新疆地方绵羊品种Cytb基因 遗传多样性与系统进化分析
2021-05-25吕雪峰许艳丽郑文新赛迪古丽赛买提
吕雪峰,许艳丽,郑文新,胡 昕,赛迪古丽·赛买提,王 乐*
(1.新疆大学生命科学与技术学院,新疆 乌鲁木齐 830000;2.新疆畜牧科学院畜牧业质量标准研究所,新疆 乌鲁木齐 830046)
【研究意义】新疆地处中国西北部,总面积占中国陆地面积六分之一,属于温带大陆性气候,降水量少,气候干燥。天山作为天然屏障将新疆分为南疆和北疆,南疆属暖温带,北疆属中温带,年平均气温10~13 ℃,其中阿勒泰地区冬季气温最低达-50 ℃,吐鲁番地区夏季气温最高达49 ℃,多样的自然环境造就了物种的丰富性。根据《中国畜禽遗传资源志羊志》[1],新疆共有13个地方绵羊品种,这些品种分布在天山南北,各大河流附近,在长期的选育下,逐渐形成了适应当地气候的品种,有着独特的优良性能。如阿勒泰羊、哈萨克羊、巴什拜羊是新疆主要的肉用羊品种,具有体格大、产肉多的特点;多浪羊具有良好的多胎性能;策勒黑羊是我国优良的裘皮用羊;塔什库尔干羊是高海拔环境下良好的肉用品种;和田羊是以生产优质地毯毛而著称的半粗毛羊等。【前人研究进展】地方品种是在特定的地区、特定的地理环境中培育出来的,是我国宝贵的遗传资源。遗传多样性反映了物种所包含基因的丰富程度和它对环境的适应能力。已经有学者通过微卫星对新疆部分绵羊品种的遗传多样性进行了分析[2-5]。由于线粒体 DNA(mtDNA)具有严格的母系遗传、分子量小、突变率高、无组织特异性等优点,目前已成为研究物种进化的重要的分子标记,通过分析线粒体DNA的多态性可以探讨种群的遗传结构、起源、地理分布、种群分歧、系统发育等[6-9]。宁礼群等[10]利用Cytb序列分析新疆10个绵羊品种,发现新疆地方绵羊品种的遗传多样性丰富,至少有3个独立的母系起源。王世峰等通过mtDNA D-loop[11]和ND2[12]基因序列分析了新疆部分绵羊品种的亲缘关系和起源,发现塔什库尔干羊、巴什拜羊、哈萨克羊与东方盘羊关系较近,摩弗伦羊是家绵羊的野生祖先。【本研究切入点】近年来,由于消费市场的需求,新疆引进肉羊品种与本地绵羊品种杂交,同时,因为交通便利,各地方品种交流也非常频繁,导致这些地方品种资源的纯度急剧下降,数量减少。为全面系统地揭示新疆地方绵羊品种资源遗传现状,本研究对新疆13个地方绵羊品种遗传多样性、亲缘关系和系统发育进行了分析。【拟解决的关键问题】为绵羊品种的分类提供合理依据,对品种资源开发、利用和保护提供科学依据。
1 材料与方法
1.1 材料
以新疆13个地方绵羊品种作为主要研究对象,分布地区见图1,以内蒙古乌珠穆沁羊和西藏绵羊做对照,共15个绵羊品种211个个体(表1)。颈静脉采血,抗凝,置于-20 ℃冰箱保存。从NCBI上下载已发表绵羊线粒体胞色素b(cytochrome b,Cytb)基因序列:KF677303.1、KU899149、KU899141;东方盘羊(Ovisvignei)EU366052、东方盘羊乌兹别克斯坦亚种(Ovisammonsevertzovi)EU366057、亚洲摩佛伦羊(Ovisorientalis)KF677307.1、和欧洲摩佛伦羊(Ovisariesmusimon)FR873152作为参照。
1.2 方法
1.2.1 基因组DNA提取 采用上海生工生物工程股份有限公司的EZUP柱式动物基因组DNA试剂盒提取。
1.2.2 引物设计与PCR扩增参考 NCBI 绵羊Cytb基因(Gen Bank登录号:EU176819)序列,用Primer Premier 5.0 软件设计能扩增Cytb基因部分序列的引物(由上海生工股份有限公司合成)。上游引物:5′-ATATACACGCAAACGGGGCA-3′,下游引物:5′-AGGGGGAGTGTTAAGTGGGT-3′,PCR反应体系25 μl:TaqPCR MasterMix试剂12.5 μl,双蒸水H2O29.5 μl,DNA 样品1 μl,上、下游引物各1 μl。PCR扩增程序为94 ℃预变性5 min,94 ℃变性30 s,60 ℃退火30 s,72 ℃延伸30 s,30个循环;72 ℃延伸5 min,4 ℃保存。PCR产物经1 %琼脂糖凝胶电泳检测后送上海生工股份有限公司测序。
1.2.3 数据分析测序结果 根据峰图进行人工校对,利用 Clustal-X 软件对序列进行比对、剪切和拼接,得到Cytb基因部分序列。利用DnaSP v5.10.0.1软件计算各碱基组成比例、单倍型多样度(Hd)、核苷酸多样性(Pi)、平均核苷酸差异数(k)、种群间核苷酸平均差异数(Kxy)、遗传分化指数(Fst)、核苷酸歧义度(Dxy)、遗传分化系数(Gst)和基因交流值(Nm),并进行Tajima’s D 中性检验;利用Network 4.6.1.3软件构建单倍型网络图;利用Arlequin V3.5.1.3 软件进行分子变异分析和错配分析。运用MEGA7.0软件计算碱基转换与颠换平均比率,构建单倍型的系统进化树(neighbor-joining, NJ法,Bootstrap抽样重复次数为1000次)。
2 结果与分析
2.1 PCR扩增结果
以基因组DNA为模板对Cytb基因进行扩增,经电泳检测,得到与预期目标片段大小一致、整齐而清晰的条带,送上海生工股份有限公司测序。
2.2 Cytb序列多态位点
测序结果经ChromasPro拼接后,获得Cytb序列,长度为1017 bp,共检出24个变异位点,其中单碱基变异位点6个,简约信息位点18个(图2),碱基含量平均值A 30.1 %、T 27.3 %、C 29.1 %、G 13.5 %,没有发现碱基缺失或插入现象。其中A+T含量(57.4 %)明显高于C+G含量(42.6 %)。本研究显示转换率明显高于颠换率,总的转换颠换偏倚率R值为25.06,T 和 C 之间的转换率最高,为62.5 %。
2.3 遗传多样性
15个地方绵羊品种单倍型多样度(Hd)在0.396~0.782,群体总Hd为0.684,其中,和田羊的单倍型多样度最低,为0.396,罗布羊的单倍型多样度最高,为0.782,各地方绵羊品种核苷酸多样度Pi在0.00056~0.00593,总群体核苷酸多样度为0.00407,其中,和田羊最低,为0.00056,巴尔楚克羊最高,为0.00593,各地方品种绵羊核苷酸平均差异数k在0.571~6.029,总群体核苷酸平均差异数为0.00407,其中和田羊最低,为0.571,巴尔楚克羊最高,为6.029。 Tajima’s D 中性检测显示,巴尔楚克羊、和田羊和塔什库尔干羊检验结果显著(P<0.05,表1),因此这3个品种可能经历过种群扩张。
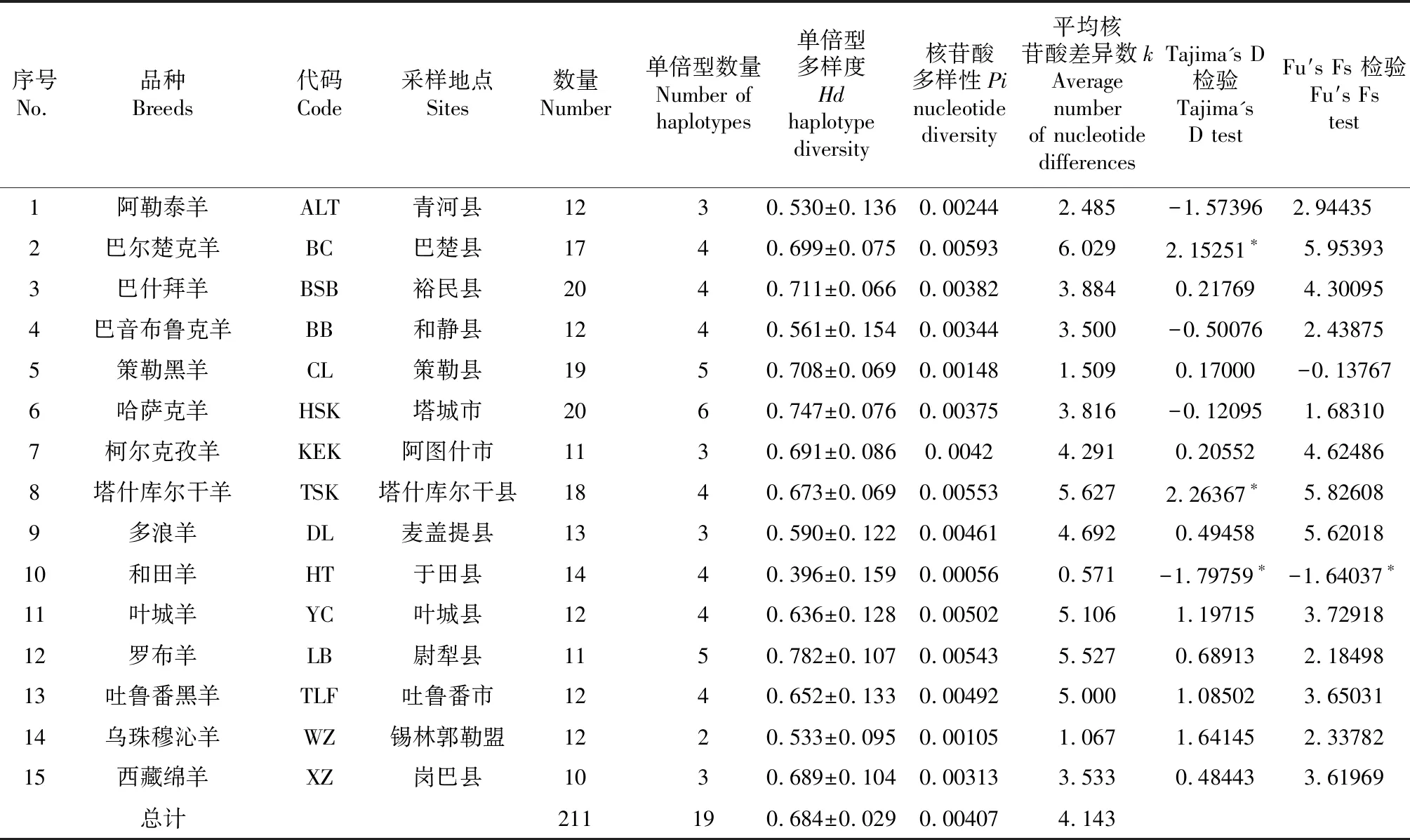
表1 15个绵羊品种遗传多样性参数
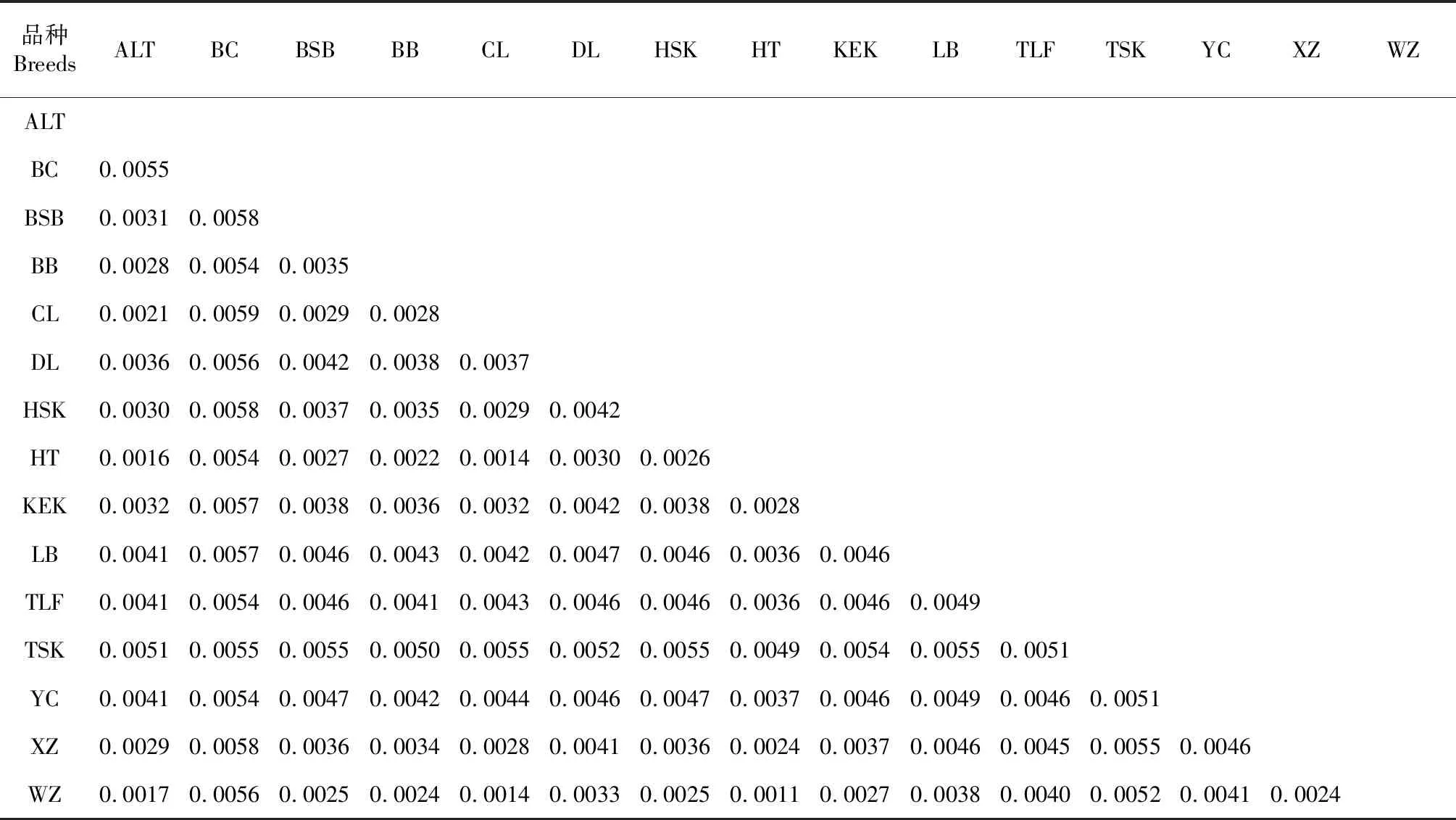
表2 15个绵羊品种群体间遗传距离
2.4 遗传距离和分化
基于 Kimura 双参数模型计算15个绵羊品种内和品种间的遗传距离(表2)。品种内遗传距离在0.0006~0.0060,最小是和田羊和乌珠穆沁羊,为0.0006,最大是巴尔楚克羊,为0.006。品种间遗传距离在0.001~0.006,在新疆13个地方绵羊品种中,和田羊和策勒羊之间的遗传距离最近,为0.0014,巴尔楚克羊和策勒黑羊的遗传距离最远,为0.0059,而且巴尔楚克羊与其它品种羊的遗传距离均较大。各地理种群总遗传分化系数Gst为 0.41938,总固定系数Fst为 0.69561,总基因流Nm为 4.5。各种群间核苷酸差异数Kxy在1.0857~5.9783,均值为4.0767;核苷酸歧义度Dxy在00014~0.0059,均值为0.0040,其中策勒羊和巴尔楚克羊的分歧度最大,达到0.0059;遗传分化系数Gst在-0.0376~0.1017,均值为0.0268;固定系数Fst在-0.0893~0.3882,均值为0.0766。
2.5 单倍型分析
在 211个个体中共发现19种单倍型,其中新疆13个地方绵羊品种有17种单倍型,西藏绵羊单独享有2个单倍型。在19种单倍型中,有 12个共享单倍型(在2个及以上个体中出现)和7个特殊单倍型(仅在1个个体中发现),其中共享单倍型1(Hap1)出现频率最高,占个体的51.66 %(109/211),推测其可能为祖先单倍型,所有的种群均共享该单倍型;其次是单倍型2(Hap2),占个体的16.59 %(35/211,表3)。用19种单倍型构建中介网络图(图3),发现归属于3个不同的单倍型群(A,B和C),分别包括 8、6和5个单倍型。在15个地方绵羊品种中,哈萨克羊单倍型数量最多,有6个单倍型;乌珠穆沁羊单倍型数量最少,仅有2个单倍型。

表3 15个绵羊品种单倍型分布情况
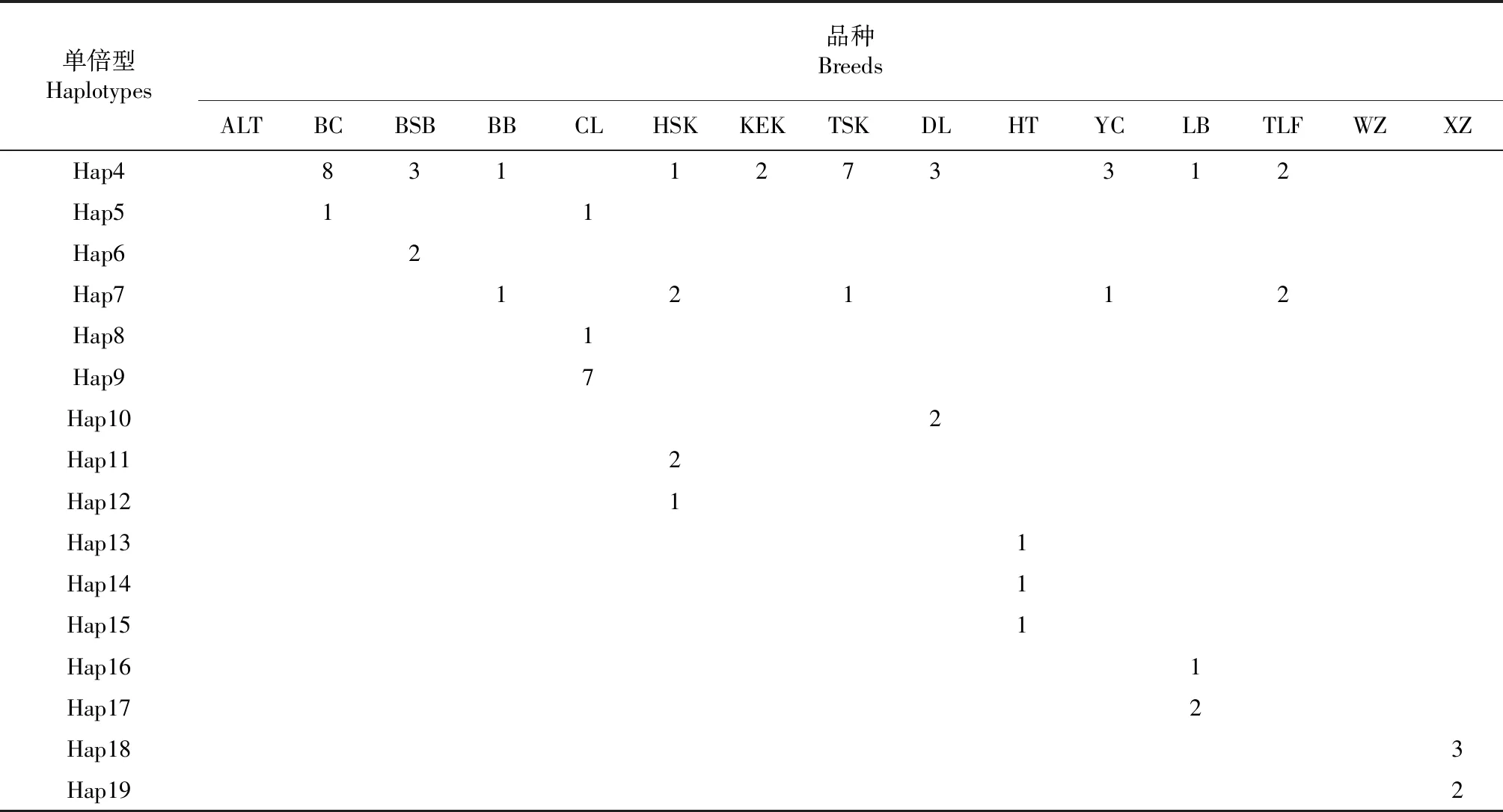
续表3 Continued table 3
2.6 种群变异分析
从表4可知,15个地方绵羊品种种群间的遗传变异(8.68 %)远远小于种群内的遗传变异(91.33 %)。说明,这些地方绵羊品种变异主要来自种群内,种群间的遗传变异较低,品种之间遗传交流非常频繁,品种分化不明显。
2.7 系统进化树构建
品种间遗传距离构建N-J系统进化树(图4)显示,巴尔楚克羊和塔什库尔干羊单独聚为一类,其它13个品种聚为一类。
以邻接法Neighbour-Joining构建19个单倍型和参照绵羊、盘羊群体的系统进化树(图5),19个单倍型聚为3支(A,B,C),单倍型A支包括除和田羊外的14个品种;单倍型B支包括所有的15个品种;单倍型C支包括12个品种,不包括和田羊、乌珠穆沁羊和策勒黑羊。欧洲摩佛伦羊与单倍型A支聚为一支,亚洲摩佛伦羊与单倍型C支聚为一支,这3支与东方盘羊聚在一起,最后再与东方盘羊乌兹别克斯坦亚种聚在一起。

表4 15个绵羊品种的分子变异分析
2.8 种群历史动态分析
Tajima's D 是对地理种群古老突变的检测,Fu′s Fs是对近期发生的种群扩张检测。根据Tajima's D 中性检测结果,巴尔楚克羊、和田羊和塔什库尔干羊可能经历过种群扩张。Fu′s Fs 检验结果中负的Fs值和差异显著的P值被认为种群在历史上有扩张迹象,提示和田羊可能经历过种群扩张。进一步利用Arlequin软件进行核苷酸错配分析,如果期望值是一条平滑的曲线,而观测值有1个单峰,并且中性检验结果显著,说明该种群可能在历史进化过程中经历过种群扩张,如果观察值呈现多峰,说明该群体大小保持相对稳定。对巴尔楚克羊、和田羊和塔什库尔干羊错配分析(图6)发现,巴尔楚克羊和塔什库尔干羊错配分布呈现多峰,且Fu′s Fs 检验结果不显著,说明该群体大小保持相对稳定,未经历种群扩张。和田羊错配分析有1个单峰,Tajima's D中性检验显著,Fu′s Fs检验为负值且差异显著,因此,可认为和田羊群体经历过种群扩张。
3 讨 论
3.1 遗传多样性分析
遗传多样性是指生物体内遗传物质发生变化,可以遗传给后代的变异程度。遗传多样性越高,遗传变异程度越大,该种群适应环境的能力就越强[13]。在新疆13个地方绵羊品种和其它2个品种(内蒙古乌珠穆沁羊和西藏绵羊)mtDNACytb基因序列中共发现了24个变异位点,定义了19个单倍型。这15个地方绵羊品种总的单倍型多样度平均为0.684±0.029,核苷酸多样性为0.00407,根据Grant等[14]将mtDNA序列遗传变异分成的4个类型,罗布羊、叶城羊、塔什库尔干羊和巴尔楚克羊属于高的单倍型多样度(Hd>0.5)和高的核苷酸多样性(Pi>0.005),说明这4个品种遗传多样性丰富,在进化中产生了较多突变,积累了丰富的单倍型和核苷酸变异,也表明该物种对当地的环境具有非常强的适应能力。其中塔什库尔干羊的遗传多样性与王世峰[12]的研究非常一致。阿勒泰羊、哈萨克羊、巴什拜羊、策勒黑羊、吐鲁番黑羊、柯尔克孜羊、多浪羊、巴音布鲁克羊、西藏羊和乌珠穆沁羊共10个品种属于高的单倍型多样度(Hd>0.5)和低的核苷酸多样性(Pi<0.005),说明这些品种也具有丰富的遗传多样性,但是没有足够的时间积累核苷酸多样性[15]。和田羊属于低的单倍型多样度(Hd<0.5)和低的核苷酸多样性(Pi<0.005),说明和田羊种群遗传多样性低,中性检验表明和田羊经历过种群扩张,种群扩张可以快速增加单倍型多样度,但没有足够的时间积累核苷酸的变异。虽然如此,和田羊的单倍型多样度仍然比较低。目前,受市场和消费的影响,我国许多地方绵羊品种的数量在下降[16],真正纯种的和田羊数量也在下降,结合遗传多样性分析,该品种需要加强遗传资源的保护。在15个地方绵羊品种中,单倍型多样度最高的是罗布羊,这是一个生存在塔里木盆地中心的罗布泊地区,极适宜荒漠草场、极端干旱气候、耐粗饲、强抗逆的独特地方羊种,罗布羊的生存地域环境相对封闭,与其它品种羊交流不多,但其遗传多样性丰富,说明其受自然和人工选择压力较小,这可能与当地传统放牧管理、自然交配方式有关,郑永富[17]的研究也证实了这一点。
基因流是一个群体迁移到另一个群体时,将某基因带到新的群体从而产生的基因流动。通常,Nm<1,说明群体可能由于遗传漂变发生了分化[18],Nm>1,说明群体间基因流水平较高,遗传分化较小;Nm>4说明种群间的基因交流更为充分,遗传分化更小[19]。15个绵羊品种总基因流Nm为4.5,表明各品种之间的基因交流非常广泛,这与品种内遗传分化程度较高的结论相对应。分子变异分析也证实了这一点,表明遗传变异主要来自于品种内,而非品种间,也充分表明各品种之间基因交流频繁,没有明显的地理遗传结构。固定系数Fst是一种种群间遗传距离的测度参数[20],可反映种群之间基因的差异程度,根据Weight[21]报道,Fst值在0~0.05表明群体间的遗传分化程度很小,在0.05~0.15表明群体间遗传分化程度中等,在0.15~0.25表明群体间的遗传分化较大,15个绵羊品种平均遗传分化指数Fst为0.0766,表明群体间的遗传分化程度中等。其中和田羊与巴尔楚克羊的遗传分化程度最大,达0.3882;这与两者的遗传距离相一致。
3.2 亲缘关系与进化起源分析
遗传距离分析显示,15个地方绵羊品种中,和田羊与策勒黑羊、乌珠穆沁羊遗传距离最近,与巴尔楚克羊遗传距离最远;阿勒泰羊与和田羊、乌珠穆沁羊遗传距离比较近,与巴尔楚克羊和塔什库尔干羊遗传距离比较远;巴尔楚克羊和塔什库尔干羊与其余13个地方绵羊品种遗传距离都较远。从遗传距离看,和田羊与乌珠穆沁羊的遗传距离比与西藏羊的遗传距离近,说明和田羊与蒙古羊系统有一定的亲缘关系。和田羊主产区与策勒黑羊主产区有重叠,两者之间的遗传距离近,说明这2个品种存在非常近的亲缘关系。策勒黑羊的育成历史不详,大概是19 世纪末,引入黑羔皮羊与当地绵羊杂交形成,推测是与和田羊杂交育成的。塔什库尔干羊分布在新疆帕米尔高原上的塔什库尔干县,与巴基斯坦、阿富汗和苏联3个国家相邻。因此,塔什库尔干羊的育成很可能受周边地区羊(如阿富汗肥尾羊和吉萨尔大尾羊)的影响,因此与其它地方品种遗传距离较远。而巴尔楚克羊则是因巴尔楚克地名而来,据《新疆简史》记载,巴尔楚克命名是在1902年,已知巴尔楚克羊品种的历史有200多年[22],它的育成史可能与塔什库尔干羊或周边国家的绵羊品种有关,值得进一步研究。罗布羊是15个地方绵羊品种中遗传多样性最高的品种,它与多浪羊、巴音布鲁克羊存在系统上的进化关系,与王慧华[23]、袁泽湖[24]的研究结果一致。新疆地方绵羊品种的亲缘与地理分布有明显的关系,从遗传距离构建的进化树可以看出,柯尔克孜羊、巴音布鲁克羊、多浪羊、罗布羊、吐鲁番黑羊、叶城羊亲缘关系均较近,这6个品种,除吐鲁番黑羊外都分布在天山以南,塔克拉玛干沙漠边缘,沿塔里木河流域,地域上的优势使得品种可能互相交流。而吐鲁番黑羊则是以巴音布鲁克羊为父本,哈萨克羊和卡拉库尔羊为母本进行杂交,长期选育而成的[25],因此,与巴音布鲁克羊有更近的亲缘关系。
研究发现,15个地方绵羊品种分为三大支系:支系 A 占所有单倍型42.1 %(8/19),包括欧洲摩佛伦羊和14个绵羊品种中(除和田羊);支系 B占所有单倍型的31.6 %(6/19),在15个地方绵羊品种中均有分布,支系 C占所有单倍型的26.3 %(5/19);包括亚洲摩佛伦羊和12个地方绵羊品种(除策勒黑羊、和田羊和乌珠穆沁羊)。表明新疆地方绵羊品种至少有三大母系起源。关于绵羊起源,一直存在双起源说[26-28]和三起源说[29-31]。本研究结果与三起源说相符,除亚洲摩佛伦羊和欧洲摩佛伦羊两大起源,还有1个不同的母系起源,这也与罗玉柱[32]、刘刚[33]等的研究一致。
对新疆13个地方绵羊品种遗传多样性的分析一定程度上可了解这些品种遗传现状、系统发育和亲缘关系,可为后期地方品种资源保护、利用提供理论依据。
4 结 论
新疆地方绵羊品种除和田羊外,遗传多样性都比较高,和田羊的单倍型多样度最低,为0.396±0.159;罗布羊的单倍型多样度最高,为0.782±0.107。和田羊和策勒黑羊之间的遗传距离最近,巴尔楚克羊和策勒黑羊的遗传距离最远。单倍型网络图显示新疆13个地方绵羊品种有3个不同的母系起源,育成史与地理分布基本一致。和田羊经历过种群扩张,但单倍型多样度仍比较低,提示和田羊需要加强品种的选育和保护。
