樟树MVA途径基因克隆与表达模式初探
2021-06-01
(江西省林业科学院 国家林业和草原局樟树工程技术研究中心,江西 南昌 330032)
樟树Cinnamomum camphora是我国重要的木本精油植物资源之一。枝叶精油是目前樟树主要的利用部位,具有得率高、主成分突出等优点。萜类是樟树枝叶精油的主要成分,常根据各主成分含量差异将樟树划分为不同化学类型。其中,橙花叔醇型樟树精油主成分为橙花叔醇,归类于倍半萜类化学型[1-2]。我国樟属植物中其它倍半萜类化学型还包括金合欢醇型、γ-榄香烯型、布莱醇型、β-荜澄茄油烯型与β-石竹烯型等[3-4]。不同化学型樟属精油用途各异,橙花叔醇型精油主要为高级香水的调香剂和定香剂[5]。
植物化学型是指群体中个体间不能通过形态学区分的化学特性,主要用来描述个体间体内化合物特别是次生代谢物特征。倍半萜类化合物为植物主要次生代谢物之一,目前已在自然界中鉴定获得数万种之多。研究显示,各类倍半萜类化合物皆享有相同的前体物合成路径即甲羟戊酸途径(Mevalonate pathway,MVA)。该途径以2分子乙酰CoA作为启始反应物,通过7步连续酶促反应,最终生成异戊烯基焦磷酸(IPP)与其同分异构体二甲烯丙酯焦磷酸(DMAPP)[6-7]。启始反应步骤为2分子乙酰CoA缩合形成乙酰乙酰-CoA,由乙酰乙酰-CoA硫解酶(AACT)催化完成,该酶通常定位于细胞质或过氧化物酶体中[7]。3-羟基-3-甲基戊二酰CoA合成酶(HMGS)催化第二步反应形成3-羟基-3-甲基戊二酰CoA,随后经HMG-CoA 还原酶(HMGR)还原形成甲羟戊酸[7]。甲羟戊酸为相关合成中枢支点,可进入多种物质合成代谢通路。随后经连续两步磷酸化反应生成甲羟戊酸-5-焦磷酸(PP-MVA),所涉及的两个催化激酶分别为甲羟戊酸激酶(MK)和磷酸甲羟戊酸激酶(PMK)[8]。甲羟戊酸焦磷酸脱羧酶(MVD)进一步将PP-MVA脱羧形成类异戊二烯共同前体物IPP[6]。最终IPP在异构酶(IDI)的作用下,异构形成另一共同前体物DMAPP,IDI同时可调控二者含量相对平衡[9]。
植物MVA途径自1960年代被发现以来,已在多种植物中开展了深入研究。现已发现MVA途径存在关键限速酶,且通过转录水平或转录后水平等对多个编码催化酶基因进行多层次调控。近几年樟树MVA途径研究虽有所涉及,但整条途径基因克隆及其表达调控模式研究还没见报道。橙花叔醇型樟树作为樟属倍半萜类中最常见的化学类型,以其为研究材料,开展倍半萜生物合成相关研究,可为后续樟属良种选育和化学型形成机制提供理论基础。
1 材料与方法
1.1 材料与主要试剂
橙花叔醇型樟树、桉叶油素型樟树与黄樟油素型岩桂C.pauciflorum采自江西省林业科学院樟属植物基因库。樟树MVA途径基因序列参考国家林业和草原局樟树工程技术中心完成的樟树5种化学类型(芳樟醇型、桉叶油素型、樟脑型、龙脑型与橙花椒醇型)叶组织转录组数据[10]、基因组数据和黄樟C.porrectum叶组织全长转录组数据[11]。
Total RNA提取试剂盒购自北京华越洋;2×Phanta Master Mi(P511-01)、反转录试剂盒(HiScript II 1st Strand cDNA Synthesis Kit,R211-01)、荧光定量PCR试剂盒(ChamQ SYBR qPCR Master Mix,Q311-02)、大肠杆菌感受态细胞TOP10与琼脂糖凝胶回收试剂盒等均购自南京诺唯赞;PMD18-T克隆试剂盒购自TaKaRa公司。有机溶剂环己烷分析纯与癸酸乙酯购自阿拉丁。实验室常规试剂和培养基购自上海生工;克隆测序由上海生工完成。
1.2 方 法
1.2.1 MVA途径系列基因克隆与生物信息学分析
以模式植物拟南芥MVA系列基因序列作为查询序列,运行本地blast X软件,比对樟树叶组织转录组和近源种黄樟叶组织全长转录组;根据序列相似性筛选樟树MVA途径系列基因Contigs,进一步借助序列拼接软件CAP3完成樟树MVA途径系列基因全长编码区电子克隆。
采集橙花叔醇型樟树花、叶、根与小枝等组织速冻于液氮中,提取各组织total RNA。DNAase I消化后去除残留DNA,混合均匀后取50 ng total RNA反转录合成cDNA第一链。基于樟树叶组织转录组数据、设计特异引物(表1),以上述混合cDNA为模板,采用PCR法特异扩增樟树MVA系列基因。电泳检测PCR产物,切取与各目标产物大小相当的扩增条带,回收后与克隆载体连接、转化大肠杆菌感受态细胞。筛选阳性克隆,送至上海生工测序,获得各系列基因编码区序列。比对樟树基因组数据,预测各基因内含子与外显子组成结构。

表1 所有引物信息Table 1 List of all primers
利用多种在线软件TargetP 1.1 ( http://www.cbs.dtu.dk/services/TargetP/)、ChloroP 1.1 ( http://www.cbs.dtu.dk/services/ChloroP/)、MitoProt ( http://ihg.gsf.de/ihg/mitoprot.html)和PTS1 ( http://www.peroxisomedb.org/diy_PTS1.html)等预测各系列基因编码蛋白亚细胞的定位情况。
1.2.2 橙花叔醇型樟树各组织精油提取与成分分析
2018年4月,采集5株橙花叔醇型樟树花各1 g,7月采集5株橙花叔醇型樟树其它组织各1 g。将新鲜组织于液氮中研磨,后转移至10 mL玻璃试管中,利用有机溶剂萃取法提取精油。有机溶剂为5 mL环己烷分析纯,添加100 μg癸酸乙酯作为内标。研磨后组织粉末于环己烷溶液中浸泡12 h,之后4 000 r/min离心5 min,取上清液,重复2次。过滤除杂,取500 μL液体转移至进样瓶之中,利用shimadzu气相色谱-质谱联用仪(GC-MS)进行精油成分分析。统计整理各成分的相对含量,根据内标癸酸乙酯含量和进样体积折算各成分的绝对含量。
载气为He,载气流速1.0 mL/min,采用分流进样,分流比20∶1。气相条件(GC):进样口温度280℃。升温程序:初始柱温50℃保持2 min,3℃/min升温至180℃保持2 min,再以8℃/min升温至240℃保持5 min,共运行60 min。质谱条件(MS):接口温度260℃,离子源温度180℃,扫描范围(m/z)50~620。
1.2.3 MVA途径系列基因组织表达特异性分析
2019年4月,分别采集橙花叔醇型樟树叶、叶柄、花、花梗、小枝、树皮与根等7种鲜嫩组织,提取total RNA。以Actin基因作为内参,采用荧光定量PCR方法(qRT-PCR)检测橙花叔醇型樟树各组织中MVA系列基因相对表达量。qRT-PCR引物见表1,反应在Bio-RAD C1000 TM Thermal Cycler 荧光定量PCR仪上完成。反应体系总体积20 μL,包括cDNA 50 ng、2×SYBR Green qPCR 10 μL、10 mmol/L的正反向引物各0.3 μL,ddH2O补足至20 μL。反应程序为95℃ 2 min;95℃ 15 s,58℃ 30 s,72℃ 20 s;40个循环,添加溶解曲线;每个样品重复3次。参照2-△△CT方法计算各基因的相对表达量[12]。
1.2.4 不同化学型樟属植物叶组织中MVA途径系列基因表达比较分析
本实验室尚未收集到叶精油主成分为苯丙素类化合物的樟树化学型。樟属近源种岩桂叶组织富含黄樟油素,故作为苯丙素类化学型代表引入本次分析。桉叶油素型樟树叶精油富含单萜类化合物,在本节分析中作为单萜类化学型代表。
课题组于2018年7月分别采集橙花叔醇型樟树、桉叶油素型樟树与黄樟油素型岩桂各3株新鲜叶组织1 g,提取精油并进行成分分析,方法同1.2.2。于2019年4月采集上述单株鲜嫩叶组织,提取total RNA。以Actin基因作为内参,采用qRTPCR检测MVA系列基因在樟属植物不同化学型叶组织的表达情况。具体引物见表1,方法同1.2.3。
1.2.5 橙花叔醇型樟树叶精油倍半萜含量年变化统计与分析
樟树每年2—3月开始萌发新叶,来年2—3月落叶。2018—2019年,隔月采集小群体橙花叔醇型樟树一个生长周期内(即3、5、7、9、11月至来年1月份)鲜叶各1 g。利用有机溶剂萃取精油,通过GC-MS分析各成分含量,方法同1.2.2。统计各单株叶精油倍半萜含量年平均数并以此为基数,计算每月倍半萜含量并与之比值作图,分析小群体各单株鲜叶精油倍半萜含量的年变化情况。
1.2.6 不用时令下橙花叔醇型樟树叶组织MVA途径系列基因表达变化分析
根据1.2.5分析结果,从小群体中挑选6株代表性单株作为本节实验材料。于2019年3月与7月分别采集新叶组织,提取total RNA,检测MVA途径系列基因在各单株叶组织中的表达量,qRTPCR方法和引物同1.2.3,并与倍半萜含量相关联,分析其对倍半萜合成的调控作用。
2 结果与分析
2.1 MVA途径系列基因克隆与生物信息学分析
以拟南芥MVA途径系列基因蛋白序列作为查询种子,在樟树叶组织转录组中获得11个MVA途径系列同源基因。设计特异克隆引物,以橙花叔醇型樟树叶组织cDNA作为模板,PCR扩增樟树MVA途径系列基因(图1a)。PCR产物切胶回收后连接至克隆载体PMD18-T,挑选阳性单克隆、测序。当中AACT、HMGS、HMGR与IDI等4个酶由2个旁系同源基因编码,其它为单拷贝基因。比对樟树基因组数据,显示该系列基因由4~13个不等外显子组成,AACT1与AACT2、HMGS1与HMGS2、HMGR1与HMGR2及IDI1与IDI2等旁系同源基因皆包含相同外显子数目,即内含子-外显子结构相同(图1b)。

图1 樟树MVA途径系列基因克隆与内含子-外显子结构预测Fig.1 PCR cloning and exon-intron structures of the MVA pathway in C.camphora
CcAACT1与CcAACT2序列相似性为86.91%,皆由405个氨基酸残基组成。催化活性位点(C99、H361与C391)、底物结合位点(K239与S260)及金属离子结合位点(A256、G257与A259)等都高度保守。C端具有典型的过氧化物酶体定位PTS1-motif (398SAL),但软件预测其可能定位于细胞溶质中。CcHMGS1与CcHMGS2序列相似性为86.50%,皆由474个氨基酸残基组成。活性位点Glu83、Gly107、Cys120、Ala125及第His247等都高度保守,在线软件预测其定位于细胞溶质中。CcHMGR1与CcHMGR2序列相似性为78.32%,分别由560和562个氨基酸残基组成,2个跨膜保守结构域分别位于序列第36~63、34~61和84~107、82~105位,活性位点包括E239、K372、D448和H546等高度保守。MVK、PMK与MVD分别由397、514和423个氨基酸残基组成,三者N端具有典型 的PTS2-motif即10KIILSGEHA、58DVKVTSPQL与44SVTLDPDHL,但软件预测分别定位于细胞溶质、过氧化物酶体和过氧化物酶体之中。CcID11与CcIDI2序相似性为73.45%,分别由296和318个氨基酸残基组成。PTS1-motifs分别位于C端的293~296位和316~318位,软件预测二者亚细胞定位多元化,定位于或过氧化物酶体或质体或线粒体之中。
2.2 橙花叔醇型樟树各组织精油分析结果
有机溶剂萃取5株橙花叔醇型樟树不同组织精油,经GC-MS分析获得各成分的相对含量,具体结果见表2。总计鉴定获得30种单萜类化合物、31种倍半萜类化合物和4种苯丙素类化合物。叶、叶柄、花与花梗等4种精油优势成分为倍半萜类化合物,相对含量分别为58.09%~80.71%、50.14%~71.02%、59.61%~78.48%与56.14%~67.15%,皆超过一半。小枝、树皮精油中优势成分则为单萜类化合物,相对含量分别为37.34%~51.67%与56.33%~78.35%。根的精油优势成分为苯丙素类化合物,相对含量则高达60.22%~75.84%。

表2 樟属各组织精油成分分析结果†Table 2 Analysis of essential oil components of Cinnamomum tissues
叶精油50种成分比较显示,橙花叔醇为第一优势成分,占32.86%~44.39%,其它主要倍半萜成分还包括β-榄香烯、β-石竹烯、a-石竹烯、大根香叶烯D、β-芹子烯、a-芹子烯、橙花叔醇、β-桉叶油醇、石竹烯氧化物与环氧蛇麻烯II等,合计占据精油总量的85.00%以上。别香橙烯、γ-马榄烯、a-布藜烯、a-榄香醇与库贝醇等5种成分为叶精油独有。叶柄精油含40种成分,花柄精油含43种成分,橙花叔醇亦为第一优势成分,含量分别为29.29%~37.37%与22.21%~36.43%。花精油含33种成分,无显著优势成分存在,δ-榄香烯、β-石竹烯、异橙花叔醇、石竹烯氧化物及β-桉叶油醇等成分相对含量占据精油谱前五位。小枝精油含33种成分,1,8-桉叶油素为第一优势成分,相对含量为22.36%~43.42%。树皮精油含28种成分,樟脑为第一优势成分,相对含量为29.31%~40.12%。根精油含33种成分,黄樟油素为第一优势成分,相对含量高达59.01%~74.26%。
橙花叔醇型樟树各组织得油率依次为:根(0.89%~1.19%)、叶(0.37%~0.54%)、叶柄(0.31%~0.42%)、花柄(0.29%~0.35%)、花(0.19%~0.26%)、小枝(0.11%~0.19%)与皮(0.08%~0.15%)。对应每100 g组织倍半萜类化合物含量,分别为根107.36~219.60 mg、叶304.94~415.83 mg、叶柄187.56~256.13 mg、花144.26~185.43 mg、小枝24.79~39.63 mg及树皮13.53~34.71 mg。小枝和树皮中倍半萜总含量远远低于其它组织。本文中将比对质谱数据库中相似度低于<85分的成分归为未确认成分。
2.3 樟树MVA途径部分基因表达具有组织特异性
以Actin基因为内参,比较分析MVA途径系列基因在橙花叔醇型樟树叶、叶柄、花、花柄、小枝、树皮及根等7个组织中的表达情况,结果如图2所示。由图2可知,系列基因分为3类表达模式。第1种模式包括AACT2、HMGR1与PMK等3个基因,即只在一种或两种组织中差异表达,AACT2基因在花、叶柄中下调表达,HMGR1基因在叶柄和小枝中上调表达,PMK基因则在花组织中下调表达。第2种模式包括MVK、HMGS1、MVD与AACT1等4个基因,即无表达组织差异。第3类基因包括HMGR2、HMGS2、IDI1与IDI2等4个基因,此类基因具有明显表达组织特异性。当中,HMGR2、HMGS2与IDI1等3个基因皆在富含倍半萜类化合物组织即叶、叶柄、花与花柄中上调表达。HMGS2基因在叶、叶柄、花、花柄中相对表达量依次是其在小枝中相对表达量的11.25倍、7.85倍、16.94倍与10.27倍。HMGR2基因在叶、叶柄、花、花柄中相对表达量依次是其在小枝中相对表达量的6.70倍、4.44倍、6.06倍与4.68倍。IDI1基因在叶、叶柄、花与花柄中相对表达量依次是其在根中相对表达量的5.76倍、4.84倍、8.63倍与4.75倍。IDI2基因稍有不同,其在叶、叶柄、花梗与小枝中显著上调表达。
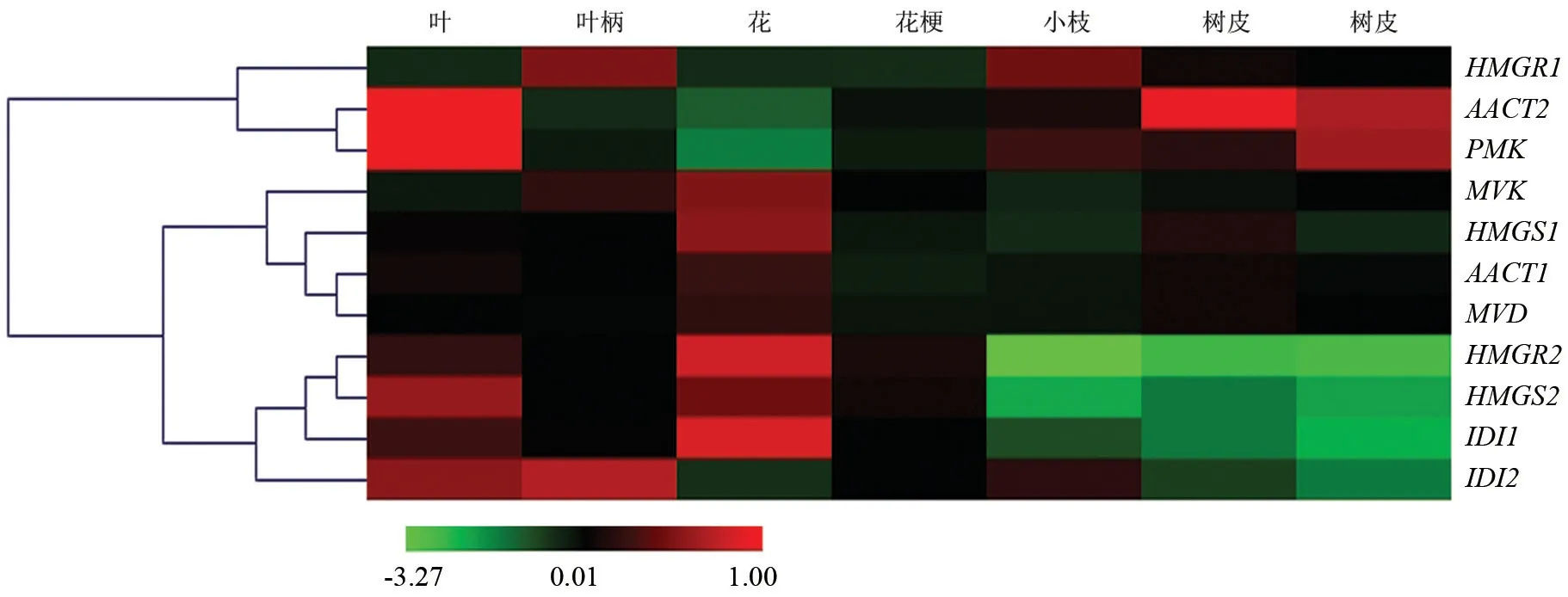
图2 樟树MVA途径系列基因表达组织特异性分析Fig.2 Expression analysis of the MVA pathway among tissues of C.camphora
旁系同源基因之间比较,AACT1基因表达丰度在所有调查组织中都高于AACT2基因表达丰度,相对差异程度介于1.80(树皮)~8.63倍(花)。HMGS2基因在叶、叶柄、花、花柄中表达丰度高于HMGS1基因表达丰度,而在小枝、树皮及根中二者无明显差异。HMGR1基因表达丰度在叶、叶柄、花、花柄中与HMGR2类似,小枝、树皮及根中则显著上调表达,上调幅度介于7.06~14.12倍。除花组织外,IDI1基因在所有组织中相对表达丰度均显著低于IDI2基因表达丰度。
2.4 MVA途径部分基因在倍半萜类樟属植物叶组织中上调表达
2018年7月,采集用于本节分析的桉叶油素型樟树与岩桂叶精油谱(表2)。桉叶油素型樟树叶精油中,单萜类化合物相对含量为81.77%~96.12%,第一优势成分为1,8-桉叶油素;黄樟油素型岩桂中,苯丙烯类化合物相对含量为90.69%~98.43%,第一优势成分为黄樟油素。桉叶油素型樟树和黄樟油素型岩桂叶组织出油率分别为1.55%~1.97%、2.89%~3.54%,对应每100 g鲜叶倍半萜类化合物含量分别为17.36~40.84、21.55~72.16 mg,与橙花叔醇型樟树小枝与树皮精油中倍半萜含量类似。具体比较叶精油倍半萜含量,橙花叔醇型樟树分别是桉叶油素型樟树和黄樟油素型岩桂的10.18~17.33倍及5.76~14.15倍。
2019年4月,提取上述3种化学型樟属植物嫩叶组织total RNA,检测HMGS2、HMGR2、IDI1和IDI2等4个基因的相对表达量(图3)。结果显示,HMGS2、HMGR2和IDI1等3个 基因在不同化学型樟属叶组织中差异性表达。相对桉叶油素型樟树和黄樟油素型岩桂,HMGS2、HMGR2与IDI1基因在橙花叔醇型樟树分别上调表达3.70~3.73倍、4.20~5.46倍及3.38~4.07倍。IDI2基因表达量在不同化学型中无显著性差异。结果显示,HMGS2、HMGR2和IDI1等3个基因上调表达可能是促成倍半萜化学型樟属植物形成的因素之一。

图3 MVA途径系列基因在不同化学型樟属叶组织表达分析Fig.3 Expression analysis of the MVA pathway in different chemical types of Cinnamomum leaves
2.5 橙花叔醇型樟树叶精油倍半萜含量高峰期分析
调查记录1个生长周期内30棵橙花叔醇型樟树不同月份叶精油倍半萜化合物的变化情况。3月至5月为新叶萌发发育期,所有单株叶精油倍半萜含量快速增加。5月至9月,小样本叶精油倍半萜含量变化存在4种模式。第1种模式仅包含编号No.11与No.13两棵单株,5月份倍半萜含量达到高峰期,之后即无明显差异(图4a)。第2种模式包含编号No.1、No.2与No.4等7棵单株,7月份倍半萜含量达到高峰期,之后无明显差异(图4b)。第三种模式包含编号No.14、No.19与No.21等6棵单株,倍半萜含量增加至9月份达到高峰期且与11月含量无明显差异(图4c)。其余15棵单株为第4种模式,9月份叶精油中倍半萜含量达到峰值,显著高于其它月份(图4d)。从精油生产角度看,70%橙花叔醇型樟树叶精油倍半萜含量在9月份最高,是原料林采收的最好时节。
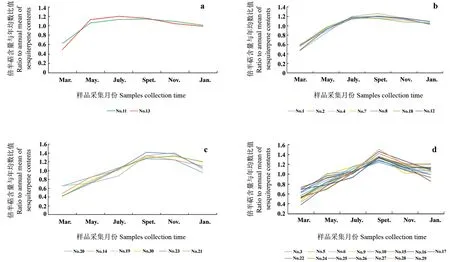
图4 橙花叔醇型樟树叶精油倍半萜含量年变化Fig.4 The annual variation of sesquiterpene contents in the leaf essential oil of trans-nerolidol chemotype C.camphora
小样本3、5、7、9、11与1月份标准差值分别为4.73、9.13、10.36、9.56、8.81与8.48,表明3月份橙花叔醇型樟树群体倍半萜含量变异性最小,7月份最大。7月份,小样本中No.2单株叶精油倍半萜含量最高,每100 g鲜叶含419.02 mg倍半萜,是最低单株(No.24)的2.80倍,也比小样本平均值高57.10%。若统计各成分相对含量,叶精油中所述的10种主要倍半萜类化合物总相对含量受时令影响变化无规律性,各单株情况不一。总体上叶精油中倍半萜相对含量3月份最低,为59.47%,7月份最高,到达68.73%。结果表明,7月份是橙花叔醇型樟树优树选择的最好时机,且橙花叔醇型樟树优树筛选具有很大空间。
2.6 不同时令下橙花叔醇型樟树叶组织MVA途径系列基因表达比较分析
根据2.4的结果,从小样本中挑选6棵代表性单株作为本节实验材料。A组(No.2、No.11、No.13)作为高倍半萜含量代表,B组(No.16、No.21、No.28)作为低倍半萜含量代表,各单株不同月份叶精油倍半萜含量见图5。3—5月,所有单株叶精油倍半萜类化合物皆处于快速积累过程;7—9月,A组叶精油倍半萜类化合物含量变化不显著,而B组叶精油倍化合物含量仍处于增加过程中。

图5 不同月份橙花叔醇型樟树叶精油倍半萜含量Fig.5 Sesquiterpene contents in leaves of trans-nerolidolchemotype C.camphora in different months
借助荧光定量PCR法检测HMGS2、HMGR2、IDI1与IDI2等基因在2019年3、7月各单株叶组织中的表达水平(图6)。设3月No.2叶组织中HMGS2、HMGR2、IDI1与IDI2表达量皆为1,其它单株对应基因相对表达量以与之比值计算,在图中以条形码长度表示。3月,HMGS2、HMGR2、IDI1和IDI2等4个基因在6株测试单株叶组织间皆无显著差异性表达。7月,HMGS2、HMGR2、IDI1和IDI2等4个基因在B组各单株显著性上调表达。

图6 不同时令下橙花叔醇型樟树单株叶组织间MVA系列基因表达分析Fig.6 Expression analysis of the MVA pathway in leaf of trans-nerolidol-chemotype C.camphora in different months
比较不同月份系列基因表达量,A组各单株3月表达丰度显著高于7月,B组各单株表达丰度在3月与7月差异不明显。3月,A、B两组半萜倍处于快速累积过程中,上述4个基因也处于转录活跃期。7月,A组各单株倍半萜含量增加不显著,上述4个基因表达水平也对应地降低。相反,B组各单株在7月还处于快速增加中,上述4个基因表达水平亦保持较高的水平。结果表明:HMGS2、HMGR2、IDI1和IDI2等4个基因表达水平对倍半萜累积速度具有重要调控作用。3—9月,A组倍半萜含量显著高于B组,并未与上述4个基因表达水平情况相关联,显示可能还有其它因素参与调控单株间倍半萜含量差异表型形成。
3 讨 论
在拟南芥基因组中,MVA路径系列基因包括AACT、HMGR、IDI和MVD等由2个旁系同源基因编码,而HMGS、MVK和PMK则由单拷贝基因编码[13]。樟树基因组中除HMGS由2个旁系同源基因编码之外,其它MVA途径系列基因与拟南芥相同。在大部分植物基因组中,HMGS由2个或多个旁系同源基因编码,如在芥菜Brassica juncea基因组中4个HMGS基因已被分离[14]。通过可变剪接或转录起始位点调控,MVA系列基因包括AACT[15]、HMGS[16]、HMGR[17]与MVK[13]等皆存在不同转录本,编码形成不同蛋白亚型,调控类异戊二烯合成或其它生长发育过程。樟树MVA途径系列基因可变剪接或转录起始位点调控目前还一无所知,需在今后开展研究予以证实。
通常认为植物MVA途径发生于细胞质之中,然而实验结果显示部分催化酶亚细胞定位呈多元化。拟南芥AtAACT1具有3种亚型,或定位于过氧化物酶体或细胞溶质中,而AtAACT2具有2种亚型蛋白,或定位于细胞溶质或细胞核中[18]。HMGR蛋白序列N端含有2段疏水跨膜结构域,体外实验显示其蛋白N端锚定于内质网膜腔,C端包含催化中心暴露于细胞质之中,瞬时表达结果则补充证实部分HMGR蛋白亦定位于细胞质中类囊泡结构之中[19]。序列分析显示MVK、PMK与MVD蛋白序列N端皆包含过氧化物酶体定位保守motif-PTS2[20],然而试验结果显示仅PMK与MVD二者定位于过氧化物酶体[21]。IDI序列N端具有质体信号肽同时C端存在另一种过氧化物酶体定位信号肽motif-PTS1,通过形成不同亚型蛋白调控其定位于质体、线粒体或过氧化物酶体中[22]。本文中部分樟树MVA系列酶包括AACT、IDI、MVK、PMK与MVD等皆包含氧化物酶体定位信号肽motif-PTS1或motif-PTS2,在线软件TargetP 1.1、ChloroP 1.1与MitoProt等预测序列基因亚细胞结果基本与其它植物一致,显示植物MVA途径基因区域化调控机制具有高度保守性。
植物MVA途径系列基因表达具有组织特异性,但在不同植物中表达模式略有差异。在不同组织器官中,拟南芥AACT2为组成型表达模式,而AACT1在花序与根中优势表达,但表达丰度远低于AACT2[18]。在巴西橡胶树Hevea brasiliensis中,HMGS基因在茎、叶柄特别在乳汁管细胞中优势表达[23]。在樟科植物山苍子Litsea cubeba中,HMGS基因在根、茎、叶与果中差异性表达,且在不同发育阶段表达丰度差异显著[24]。AtHMGR1S在所有组织器官均可被检测,尤其在早期发育时期和花序中优势表达,而AtHMGR1L与AtHMGR2主要在分生组织与花器官中优势表达且表达丰度显著弱于AtHMGR1S[25]。IDI基因组织表达特异性也常见于多种植物,如在海岛棉Gossypium barbadense茎中优势表达[26]。在本研究所涉及的7个樟树组织中,AACT1、HMGS1、MVK与MVD等4个基因为组成型表达模式,其它基因表达则具有组织特异性。HMGS2、HMGR2与IDI1等3个基因具有相似的表达模式,即都在富含倍半萜类物质的叶、叶柄、花与花梗等中优势表达。此类现象也见于橡胶乳汁管细胞中HMGS与HMGR基因,二者共同优势表达且受乙烯利诱导表达[27]。在番茄Solanum lycopersicumMVA途径研究中也发现HMGS与HMGR同时在腺毛中优势表达[28]。
部分植物MVA途径系列基因转录水平调控是倍半萜类化合物生物合成的重要调控方式之一。拟南芥hmgr1与hmgr2突变体中三萜类化合物分别下降了65%和15%,证明了HMGR对植物萜类合成的重要作用[29]。橡胶性状关联性分析结果显示,优良无性系中HMGS基因表达量、HMGS酶活性与橡胶产量具有正相关性[30]。转基因结果显示,过量表达BjHMGS1、LcHMGS与EuIDI基因可分别提高芥菜甾醇[31]、山苍子倍半萜[24]及杜仲Eucommia ulmoides反式橡胶的产量[32]。在本研究中,HMGS2、HMGR2与IDI1等基因在倍半萜类单株叶组织中相对其它化学型单株优势表达,与倍半萜含量呈正相关性,暗示三者对倍半萜类化合物合成或倍半萜类化学型形成具有重要作用。比较不同倍半萜含量橙花叔醇型樟树单株叶组织,倍半萜累积含量与HMGS2、HMGR2与IDI1等基因表达量无显著相关性,显示可能有其它因素如精油细胞大小、密度或精油合成后转运能力等也参与了调控倍半萜累积过程。
近年来部分樟树MVA途径基因克隆与表达研究亦鉴于其他作者研究报道中,部分基因表达结果与本文结果有所差异[33-35]。由于樟树精油累积量处于动态过程之中,且单株精油含量差异显著,因此在研究基因表达时空变化、组织特异性及化学型之间比较时,取样时间段与取样单株选择尤为重要,不同的单株或取样时间甚至可导致相反的结果。与精油谱数据同步分析可提高基因表达分析结果的可靠性,此外扩大分析样本数亦可提高分析结果的可靠性。本研究涉及表达研究时,出于工作量考虑只挑选了6株代表性单株,所获结果需在下阶段通过增加样本量进一步验证。此外,推定HMGR2、HMGS2与IDI1等基因作为重要调控基因缺少蛋白水平的证据支持。因此,本课题组将比较分析MVA途径各基因启动子序列、寻找调控转录因子及转化模式植物等作为下一阶段研究樟树精油调控机制的主要内容。
