已对忍冬Lonicera japonica[12-13]、丹参Salvia miltiorrhiza[14-15]、黄芩Scutellaria baicalensis[16]、三枝九叶草Epimedium sagittatum[17]、圣罗勒Ocimum basilicum[18]、芸香Ruta graveolens[19]、红花Carthamus tinctorius[20]、桑葚Morus notabilis[21]、枇杷Eriobotrya japonica[22]、葛根Pueraria lobata[23]、多穗柯Lithocarpus polystachyus[24]等药用植物进行4CL基因克隆、生物信息学及表达模式分析的研究,但未见灰毡毛忍冬4CL基因研究的有关报道。4CL基因在活性功能调控研究方面更多集中在木质素的生物合成,在非木质素类的中间衍生物方面研究的并不多。鉴于4CL基因在绿原酸合成途径中的重要性,本研究依据灰毡毛忍冬转录组数据库,获得2个Lm4CL基因,并进行基因结构与表达分析,为深入研究灰毡毛忍冬Lm4CL基因在绿原酸合成途径中的分子调控机理奠定理论基础。
1 材料与方法
1.1 材 料
1.1.1 植物材料
供试材料采自湖南省林业科学院忍冬属植物种质资源圃,经湖南省林业科学院王晓明研究员鉴定为灰毡毛忍冬新品种‘龙花’Lonicera macranthoides‘Longhua’(品种权号:20180256)。随机选择3株生长健壮、无病虫害的植株,于2019年3月10日采集幼叶和幼茎,4月27日采集成熟叶和主茎(完全木质化),5—6月采集早期花(呈棒状,长0.5~1.0 cm,上部稍膨大,表面绿色)、中期花(呈棒状,长1.5~2.5 cm,顶部似米粒,表面绿色至黄绿色)、末期花(呈棒状,长3.0~4.5 cm,顶部膨大,向内弯曲,表面黄色至黄白色)(图1),将采集到的样品用密封袋封好,迅速置于液氮中速冻,于-80℃保存备用。

图1 灰毡毛忍冬的茎、叶和花Fig.1 Stems,leaves and flowers of L.macranthoides
1.1.2 试剂与仪器
EASYspin Plus多糖多酚复杂植物RNA快速提取试剂盒,北京艾德莱生物科技有限公司;PrimeSTAR®GXL DNA Polymerase高保真酶、PrimeScriptTMII 1st Strand cDNA Synthesis Kit克隆用反转录试剂盒、TaKaRa Agarose Gel DNA Extraction Kit Ver.4.0胶回收试剂盒、Prime ScriptTMRT reagent Kit定量用反转录试剂盒、SYBR®Premix Ex TaqTM-Tli RNaseH Plus荧光定量试剂盒,宝生物工程(大连)有限公司;PUCm-T克隆载体、pCold表达载体,上海生工生物工程有限公司;绿原酸>98%,中国食品药品检定研究院;Veriti梯度PCR仪,美国ABI;Step one荧光定量PCR仪,美国ABI;LC-20AT高效液相色谱仪,日本岛津。
1.2 方 法
1.2.1 灰毡毛忍冬总RNA的提取与检测
按照制造商的说明,使用EASYspin Plus多糖多酚复杂植物RNA快速提取试剂盒(艾德莱,中国)从‘龙花’茎、叶和花中分别提取总RNA。1%琼脂糖凝胶电泳检测RNA完整性,使用Eppendorf生物光度计(德国)测定RNA浓度和纯度。
1.2.2 灰毡毛忍冬4CL基因中间片段的克隆
基于灰毡毛忍冬的转录组数据,通过KEGG分析,4CL的注释unigenes共有21个,长度从202 bp到2 344 bp,与拟南芥、粳稻、葡萄的序列相似。剔除未Nr注释及Nr注释得分低于100分的unigenes,留下与GenBank中已知4CL基因CDS区域长度比较一致的unigenes。比较发现,Unigene18557_All、Unigene18499_All与 已 知 的4CL基因高度同源,并作为核心片段进行后续分析,根据其序列设计引物(表1),并由苏州金唯智生物科技有限公司合成。以cDNA为模板进行PCR扩增,反应体系为:正、反引物各0.6 μL,cDNA模板2 μL,反应液12.5 μL,ddH2O 9.3 μL,总体积25 μL。反应程序:94℃ 5 min,94℃ 45 s,48℃ 45 s,72℃ 2 min,35个循环,72℃ 10 min。用1 % 琼脂糖凝胶电泳进行PCR产物观察,回收产物连接PUCm-T载体,经蓝白斑筛选、测序。
1.2.3 灰毡毛忍冬4CL基因5′和3′末端RACE的扩增
根据4CL基因中间片段,设计末端扩增特异性引物(表1),按照SMARTerTMRACE试剂盒扩增步骤进行。扩增结果用1%琼脂糖凝胶电泳分检测,PCR产物连接pUCm-T载体,经蓝白斑筛选、测序。
1.2.4 灰毡毛忍冬4CL基因全长CDS的克隆
使用DNAMAN软件对5′RACE和3′ RACE测序结果进行序列拼接,获得4CL基因的全长cDNA,根据全长序列设计CDS扩增引物(表1),并由苏州金唯智生物科技有限公司合成,以cDNA为模板,PCR反应程序为:94℃ 2 min,94℃ 30 s,56℃ 30 s,72℃ 2 min,35个循环,72℃延伸5 min。
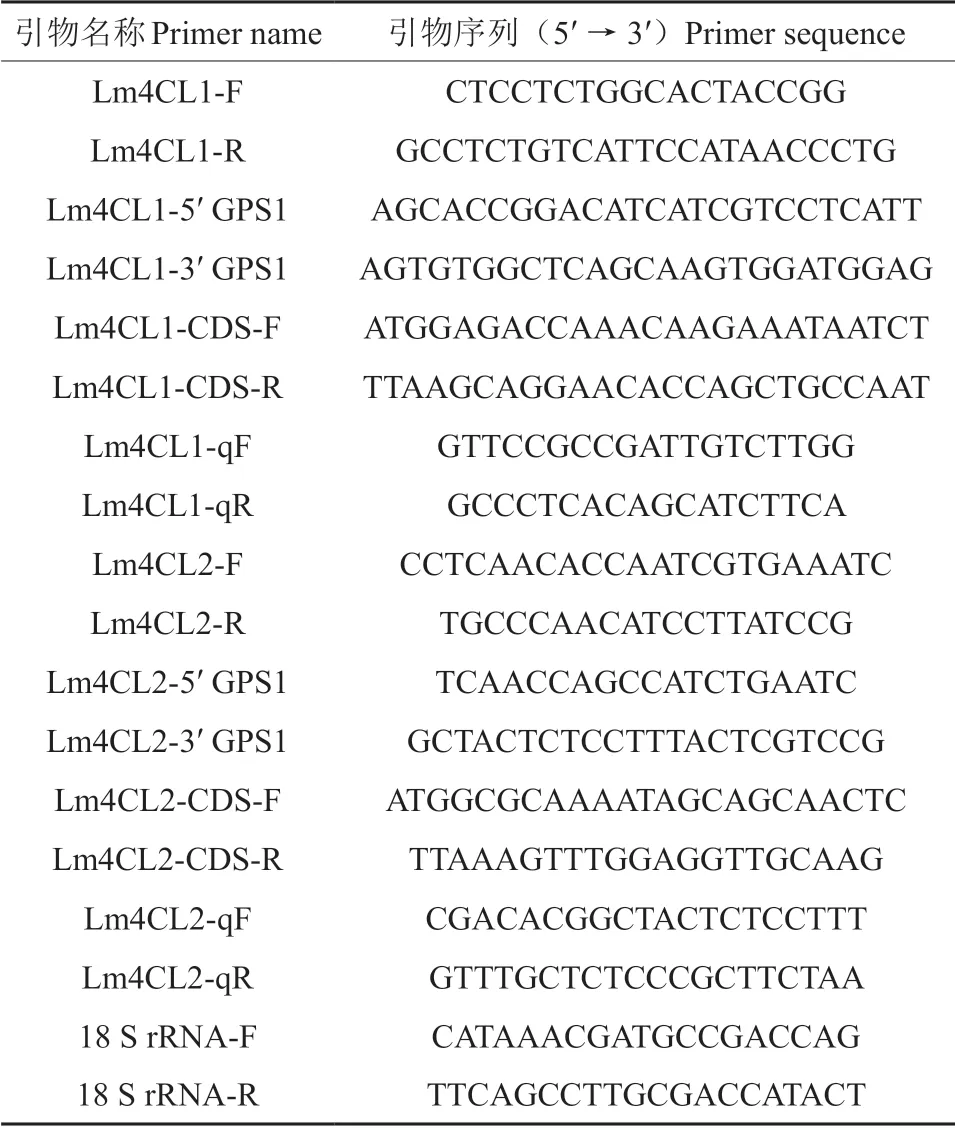
表1 引物名称及序列Table 1 Primer name and sequence
1.2.5 灰毡毛忍冬4CL基因的生物信息学分析
使用NCBI(https://blast.ncbi.nlm.nih.gov)进行基因功能注释,Pfam(https://pfam.xfam.org/search)进行蛋白结构域预测,MEME( http://meme.nbcr.net/meme/)进行保守基序(Motif)搜索,ClustalW( http://www.genome.jp/tools-bin/clustalw)进行多序列比对,Swiss model(https://swissmodel.expasy.org/interactive)进行蛋白质三级结构预测,ProtParam( http://web.expasy.org/protparam/)进行蛋白特性分析,TMHMM2.0( http://www.cbs.dtu.dk/servi-ces/TMHMM/)进行蛋白质跨膜结构预测,SignalP( http://www.cbs.dtu.dk/services/SignalP/)进行信号肽分析,PSORT Prediction( http://psort1.hgc.jp/form.html)进行亚细胞定位预测,MEGA6.0软件构建系统进化树[25]。
1.2.6 灰毡毛忍冬4CL基因原核表达载体的构建
用Xhol I和Hind III两种酶对回收后的PCR产物、pCold载体进行双酶切,将纯化后的pCold和PCR产物用T4 DNA连接酶4℃连接过夜。通过热激转化的方法将连接产物转化到DH5α感受态中,在转化后的产物中加入LB液体培养基,37℃、170 r/min震荡培养45 min,6 000 r/min离心1 min后,倒掉部分上清液,取150 μL涂在含100 μg/mL氨苄固体培养基上,37℃培养过夜。次日挑选阳性克隆菌斑PCR鉴定后,送至长沙擎科公司测序。
1.2.7 工程菌的制备
提取质粒并转化到宿主菌BL21 star (DE3)中,涂布于含有100 μg/mL氨苄青霉素的LB固体培养基上,37℃倒置培养、过夜,随机挑选单菌落,进行菌液PCR鉴定。
1.2.8 基因的诱导表达
将阳性工程菌在含有100 μg/mL的氨苄青霉素LB液体培养基中培养至吸光度达到0.6左右时,加入终浓度分别为0.0、0.4、0.6、0.8 mmol/L的IPTG诱导表达,SDS-PAGE电泳鉴定表达情况。
1.2.9 灰毡毛忍冬4CL基因的表达模式分析及绿原酸含量的检测
荧光引物(表1)使用在线软件Primer-BLAST(https://www.ncbi.nlm.nih.gov/tools/primer-blast/index.cgi?LINK_LOC=BlastHome)进行设计。以18 S为内参基因,采用SYBR Green荧光染料法进行基因的相对表达水平分析。正、反引物各0.3 μL,cDNA 1.5 μL,SYBR qPCR Master Mix 10 μL,ddH2O 7.9 μL,反应总体积为20 μL。反应程序参照荧光定量试剂盒说明书。
采用高效液相法(HPLC)测定绿原酸含量,参照《中国药典》2015年版制备样品溶液,设定液相条件[1]。
2 结果与分析
2.1 灰毡毛忍冬总RNA的提取与检测
以灰毡毛忍冬叶片为材料提取的总RNA,核酸蛋白检测仪(Eppendorf)测得A260/A280=1.98、A260/A280=2.22,说明RNA纯度高,没有蛋白、盐离子等污染;琼脂糖凝胶电泳检测(图2A)显示,28 S、18 S rRNA条带清晰,且28 S rRNA条带亮度大约为18 S rRNA条带亮度的2倍,表明提取的总RNA完整、无降解。
2.2 灰毡毛忍冬Lm4CL基因全长cDNA的获得
以灰毡毛忍冬转录组数据为基础,筛选出2个编码4-香豆酸:CoA连接酶的Unigene18557_All、Unigene18499_All部分序列。利用RACE技术,从灰毡毛忍冬叶片中成功扩增出两个全长cDNAs,分别命名为Lm4CL1、Lm4CL2,序列提交GenBank(登录号:MN103349、MT123898)(图2D)。Lm4CL1基因全长为1 960 bp,5′端非编码区(UTR)长68 bp,3′端非编码区(UTR)长275 bp,ORF(Open Reading Frame)长1 617 bp,编码538个氨基酸;Lm4CL2基因全长2 133 bp,5′端非编码区(UTR)长176 bp,3′端非编码区(UTR)长273 bp,ORF长1 683 bp,编码560个氨基酸。
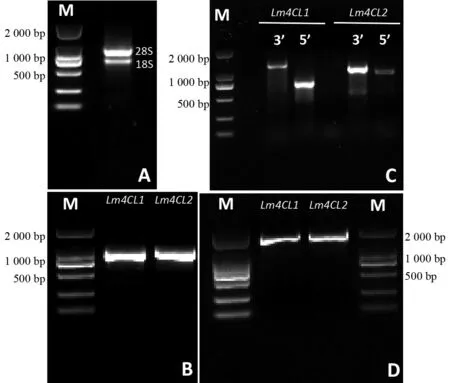
图2 灰毡毛忍冬Lm4CLs基因的克隆Fig.2 Cloing of Lm4CLs in L.macranthoides
2.3 灰毡毛忍冬Lm4CL基因生物信息学分析
2.3.1 灰毡毛忍冬Lm4CL基因编码蛋白质理化性质分析
ProtParam分析结果(表2)表明,Lm4CL1蛋白的分子质量约为58 736.29,等电点(pI)为5.73,总原子个数为8 317,分子式为C2635H4200N680O771S31,带正电残基(Arg+Lys)个数为55,带负电残基(Asp+Glu)个数为63,脂肪系数为94.24,不稳定系数为40.84>40,推测该蛋白属于不稳定蛋白,亲水性平均系数(GRAVY)为0.056;Lm4CL2蛋白的分子质量约为60 387.24,等电点(pI)为8.15,总原子个数为8 550,分子式为C2690H4306N718O817S19,带正电残基(Arg+Lys)个数为54,带负电残基(Asp+Glu)个数为52,脂肪系数为92.3,不稳定系数为30.95<40,推测该蛋白属于稳定蛋白,亲水性平均系数(GRAVY)为-0.003。且ProtScale分析表明,Lm4CL1疏水性氨基酸分布大于亲水性氨基酸,Lm4CL2疏水性氨基酸分布小于亲水性氨基酸,推测Lm4CL1为疏水性蛋白,Lm4CL2为亲水性蛋白。跨膜结构预测显示,Lm4CL1没有跨膜结构,Lm4CL2在氨基酸位点的前100和200~300有两个跨膜区域;信号肽位点预测显示,Lm4CL1和Lm4CL2均不存在信号肽位点;亚细胞定位显示,Lm4CL1定位于内质网,Lm4CL2定位于叶绿体类囊体。
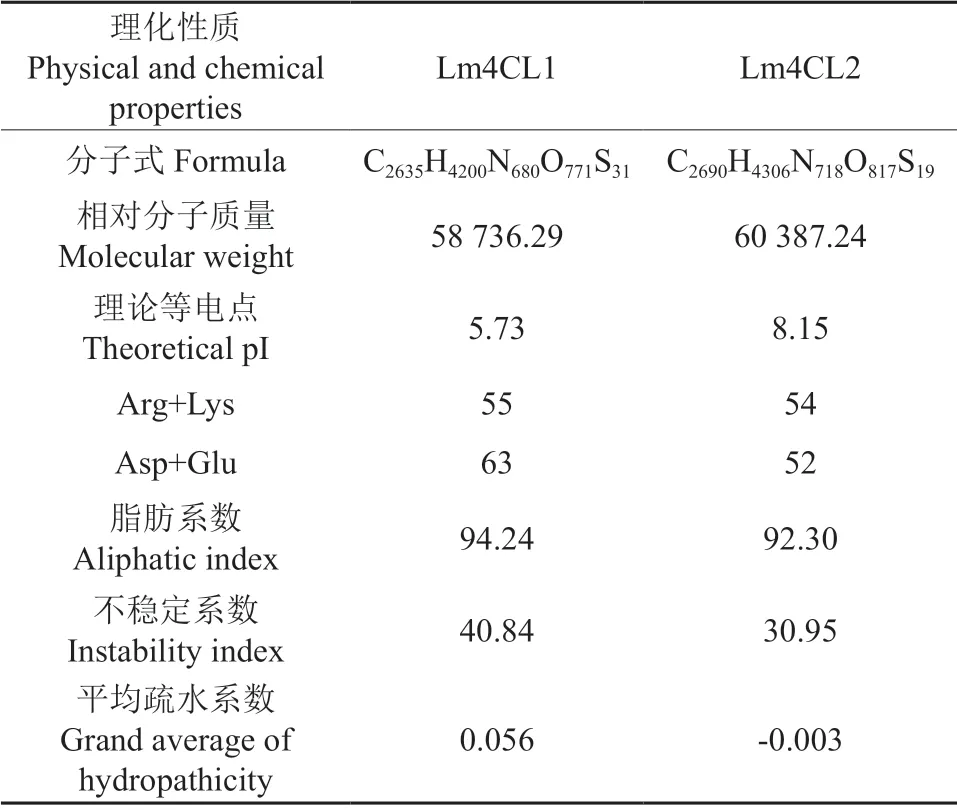
表2 灰毡毛忍冬Lm4CL1蛋白的理化性质Table 2 Physical and chemical properties of Lm4CL1 in L.macranthoides
2.3.2 灰毡毛忍冬Lm4CLs蛋白同源性及系统进化分析
以拟南芥(At4CL1_AAA82888,At4CL2_AAD47193,At4CL3_AAD47195)、毛白杨(Pto4CL1_AAL02145,Pto4CL2_AFC89538)、大豆(Gm4CL1_AAL98709,Gm4CL2_AAC97600)、烟草(Nt4CL2_AAB18638.1)等蛋白的氨基酸序列为参考,使用DNAMAN软件进行序列比对,总相似性达到69.62%,灰毡毛忍冬Lm4CL1与Lm4CL2蛋白的相似性为35.15%,Lm4CL1、Lm4CL2蛋白与烟草Nt4CL2蛋白的相似性分别为83.58%、39.27%。Lm4CL1、Lm4CL2蛋白都含有3个保守的催化残基,分别是催化硫酯合成反应的Lys-441和Gln-446及催化半腺苷化反应的Lys-526(图3,保守催化残基用黑色三角形表示,相对于Nt4CL2蛋白中氨基酸的位置)。与底物结合的区域位于SSGTTGLPKGV(Box I)和GEICIRG(Box II)这两个AMP结合位点中间,在Lm4CL2蛋白中两个位点分别被SSGTTGASKGV和GEIWLRG取代。

图3 灰毡毛忍冬与其它物种的4CL蛋白序列比对Fig.3 Alignment of Lm4CLs with others
为了研究灰毡毛忍冬Lm4CLs家族成员的亲缘关系,基于Lm4CL1、Lm4CL2蛋白的氨基酸序列,同时选择拟南芥Arabidopsis thaliana、烟草Nicotiana tabacum、杨树Populus tremuloides、杂交杨Populus trichocarpa×Populus deltoides、毛白杨Populus tomentosa、大 豆Glycine max、忍冬Lonicera japonica、桑葚Morus notabilis、丹参Salvia miltiorrhiza、三枝九叶草Epimedium sagittatum、覆盆子Rubus idaeus、芸香Ruta graveolens、紫草Lithospermum erythrorhizon、茶树Camellia sinensis、藿香Agastache rugosa、黄芩Scutellaria baicalensis、红花Carthamus tinctorius、桔梗Platycodon grandiflorus、枇杷Eriobotrya japonica、圣罗勒Ocimum basilicum、葛根Pueraria lobata等21种植物的4CL蛋白序列作为参照,使用MEGA X软件、邻近法(Neighborjoining)构建系统进化树。聚类结果(图4)显示,Lm4CL1属于ClassⅠ蛋白,Lm4CL2属于ClassⅡ蛋白;Lm4CL1与忍冬(Lj4CL1)亲缘关系最近,Lm4CL2与丹参(Sm4CL4)亲缘关系最近。

图4 不同植物中4CLs蛋白序列构建的系统树Fig.4 Phylogenetic tree of 4CLs in different plants
2.3.3 灰毡毛忍冬Lm4CLs蛋白结构分析
以Lm4CL1、Lm4CL2蛋白的氨基酸序列为基础,构建了这两种4-香豆酸辅酶A连接酶的三维(3D)蛋白模型,阐明了蛋白的结构特征。蛋白质二级结构分析表明,Lm4CL1有17个α-helices和21个β-strands(图5A),Lm4CL2有17α-helices和25个β-strands(图5B)。
使用烟草4CL2-5bsr.1.A (Nt4CL2)作为同源建模的模板,生成Lm4CL1蛋白的三级结构(图5C),Lm4CL1蛋白与模板的序列相似性为83.58%,模型的GMQE为0.96,QMEAN为0.65,建模质量良好。Lm4CL1蛋白的三级结构由一个大的N域和一个小的C域组成,N域由425个残基组成(Glu-6到Asp-430),C域由96个残基组成(Lys-437到Leu-532),N域和C域之间通过一个高度灵活的链接器“铰链环”(Arg-431到Ile-436)连接。
使用烟草4CL2-5bsr.1.A (Nt4CL2)作为同源建模的模板,生成Lm4CL2的三维模型(图5D),Lm4CL2与模板的序列相似性为39.27%,模型的GMQE为0.70,QMEAN为-2.27,建模质量良好。Lm4CL2的模型结构由一个大的N域和一个小的C域组成,N域由529个残基组成(Asn-26到Asp-454),C域由96个残基组成(Lys-461到Ala-556)。N域和C域之间通过一个高度灵活的链接器“铰链环”(Arg-455到Ile-460)连接。
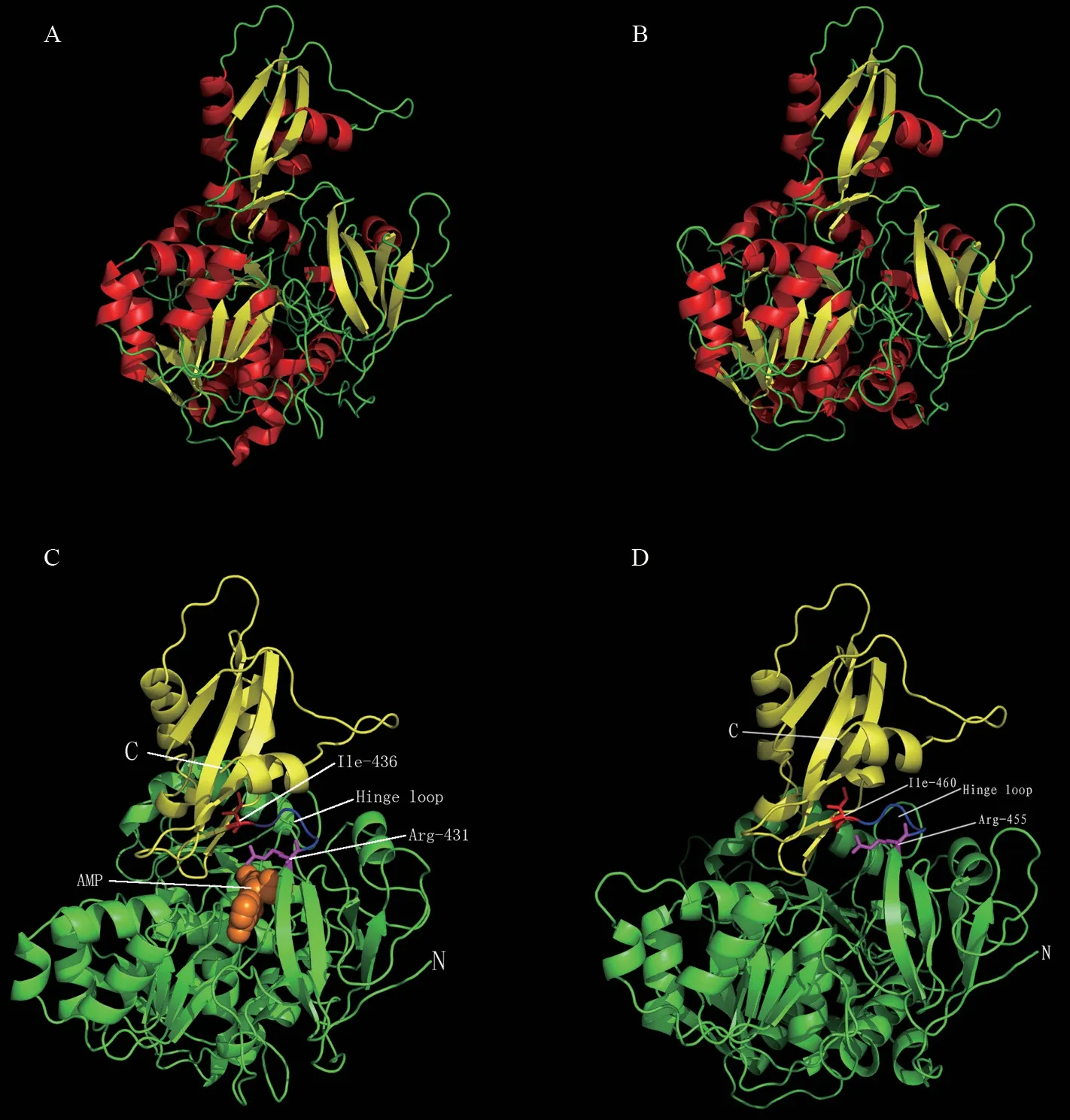
图5 Lm4CL1和Lm4CL2的蛋白结构模型Fig.5 Protein structural models of Lm4CL1 (A,C) and Lm 4CL2 (B,D)
2.4 灰毡毛忍冬Lm4CL1基因的原核表达
将构建的重组质粒pCold-Lm4CL转化大肠杆菌BL 21 star (DE3)菌株的感受态细胞后,用IPTG诱导表达,经SDS-PAGE电泳分析(图6),空白对照和阴性对照都没目的蛋白表达,而转入重组质粒pCold-Lm4CL的菌体经浓度为0.4、0.6、0.8 mmol/L ITPG15℃诱导24 h后,在100 kD附近有清晰的蛋白条带,该蛋白大小与目的蛋白大小一致(Lm4CL1蛋白大小为58.74 kD,pCold在N端有一个45.0 kD的分子伴侣TF)。此外,蛋白的表达量随着IPTG浓度的提高表现出先增后降的规律,IPTG浓度为0.6 mmol/L时蛋白表达量最高。

图 6 Lm4CL1基因表达产物的SDS-PAGE分析Fig.6 SDS-PAGE analysis of gene expression products
2.5 灰毡毛忍冬Lm4CLs基因的表达特征及与绿原酸含量的关系
以‘龙花’为试验材料,分别在不同的组织部位及花器官的发育过程中分析了Lm4CL1、Lm4CL2基因的相对表达水平(图7)。qRT-PCR结果显示,Lm4CLs基因的表达存在明显的组织特异性。Lm4CL1基因在茎中表达量最高,其次为叶,花中表达量最低;Lm4CL2基因在花中表达水平最高,其次为叶,茎中表达水平最低。随着‘龙花’花器官的发育,Lm4CL1基因的表达量呈现逐渐降低的趋势,Lm4CL2基因的表达量在前期升高,中后期表达量稳定。
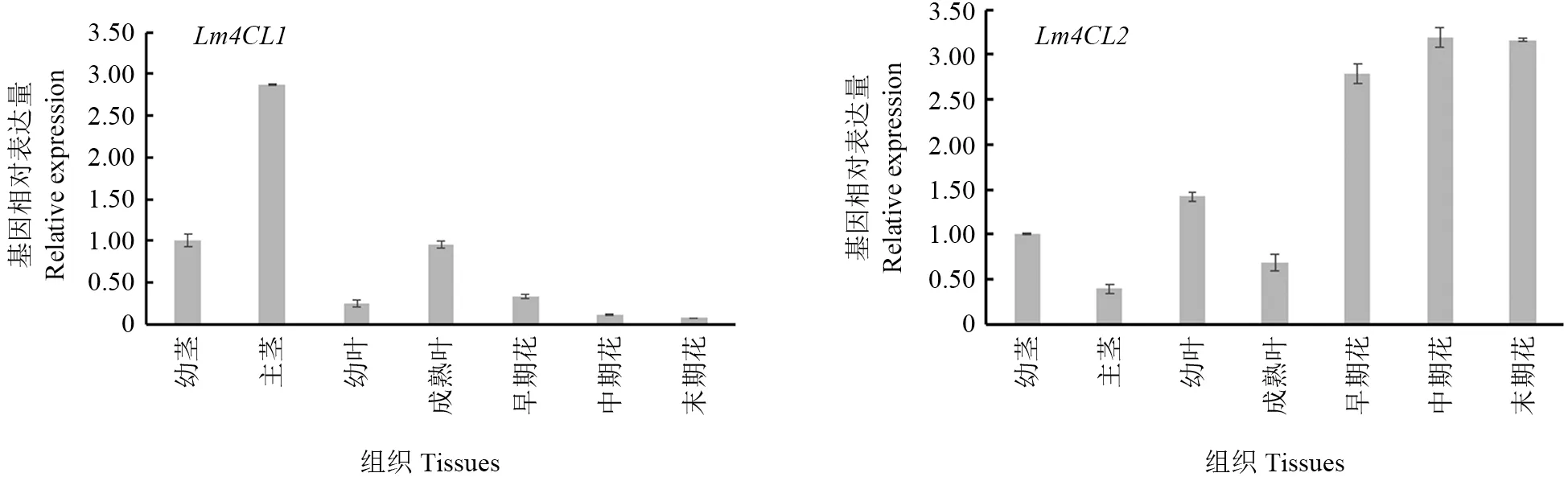
图7 Lm4CLs基因在‘龙花’不同组织部位的表达量Fig.7 Relative expression of Lm4CLs in Lonicera macranthoides ‘Longhua’
绿原酸含量分析结果(图8)显示,‘龙花’的茎、叶和花中均含有绿原酸,花中绿原酸含量显著高于茎和叶。分析Lm4CLs基因表达水平与灰毡毛忍冬不同组织的绿原酸含量的相关性,结果表明Lm4CL1基因的表达量与绿原酸含量呈负相关(r=-0.63),Lm4CL2基因的表达量与绿原酸含量呈正相关(r=0.94)。
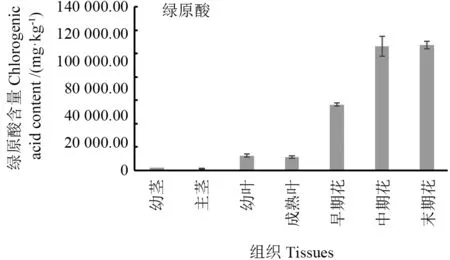
图8 ‘龙花’不同组织部位的绿原酸含量Fig.8 Chlorogenic acid contents in Lonicera macranthoides ‘Longhua’
3 结论与讨论
3.1 讨 论
4CL基因在多种植物中已被成功克隆,并以基因家族的形式存在,如拟南芥中有3个4CL基因(At4CL1,At4CL2,At4CL3)[26],烟草中有3个4CL基因(Nt4CL,Nt4CL1,Nt4CL2)[27],杨树中有2个4CL基因(Pt4CL1,Pt4CL2)[28],杂交杨中有4个4CL基因(Ptd4CL1,Ptd4CL2,Ptd4CL3,Ptd4CL4)[29],大豆中有4个4CL基因(Gm4CL3,Gm4CL4,Gm4CL3,Gm4CL4)[30]。研究发现4CL蛋白分为两个大的进化类群,Ⅰ类主要与木质素的合成有关,如拟南芥(At4CL1,At4CL2)、杨树(Pt4CL1)、杂交杨(Ptd4CL1,Ptd4CL2)等;Ⅱ类与类黄酮的生物合成有关,如拟南芥(At4CL3)、大豆(Gm4CL3、Gm4CL4)、杨树(Pt4CL1)等[11]。本研究通过灰毡毛忍冬转录组分析,筛选出两条注释为4CL的基因片段,并克隆出Lm4CL1、Lm4CL2两个4CL基因的全长cDNA序列。系统进化分析结果显示,Lm4CL1基因与拟南芥中At4CL1、At4CL2基因,大豆中Gm4CL1、Gm4CL2基因,毛白杨Pto4CL1基因聚为Ⅰ类,这些基因都与植物木质素的合成有关;Lm4CL2基因与拟南芥中At4CL3基因、毛白杨中Pto4CL2基因聚为Ⅱ类,而这些基因参与植物的类黄酮及其它物质合成[31-33]。DNAMAN比对这些基因发现,保守结构域Box I(SSGTTGLPKGV)中的亮氨酸(Leu)和脯氨酸(Pro)在Lm4CL2中被丙氨酸(Ala)和丝氨酸(Ser)替代,保守结构域Box II(GEICIRG)中的半胱氨酸(Cys)和异亮氨酸(Ile)被色氨酸(Trp)和亮氨酸(Leu)替代,保守结构域中氨基酸的改变可能直接导致Lm4CL2蛋白具有新的催化功能,但是蛋白结构与功能的关系还需要进一步的研究。
木质部、叶片的维管组织等是植物木质素生物合成的主要部位,在分析灰毡毛忍冬不同组织部位的样品时发现,Lm4CL1基因在幼叶中有表达,在幼茎和主茎中的表达量明显高于其它部位,结合进化分析结果,推测该基因与灰毡毛忍冬木质素的生物合成相关,这与在忍冬中的研究结果不同[13]。Lm4CL2基因在花中表达水平最高,在花器官发育前期表达量急剧升高,后期表达量稳定,且与绿原酸合成呈显著正相关(r=0.94),推测Lm4CL2基因是调控灰毡毛忍冬绿原酸生物合成的重要基因。目前正在构建灰毡毛忍冬遗传转化体系,为明确Lm4CL基因在灰毡毛忍冬中的功能和作用机制提供理论基础。
3.2 结 论
从灰毡毛忍冬新品种‘龙花’中克隆获得Lm4CL1和Lm4CL2两个基因,两者氨基酸相似性为35.15%,系统发育分析显示Lm4CL1属于Ⅰ类蛋白,Lm4CL2属于Ⅱ类蛋白;Lm4CL1在茎中呈现高表达且与绿原酸含量呈负相关(r=-0.63),Lm4CL2在花中呈现高表达且与绿原酸含量呈正相关(r=0.94),推测Lm4CL1基因与木质素合成有关,Lm4CL2基因与绿原酸合成有关。
