水稻bip130与OSA7蛋白互作的验证及bip130对质膜H+-ATPase活性的影响
2021-05-24张雅芬刘亚琴蒋明义
张雅芬,刘亚琴,蒋明义
(南京农业大学生命科学学院,江苏 南京 210095)
植物响应逆境胁迫的有效途径之一为胞内ABA的迅速累积,从而引起一系列的抗逆反应,例如气孔开闭、叶片扩张速度下降、细胞渗透势降低等[1-2]。研究发现,ABA参与质膜ATP酶和液泡膜ATP酶的修饰过程[3],可以为离子的逆向转运蛋白提供更多驱动力。质膜H+-ATPase是广泛存在于植物中的一种膜蛋白。它可以水解ATP产生能量,把H+从细胞内泵出到膜外,使细胞外积累大量正电荷,与细胞内的大量负电荷形成跨膜电化学势梯度,为离子的跨膜运输提供能量来源[4]。H+-ATPase属于P型ATP酶,由 1条多肽链组成,相对分子质量约为1×105,拥有10个跨膜螺旋构成的M-结构域,以及A-、P-、N-、R-这4个胞质结构域。植物对胁迫的耐性提高与植物的质膜H+-ATPase活性增强有关。例如:盐胁迫下质膜H+-ATPase转运H+使细胞膜内外的质子浓度梯度改变,让Na+/H+逆向转运蛋白体将细胞内过多的Na+泵出细胞[5],从而减轻盐胁迫对植物细胞的伤害。质膜H+-ATPase活性会影响一些生理过程,例如:细胞扩张、营养吸收、营养易位、糖易位、气孔控制和激素流动等[6-9]。
水稻质膜H+-ATPase共有10个同源蛋白,OSA7为其中之一。研究表明,OSA7在所有同源基因中表达最高,且对气孔的开放有很大影响;osa7突变体会表现出严重的生长缺陷,在生殖期到达前就会枯萎[10-11]。这些结果说明OSA7在水稻中起着至关重要的作用。bip130最初是由Hirabayashi等[12]从水稻cDNA文库中筛选到的可能与油菜素内酯(BR)受体BRI1互作的蛋白。目前关于bip130的报道甚少,本实验室对bip130进行研究后发现,ABA可诱导水稻中的bip130表达,bip130参与ABA诱导的抗氧化防护途径[13];bip130与促分裂原活化蛋白激酶OsMPK1相互作用[14],而MAPK家族基因广泛参与植物的胁迫响应过程[15];为了进一步探究bip130参与植物胁迫响应的作用机制,本实验室前期利用酵母双杂交技术,以bip130为诱饵筛选到水稻中可能与bip130相互作用的靶蛋白OSA7。然而,酵母双杂交试验具有假阳性。本研究使用多种方法从体内外验证bip130与OSA7相互作用的真实性,并找出bip130与OSA7相互作用的区域以及ABA信号通路下bip130与OSA7的上、下游关系,最后探究bip130是否会调控质膜H+-ATPase的活性,旨在为胁迫条件下bip130的作用机制研究提供理论基础。
1 材料与方法
1.1 材料
蛋白互作试验的植物材料:粳稻(OryzasativaL.)品种‘日本晴’、本生烟(Nicotianabenthamiana)。‘日本晴’、bip130-OE过表达材料以及bip130-RNAi干扰材料用于检测质膜H+-ATPase活性以及RT-qPCR。
1.2 酵母双杂交试验(Y2H)验证蛋白互作
根据pGADT-7图谱和OSA7序列选择KpnⅠ和BamHⅠ酶切位点构建OSA7-pGADT-7重组载体。分别将OSA7-pGADT-7和bip130-pGBKT-7转入酵母Y187和Y2H细胞,于SD-Leu/-Trp上融合培养后,转接到SD-Leu/-Trp液体培养基中振荡培养。收集菌液后滴定到SD-Trp/-Leu/-His/-Ade/X-α-gal四缺培养基上观察菌落是否变蓝。
1.3 萤火虫荧光素酶互补成像系统试验(LCI)验证蛋白互作
根据OSA7序列和pCAMBIA1300-nLUC载体图谱选择BamHⅠ和MluⅠ酶切位点构建重组载体。将pCAMBIA1300∶nLUC-bip130和pCAMBIA1300∶OSA7-nLUC转入GV3101农杆菌后液体培养16~24 h,收集菌液,调整浓度,按体积比1∶1∶1将pCAMBIA1300∶cLUC-bip130、pCAMBIA1300∶OSA7-nLUC以及P19菌液混合后侵染烟草叶片。暗培养24 h后,光照培养2~3 d,喷荧光素酶底物避光反应30 min后,用光学成像仪Tanon 5200观察荧光。
1.4 谷胱甘肽转移酶下拉试验(GST pull-down)验证蛋白互作
根据OSA7序列和pET-30a载体图谱选择KpnⅠ和Hind Ⅲ酶切位点构建重组载体。原核表达并纯化OSA7-N-His、OSA7-C-His以及bip130-GST蛋白,将纯化好的GST-bip130蛋白结合到MagneGSTTM珠子上,分别加入靶蛋白OSA7-N-His和OSA7-C-His,在垂直旋转仪上4 ℃反应1 h。以只含有GST-bip130的MagneGSTTM珠子和携带GST空载体的MagneGSTTM珠子为对照。反应终止后用漂洗缓冲液清洗珠子,使用His抗体进行检测。
1.5 双分子荧光互补试验(BiFC)验证蛋白互作
根据OSA7序列以及pSPYNE载体图谱选择KpnⅠ和BamHⅠ酶切位点构建重组载体。将培养14 d的水稻幼苗的茎部切成小段放入酶解液,抽真空后黑暗低速旋转4 h左右,收集水稻原生质体,在显微镜下观察并计数,将水稻原生质体稀释至(1~2)×106mL-1。将pSPYCE-bip130与pSPYNE-OSA7重组质粒共同转入水稻原生质体,25 ℃暗培养12~16 h后,将细胞置于激光共聚焦显微镜下观察荧光。
1.6 双链RNA的体外合成与纯化
在OSA7的CDS区选择约200 bp的片段作为dsRNA模板,使用RiboMAXTMLarge Scale RNA Production System-T7试剂盒(Promega)合成dsRNA。加入等体积的酚和氯仿抽提并纯化,使用RNA-free水溶解后于紫外分光光度计检测浓度。
1.7 原生质体分离及PEG介导转化
参照Zhang等[16]的方法,提取3叶期的水稻幼苗原生质体:茎部切成约0.5 mm的小段,放入酶解液中黑暗抽真空1 h后,黑暗慢速摇4 h;过滤酶解后的混合液,室温下150g离心5 min,去上清液;用原生质体洗涤液悬浮后,150g离心5 min,去上清液;取适量原生质体溶液滴在血球计数板上,观察原生质体状态和数量;用W5溶液稀释原生质体至(1~2)×106mL-1。取500 μL稀释好的原生质体加入50 μg重组质粒,混合均匀;加入等体积的40% PEG溶液,缓慢混合均匀至无丝状,室温黑暗培养15 min后,加入9倍体积培养溶液,室温静置5 min,150g离心3 min;弃上清液,加入500 μL培养液重新悬浮,室温下150g离心 3 min;弃上清液,转移到用小牛血清润洗过的细胞培养板中,28 ℃黑暗培养12~16 h。
1.8 水稻总RNA提取及cDNA合成
参照Zhu等[17]的方法,将水稻根部加液氮研磨至粉状后加入预冷的600 μL Trizol试剂充分混匀,冰浴 5~10 min;加入200 μL氯仿混匀,冰浴5 min;4 ℃、12 000 r·min-1离心10 min;取400 μL上清液,转入新的1.5 mL EP管中;加入等体积的异丙醇,充分混匀后冰浴30 min;4 ℃、12 000 r·min-1离心10 min,弃上清液;加入400 μL 75%乙醇洗涤沉淀,4 ℃、12 000 r·min-1离心5 min;弃上清液,加入20 μL DEPC溶解总RNA,利用Prime ScriptTM反转录试剂盒(TaKaRa)进行反转录后得到cDNA。
1.9 实时荧光定量PCR
在美国国家生物信息中心网站(NCBI)https://www.ncbi.nlm.nih.gov查询OSA7及其同源基因的序列,用Primer Premier 6设计特异性引物(表1)。参照TaKaRa SYBR GreenⅠ的说明书进行实时荧光定量PCR(RT-qPCR)。总体系为20 μL:正、反引物各0.4 μL,2×SYBRPremixExTaq10 μL,50×ROX Reference DyeⅠ0.4 μL,cDNA模板1 μL,DEPC水7.8 μL。反应程序:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 34 s,95 ℃ 15 s, 40个循环;60 ℃ 1 min;95 ℃ 15 s。

表1 实时荧光定量PCR所用引物Table 1 The primers used to quantitative real-time PCR
1.10 ABA处理下OSA7基因表达检测
将野生型‘日本晴’水稻培养至3叶期后,选取长势一致的幼苗,去伤害处理4 h后用100 μmol·L-1ABA处理水稻幼苗,分别于0、15、30、60、90、120、180和240 min迅速取样,以同等条件下超纯水处理的幼苗作为对照组。提取样品的RNA并反转为cDNA后,进行荧光定量PCR检测OSA7基因表达。以β-actin作为内参,将处理0 min时的相对表达水平作为1。
1.11 OSA7瞬时表达与干扰效果的检测
利用PEG介导转化法将合成的OSA7dsRNA以及pXZP008-OSA7瞬时转入水稻原生质体中,以同等条件下转入ddH2O作为对照组。提取RNA,反转为cDNA后,通过RT-qPCR检测dsRNA的干扰效果以及瞬时转化pXZP008-OSA7后OSA7基因的表达情况。
1.12 bip130与OSA7的上、下游关系检测
以野生型‘日本晴’、bip130过表达突变体以及bip130干扰突变体为材料,用100 μmol·L-1ABA处理30 min后取样,以同等条件下ddH2O处理幼苗作为对照,提取RNA并反转为cDNA后,通过RT-qPCR分析不同材料中OSA7的表达差异;利用原生质体瞬时表达体系,在水稻原生质体中瞬时过表达和沉默OSA7,过夜培养后用10 μmol·L-1ABA处理5 min,以瞬时转入ddH2O作为对照组。提取RNA并反转为cDNA后,通过RT-qPCR分析不同处理中bip130的表达情况。
1.13 质膜H+-ATPase活性测定
参考Larsson等[18]的两相分离法提取和纯化水稻叶片质膜蛋白:取适量水稻根部,加入2倍体积预冷的研磨缓冲液,磨成匀浆后用4层纱布过滤,4 ℃、5 000g离心10 min;取上清液,4 ℃、60 000g离心 30 min;去上清液,用悬浮缓冲液重悬沉淀,加入6.3%的二相系统,上下摇匀40次后,4 ℃、4 200g离心 5 min;取上相和下相继续进入二相系统,分离3次后合并上相,稀释3~5倍;4 ℃、60 000g离心30 min;取沉淀,加入悬浮缓冲液悬浮,参考Bradford[19]的方法测定水稻质膜蛋白含量。
水稻质膜H+-ATPase的活性测定参考Ohnishi等[20]的方法:50 μL质膜微囊加入0.5 mL反应液,放入37 ℃水浴30 min后,加入0.5 mL 10% SDS溶液终止反应,补水至3 mL后,加入3 mL显色液,混匀,放入37 ℃水浴保温20 min,测定波长660 nm处的吸光值,计算无机磷含量。
2 结果与分析
2.1 bip130与OSA7互作关系的验证
酵母双杂交试验结果如图1-A所示,在SD-Trp/-Leu/-His/-Ade/X-α-gal四缺培养基上,试验组OSA7-AD/bip130-BD和阳性对照pGBKT-7-p53/pGADT-7呈现出蓝色菌落,阴性对照pGBKT-7-lam/pGADT-7和OSA7-AD/BD、AD/bip130-BD在四缺培养基上没有生长。结果说明在酵母中OSA7与bip130相互作用。GST pull-down试验结果如图1-B所示,OSA7-N-His蛋白和GST-bip130蛋白相互作用形成了复合蛋白,可以通过Western blot试验检测出OSA7-N-His的条带,而GST空载蛋白不能与OSA7-N-His或OSA7-C-His结合,无法显示出条带,即证明了OSA7的N端与bip130在体外存在相互作用。
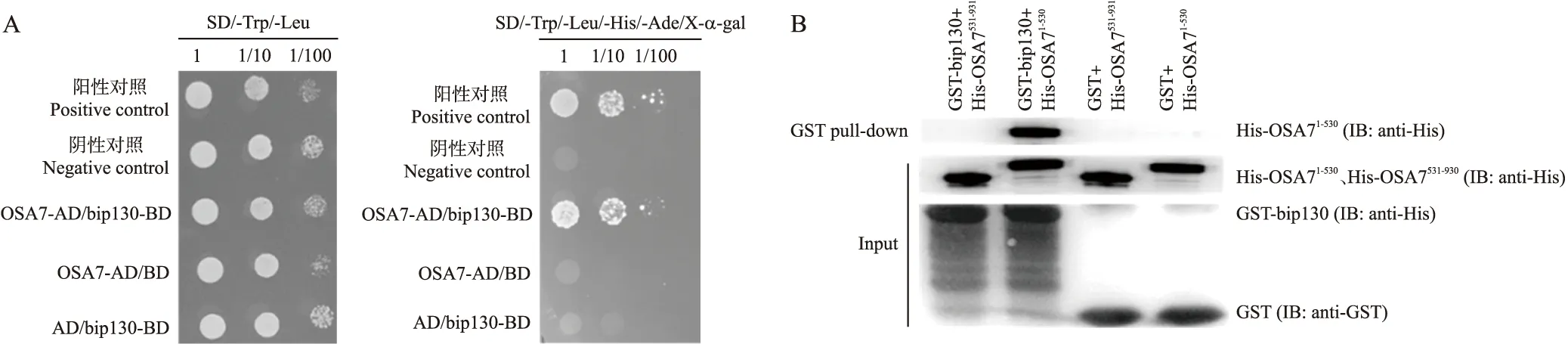
图1 酵母双杂交试验(A)和谷胱甘肽转移酶下拉试验(B)验证bip130与OSA7的相互作用Fig.1 Yeast two-hybird(Y2H,A)and GST pull-down(B)assay of the interaction between bip130 and OSA7
LCI试验结果如图2-A所示:在阳性对照OsDMI3-cLUC+OsPP45-nLUC和试验组bip130-cLUC+OSA7-nLUC共侵染的烟草部位观察到较强的荧光,而阴性对照OSA7-nLUC+cLUC和nLUC+bip130-cLUC没有荧光。这说明bip130与OSA7在烟草内存在相互作用。以水稻原生质体为材料,利用BiFC技术验证bip130与OSA7的相互作用(图2-B),在激光共聚焦显微镜下可以清楚看到bip130-cYFP+OSA7-nYFP共转的水稻原生质体和OsPP45-nYFP+OsDMI3-cYFP阳性对照发出黄色荧光,而阴性对照OSA7-nYFP+cYFP和bip130-cYFP+nYFP都无发光现象,说明OSA7与bip130在原生质体内也相互作用。
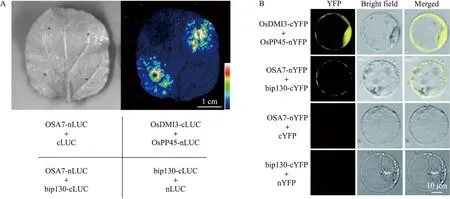
图2 萤火虫荧光素酶互补成像系统试验(A)和双分子荧光互补试验(B)体内验证bip130与OSA7相互作用Fig.2 Firefly luciferase complementary imaging system(LCI,A)and bimolecular fluorescence complementary technology(BiFC,B)assay of the interaction between bip130 and OSA7 in vivo
2.2 bip130与OSA7互作区域的验证
在欧洲生物信息学研究所网站(EBI)分析OSA7的蛋白质结构域,其中13~130 aa、227~321 aa、635~845 aa为跨膜结构域;130~227 aa为A域超家族;323~633 aa为卤酸脱氢酶(HAD)超家族;341~486 aa为胞质N结构域(图3-A)。按照其结构域的成分,将OSA7截段为1~321 aa的跨膜结构域、341~486 aa的胞质N结构域、486~633 aa的HAD超家族域、634~951 aa的跨膜结构域。
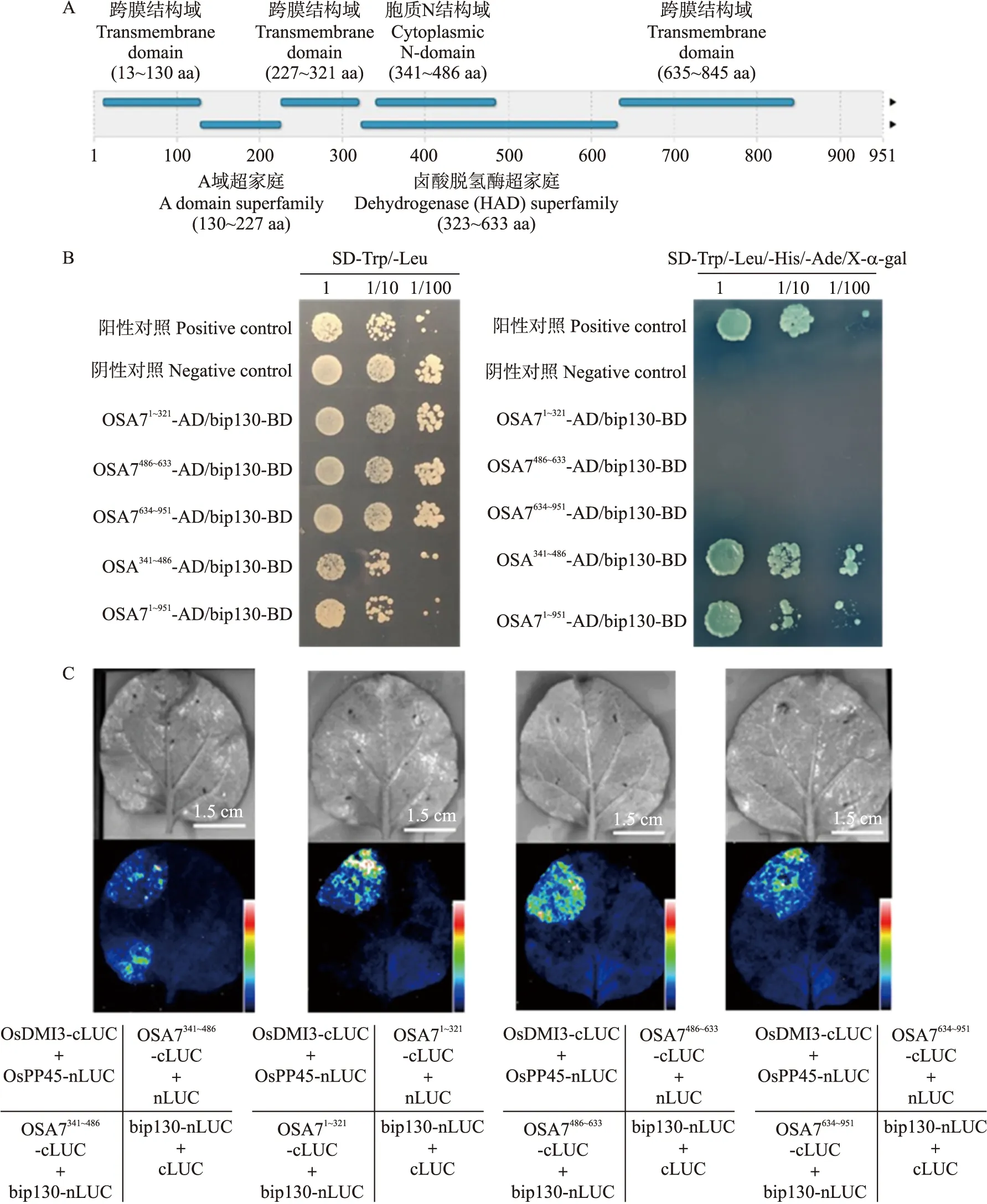
图3 bip130与OSA7互作区域的验证Fig.3 Identification of the interaction region between bip130 and OSA7A. OSA7蛋白结构域分析。B. 酵母双杂交试验体内验证OSA7341~486和bip130的相互作用。C. LCI分析表明烟草叶片中OSA7的胞质N结构域与bip130相互作用。A. The protein structural analysis of OSA7. B. Identification the ineraction between bip130 and OSA7341-486 by yeast two-hybrid in vivo. C. LCI assay shows that the interaction between cytoplasm N-domain of OSA7 and bip130 in tobacco leaves.
酵母双杂交试验结果(图3-B)显示,在SD-Trp/-Leu二缺固体培养基上所有菌落长势良好,说明质粒均正常转入酵母菌中并融合成功。在SD-Trp/-Leu/-His/-Ade/X-α-gal四缺固体培养基上只有OSA7-AD/bip130-BD、OSA7341~486-AD/bip130-BD以及阳性对照BD-P53/AD-SV40长势良好并呈现蓝色,阴性对照BD-Lam/AD-SV40以及其他截段区域与bip130融合的菌落没有生长,表明在酵母体内bip130与OSA7的胞质N结构域相互作用。LCI试验结果(图3-C)显示,4组试验图片中阳性对照OsDMI3-cLUC+OsPP45-nLUC均正常发出荧光,试验组中仅bip130-nLUC+OSA7341~486-cLUC发出荧光,说明在烟草叶片体内bip130作用于OSA7的胞质N结构域。
2.3 bip130与OSA7同源蛋白的关系
为了确定bip130是否也会与OSA7的同源蛋白相互作用,使用DNAMAN软件分析OSA7与其他9个同源基因的氨基酸序列同源性,比对结果见表2和图4。研究发现OSA1—OSA5与OSA7的同源性高达80%以上,因此克隆了OSA1—OSA5的胞质N结构域,利用Y2H试验和LCI试验验证bip130与OSA1—OSA5的关系。

表2 OSA7同源基因的分析Table 2 Analysis of OSA7 homologous genes

图4 OSA7与同源蛋白序列间的胞质N结构域比对分析Fig.4 Contrast analysis of cytoplasmic N-domain between OSA7 and homologous protein sequences
在Y2H试验中,SD-Trp/-Leu二缺固体培养基上所有菌落长势良好,说明质粒均正常转入酵母菌中并融合成功。在SD-Trp/-Leu/-His/-Ade/X-α-gal四缺固体培养基上只有阳性菌落BD-P53/AD-SV40长势良好且呈现蓝色,所有试验组均无法观察到荧光,该结果表明在酵母中OSA1—OSA5的胞质N结构域与bip130不互作(图5-A)。在LCI试验中,仅有阳性对照OsDMI3-cLUC+OsPP45-nLUC观察到较强的荧光,试验组均无荧光,说明OSA1—OSA5的胞质N结构域与bip130在烟草内不存在相互作用(图5-B)。
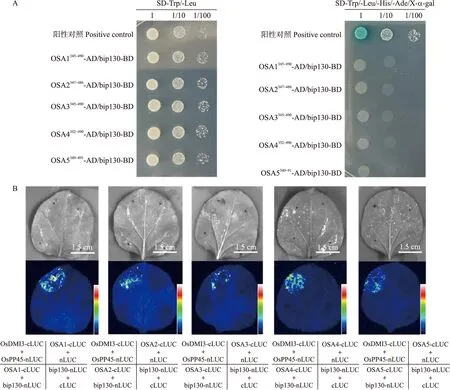
图5 bip130与OSA7同源蛋白的关系Fig.5 The relationship between bip130 and OSA7 homelogous proteinsA. 酵母双杂交技术分析bip130与OSA1—OSA5胞质N结构域的相互作用。B. 萤火虫荧光素酶互补成像系统分析bip130与OSA1—OSA5胞质N结构域的相互作用。A. Yeast two-hybrid analysis of interaction between bip130 and OSA1-OSA5 cytoplasmic N-domain. B. LCI analysis of the interaction between bip130 and OSA1-OSA5 cytoplasmic N-domain.
2.4 外源ABA对OSA7基因表达的影响
从图6可知:ABA可诱导OSA7基因表达,在30 min时到达峰值,4 h后基本回到对照组水平。这说明在水稻根部OSA7基因表达受ABA的诱导上调。
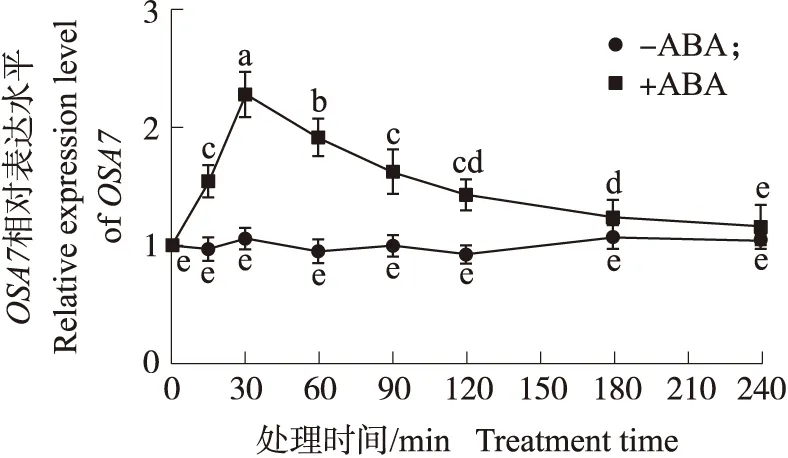
图6 100 μmol·L-1 ABA对水稻根部OSA7基因表达的影响Fig.6 Expression analysis of OSA7 in rice roots exposed to 100 μmol·L-1 ABA treatment不同小写字母表示在0.05水平差异显著(Duncan’s检测法)。下同。Different small letters indicate significant difference at 0.05 level according to Duncan’s multiple range test. The same as follows.
2.5 RT-qPCR分析bip130与OSA7在ABA信号通路中的上、下游关系
2.5.1OSA7的瞬时表达与沉默效果的检测瞬时表达会显著提高OSA7基因表达,是对照组的2.6倍;瞬时沉默效果达到70%,且OSA1—OSA5基因表达几乎不受影响(图7)。因此,可以使用pXZP008-OSA7质粒和合成的dsRNA用于后续原生质体瞬时表达和沉默OSA7的试验。

图7 RT-qPCR分析不同基因在原生质体的瞬时过表达(A)和沉默OSA7效果(B)Fig.7 RT-qPCR analysis of transient expression of different gene in protoplasts(A)and silencing of OSA7(B)A. RT-qPCR分析dsRNA沉默OSA7的效果及沉默OSA7对其同源基因的影响(dsOSA7:原生质体中瞬时沉默OSA7);B. 将pXZP008-OSA7转入原生质体,RT-qPCR分析OSA7的过表达效果(OSA7-OE:原生质体中瞬时过表达OSA7)。Control为野生型水稻原生质体。数据为3个独立试验的平均数。下同。A. RT-qPCR analysis of the effect of dsRNA silencing OSA7 and the effect of silencing OSA7 on its homologous genes(dsOSA7:transient silencing of OSA7 in protoplasts). B. pXZP008-OSA7 was transferred into rice protoplasts and RT-qPCR was used to analyze the expression of OSA7(OSA7-OE:transient expression of OSA7 in protoplasts). Control is wild rice in protoplasts. Values are means±SE of three independent experiments. The same as follows.
2.5.2 ABA信号通路中bip130与OSA7的上、下游关系在bip130过表达材料中,OSA7表达水平明显高于野生型水稻,ABA处理后OSA7表达水平进一步上升;bip130 RNAi突变体中,OSA7表达水平明显低于野生型,ABA处理后OSA7表达水平略有上升。结果表明ABA信号途径中bip130会影响OSA7的表达。在野生型原生质体中瞬时过表达和沉默OSA7后,bip130表达水平与对照组相比无明显差异;ABA处理后无论是瞬时过表达水平还是沉默OSA7,bip130的表达水平与对照组相比均无明显差异(图8)。试验结果表明OSA7不能影响ABA对bip130的诱导。总之,ABA信号途径中bip130位于OSA7的上游。
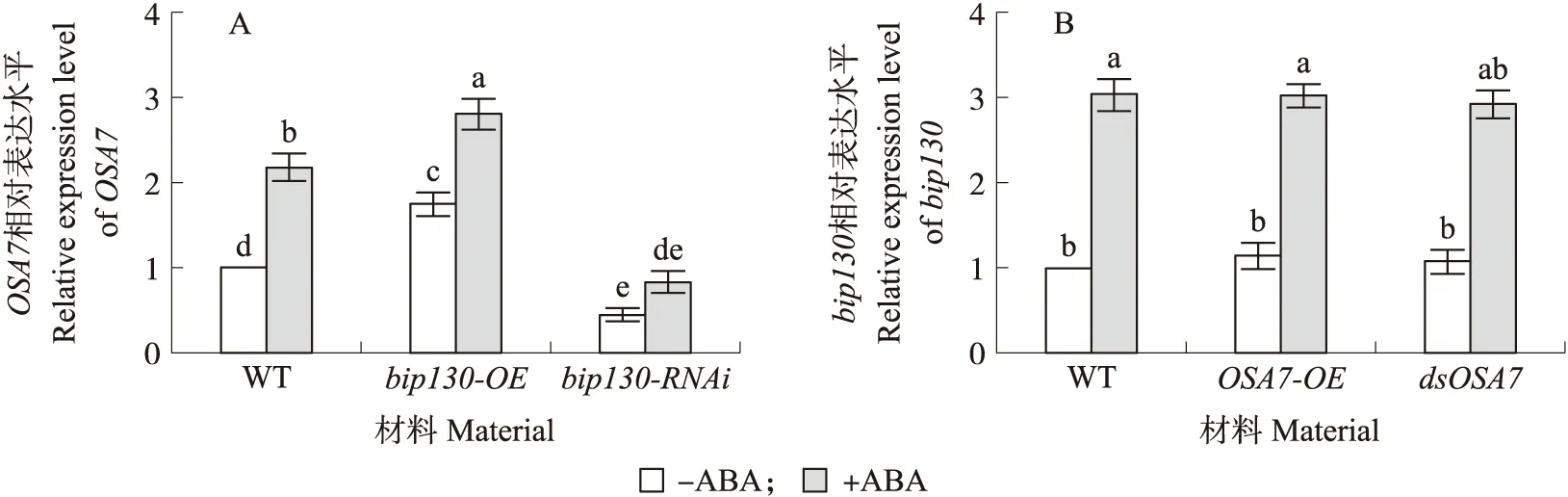
图8 RT-qPCR分析bip130与OSA7的上、下游关系Fig.8 The upstream and downstream relationships of bip130 with OSA7 analyzed by RT-qPCRA. OSA7在bip130突变体中的表达,以野生型‘日本晴’(WT)为对照,bip130-OE为bip130过表达突变体材料,bip130-RNAi为bip130 RNA干扰突变体材料。B. bip130在瞬时过表达和瞬时沉默OSA7的水稻原生质体中的表达水平,以瞬时转入ddH2O作为对照组(以β-actin作为内参)。A. The expression of OSA7 in bip130 mutant,using wild type ‘Nipponbare’(WT)as control(bip130-OE:bip130 overexpression mutant material;bip130-RNAi:bip130 RNA interference mutant material). B. The expression level of bip130 in the protoplasts transiently overexpressing and silencing OSA7,transformation of ddH2O into rice protoplasts as control(using β-actin as an internal reference).
2.6 ABA信号通路中bip130对质膜H+-ATPase活性的影响
以野生型水稻为对照,水处理时,bip130-OE中的质膜H+-ATPase活性增加,ABA处理后,增加更明显;水处理时,bip130-RNAi中的质膜H+-ATPase活性与野生型相比降低,ABA处理后,质膜H+-ATPase活性上升,与未加ABA处理的野生型对比略有升高,但仍比ABA处理后的野生型水稻根部中的质膜H+-ATPase活性低(图9-A)。这些结果说明在ABA信号通路中bip130可以正调控质膜H+-ATPase的活性。
bip130-RNAi材料中的质膜H+-ATPase活性与野生型相比降低,ABA处理后活性增加但比ABA处理后的野生型活性低;与野生型相比,瞬时过表达OSA7后质膜H+-ATPase活性明显增加,ABA处理后活性进一步增加;在bip130-RNAi材料中瞬时过表达OSA7,发现该组质膜H+-ATPase活性介于瞬时过表达OSA7组和bip130-RNAi组之间(图9-B)。与野生型相比,在bip130过表达原生质体中,质膜H+-ATPase活性增加,ABA处理后活性增加更为明显;将OSA7瞬时沉默后,质膜H+-ATPase活性与野生型相比显著下降,ABA处理后活性略有增加但远低于野生型;在bip130过表达材料的原生质体中瞬时沉默OSA7,发现该组质膜H+-ATPase活性与dsOSA7组相比增加,但没有野生型组的活性高,ABA处理后结果相似。这些试验进一步说明在ABA信号途径中bip130通过调控OSA7的活性而影响质膜H+-ATPase的活性。
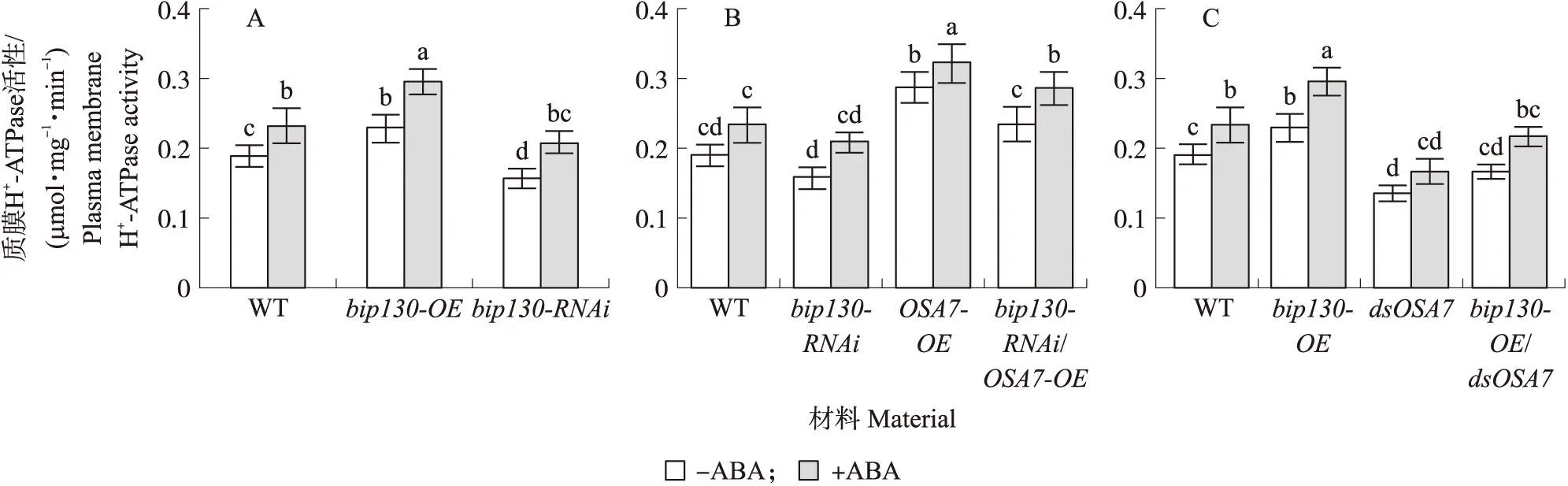
图9 ABA信号途径中bip130对质膜H+-ATPase活性的影响Fig.9 The effect of bip130 on plasma membrane H+-ATPase in ABA signaling pathwayA. 100 μmol·L-1 ABA处理对bip130突变体中质膜H+-ATPase活性的影响。B、C. 在bip130过表达突变体和沉默突变体的水稻原生质体内瞬时过表达和沉默OSA7后检测质膜H+-ATPase活性。A. The effects of 100 μmol·L-1 ABA treatment on the activity of plasma membrane H+-ATPase in bip130-OE and bip130-RNAi mutants. B,C. The activities of plasma membrane H+-ATPase in the bip130 mutant protoplasts transiently overexpress and silencing OSA7.
3 讨论
质膜H+-ATPase在逆境胁迫中具有重要作用。在盐胁迫下,质膜H+-ATPase将Na+主动排除在质外体中,并由质膜和液泡膜中特定的Na+/H+反向转运体催化,使植物在细胞质中维持较低的Na+浓度[5]。在水分胁迫下,水稻根尖积累的ABA能够调节根尖生长素的运输,从而激活质膜H+-ATPase,在根尖释放更多的H+,维持根的伸长与根毛的发育[21]。ABA作为一种逆境激素在植物响应逆境胁迫中起着十分重要的作用。干旱与盐胁迫下ABA可通过降低叶片气孔导度,从而降低植物蒸腾速率以及盐分在根冠的运输,缓解胁迫对植物的损伤[22];ABA还可以提高脯氨酸、甜菜碱、可溶性糖等渗透调节物质的含量[23],增强离子的选择性吸收,调控光合作用,以及加强活性氧物质的清除等方式来调控植物代谢,使细胞由活跃的生长状态向适应胁迫状态转化,最终调控植物适应外界胁迫环境。我们前期研究发现水稻中bip130参与ABA诱导的抗氧化防护途径,且与促丝裂原活化蛋白激酶OsMAPK1相互作用于OsMAPK1的磷酸化激酶结构域[14],而MAPK级联被广泛验证参与ABA的信号转导[24],说明bip130在ABA信号通路中有着重要的作用。本文通过GST pull-down、酵母双杂交技术、萤火虫荧光素酶互补成像系统和双分子荧光互补技术证实bip130与水稻质膜H+-ATPase 7(OSA7)互作的真实性。
质膜H+-ATPase拥有10个跨膜区域,其N端与C端都位于细胞质侧,且拥有一个大胞质环。胞质环可分为3个区域,分别为A、P和N结构域。其中N结构域包含核苷酸结合位点[25]。然而,bip130的功能域未知。为了探究bip130与OSA7具体相互作用的区域,本文分析OSA7蛋白的结构域后将其截段为 4个区域,利用Y2H和LCI验证bip130与OSA7的胞质N结构域相互作用。为了探究bip130是否与OSA7的同源蛋白相互作用,选择与OSA7同源性为80%的5个同源蛋白OSA1—OSA5,并克隆它们的胞质N结构域,利用Y2H和LCI证明它们都不与bip130相互作用。
植物对胁迫的耐性提高与植物的质膜H+-ATPase活性增强有关。质膜H+-ATPase的C末端为自抑制域,H+-ATPase的激活一般是通过C末端倒数第2个苏氨酸磷酸化后与14-3-3蛋白的结合而发生的。除了H+-ATPase的倒数第2个苏氨酸磷酸化外,在拟南芥中AHA2的其他磷酸化位点也可以调节其活性:蛋白激酶5(PKS5)中丝氨酸/苏氨酸Ser931的磷酸化参与AHA2的抑制[26],Thr881参与激活响应蔗糖处理[27]等。有研究发现,第1和第4个跨膜段以及胞质环等区域在调控质膜H+-ATPase活性时也起作用[28],说明H+-ATPase和质子泵活性变化的调节信号并不局限于C末端区域,也可能是通过其他区域的调节起作用。本文验证了OSA7在胞质N结构域与bip130相互作用,且在ABA信号途径中bip130作用于OSA7的上游。利用bip130-OE过表达材料和bip130-RNAi干扰材料对ABA处理下质膜H+-ATPase的活性进行检测,发现bip130-OE过表达植株可进一步增强ABA诱导的质膜H+-ATPase活性,而bip130-RNAi阻止ABA诱导的质膜H+-ATPase活性增加。本文利用水稻原生质体瞬时表达体系在bip130-OE过表达原生质体和bip130-RNAi干扰原生质体中瞬时过表达和沉默OSA7后检测质膜H+-ATPase活性,试验结果表明ABA信号途径中bip130通过调节OSA7的活性而影响质膜H+-ATPase。
综上所述,本试验证明在水稻中bip130仅与10个质膜H+-ATPase中的OSA7相互作用,且作用于OSA7的胞质N结构域。在ABA信号途径中bip130位于OSA7的上游,bip130可通过调控OSA7的活性而影响质膜H+-ATPase的活性。这些研究的开展不仅可以进一步为ABA信号通路中bip130的作用机制提供理论基础,还能使人们对逆境下质膜H+-ATPase活性调控机制有新的认识。
