梨小食心虫细胞色素P450 CYP6AE123 基因的克隆及表达分析
2021-05-24王彪龙马瑞燕高玲玲郭艳琼
韩 慧,王彪龙,李 恩,胡 军,马瑞燕,高玲玲,郭艳琼
(1.山西农业大学植物保护学院,山西太谷030801;2.澳大利亚联邦科学与工业研究组织农业与食品部,文布利西澳6913)
梨小食心虫Grapholita molesta(Busck)属鳞翅目卷蛾科小食心虫属,在全世界广泛分布,主要为害桃、梨等经济作物,导致果品产量和质量严重下降[1-6]。目前,常用的防治手段为化学防治,但长时间的不合理用药导致梨小食心虫对某些杀虫剂敏感度降低[7]。已有不少研究表明,细胞色素P450 在昆虫对杀虫剂的代谢中起着非常重要的作用。
细胞色素P450(Cytochrome P450,CYP)是一类能够代谢昆虫内源化合物(如激素等)和外源性化合物(如植物次生物质、杀虫剂等)的酶系,在昆虫的解毒、细胞代谢和平衡中起着关键作用[8]。CYP 酶的诱导或抑制是昆虫与药物相互作用的基础,它可以使昆虫产生拒食、趋避、中毒等症状。细胞色素P450 具有多个进化支,包括CYP2、CYP3、CYP4 以及线粒体CYP 簇[9],其中CYP3 簇最大,包含CYP6、CYP9 家族在内的多个家族。CYP6 家族属于昆虫特有的家族,不少研究证明,该家族基因参与外源物质的代谢。例如飞蝗CYP6 家族参与了外源杀虫剂的代谢[10],其中CYP6HC1与CYP6HCL1在氨基甲酸酯类和拟除虫菊酯类杀虫剂代谢中起关键性作用[8];冈比亚按蚊CYP6M2基因可对多种不同类别的杀虫剂产生耐药性[11]。此外,目前常用的杀虫剂有阿维菌素、吡虫啉以及高效氯氟氰菊酯等。其中,阿维菌素因其杀虫效果好、对人畜毒性低等优势被广泛应用于梨小食心虫的防治[12]。
本研究从前期构建的转录组数据库进行BLAST 检索,获得一个细胞色素P450 基因序列。通过克隆获得cDNA 全长序列,提交P450 国际命名委员会命名为CYP6AE123,并对其进行生物信息学分析和基因表达模式分析,以明确其不同发育阶段和不同组织部位的表达特性,及其对不同浓度阿维菌素的响应,为进一步研究其功能奠定基础。
1 材料和方法
1.1 试验材料
本试验所用梨小食心虫采自山西省晋中市太谷区西山底桃园,饲养于山西农业大学生物安全与生物防治研究基地人工培养箱中(MGC-450HP),经室内人工饲养50 代以上,光周期15 L∶9 D,相对湿度70%~80%;幼虫饲喂人工饲料,成虫饲喂10%蜂蜜水[13],期间未接触任何杀虫剂。
收集梨小食心虫不同发育阶段样品,成虫3 头、蛹5 头、五龄幼虫5 头、四龄幼虫5 头、三龄幼虫10 头、二龄幼虫30 头、一龄幼虫50 头以及卵100 粒[14]。不同组织选取羽化30 h 之内的健康梨小食心虫成虫,分别取头、前肠、中肠、后肠、脂肪体、马氏管、精巢、卵巢[15],每个重复取60 头成虫。采用阿维菌素LC1(019.82 μg/mL)、LC3(072.90 μg/mL)、LC5(0179.66 μg/mL)分别处理健康的梨小食心虫成虫,24 h 后收集存活的成虫样品。以上材料均放置于液氮中速冻,存于-80 ℃冰箱中备用,以上样品均设置3 次生物学重复。
1.2 梨小食心虫CYP6AE123 克隆
以云杉卷叶蛾CYP6AE83基因为参考序列,对梨小食心虫转录组数据检索、BLAST 比对,获得P450 基因的全长cDNA 序列。利用Primer Preimer 5.0 设计特异性引物(表1)。
根据RNAisoTMPlus 试剂盒说明书提取成虫总RNA,并取1 μg 反转录成cDNA,作为PCR 扩增的模板。PCR 反应体系(50 μL):5×TransStartRFast Pfu Buffer 10 μL、2.5 mmol/L dNTPs 4 μL、上下游引物各1 μL、cDNA(1×)2 μL、TransStartRFast Pfu DNA polymerase 1 μL、ddH2O 31 μL。反应程序:95 ℃2 min;95 ℃20 s,60 ℃20 s,72 ℃1 min,40 个循环;72 ℃5 min。反应产物经1%琼脂糖凝胶电泳后于紫外光下切下目的条带回收纯化。将目的基因连接至pEASY-Blunt 载体中,转化至大肠杆菌DH5α感受态细胞。将其涂布于LB 固体培养基中,挑取单克隆验证,并进行LB 液体培养基过夜培养。菌液验证后送上海生工生物有限公司测序,提交P450 命名委员会进行命名。

表1 引物
1.3 梨小食心虫CYP6AE123 生物信息学分析
P450基因的开放阅读框(openreadingframe,ORF)及氨基酸序列使用ORF Finder(http://www.ncbi.nlm.nih.gov/orffinder)预测;采用ExPASy 的在线工具Prot-Param (http://web.expasy.org/protparam/)预测CYP6AE123 蛋白的理化性质;采用SignalP 4.1 Server(http://www.cbs.dtu.-dk/services/SignalP/)分析信号肽;用NPS@_HNNsecondary[15](https://npsa-prabi.ibcp.fr/cgi-bin/secpred_consensus.pl)预测蛋白质的二级结构;采用SWISS-MODEL(https://swissmodel.expasy.org/interactive)进行蛋白质的三级结构同源建模。
根据NCBI BLASTX(https://blast.ncbi.nlm.nih.gov/Blast.cgi)的比对结果找出同源性较高的其他物种的氨基酸序列,在DNAMAN 进行序列比对;采用MEGA 7.0 软件并运用邻接法(Neighbor Joining,NJ)构建进化树,抽样次数(Bootstrap)设置1 000 次。
1.4 梨小食心虫CYP6AE123 表达水平分析
提取不同发育阶段、不同组织及不同浓度阿维菌素处理后的梨小食心虫总RNA,并取1 μg 为模板反转录为cDNA。以β-actin为内参基因,RT-qPCR检测CYP6AE123基因的相对表达量。每组样品均设置3 次技术重复。反应体系(20 μL):SYBR qPCR Master Mix 10 μL、10 μmol/L 的上下游引物(表1)各0.4 μL、稀释10 倍的cDNA 模板1 μL、去离子水8.2 μL。扩增条件为:94 ℃30 s;94 ℃10 s,50 ℃30 s,72 ℃,30 s,于72 ℃收集荧光,共40 个循环。采用2-ΔΔCT 法[16]计算相对表达量。
1.5 数据分析
使用SPSS 22.0 软件进行单因素方差分析(one way-ANOVA),采用Tukey 法进行差异显著性分析,采用Excel 2016 进行数据处理和绘图。
2 结果与分析
2.1 梨小食心虫CYP6AE123 的克隆及序列分析
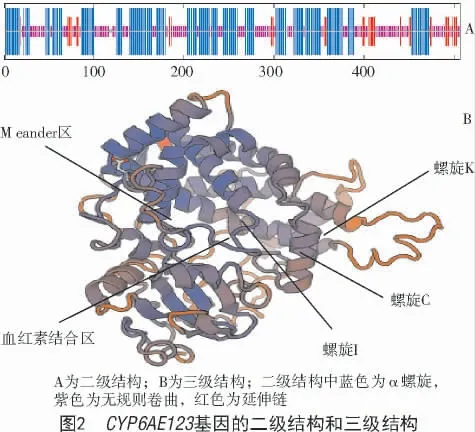
通过检索、比对、克隆和测序,获得梨小食心虫细胞色素P450CYP6AE123的cDNA 全长序列(图1),提交P450 命名委员会,命名为CYP6AE123。ORF 长度为1 524 bp,编码507 个氨基酸;蛋白分子质量为58 176.72 u,理论等电点为7.07,不稳定指数为37.50,故该基因编码的蛋白是稳定的;脂肪系数为89.41,总平均亲水性为-0.110,属疏水性蛋白。二级结构表明,CYP6AE123编码的蛋白主要由α 螺旋(46.55 %),无规则卷曲(42.80%)以及延伸链(8.48 %)组成,三级结构同源建模模拟了P450CYP6AE123的特征序列和空间结构(图2)。

2.2 梨小食心虫CYP6AE123 基因的系统发育分析
BLAST 比对结果表明,梨小食心虫与鳞翅目昆虫苹果蠹蛾Cydia pomonella(Cp)、小线角木蠹蛾Streltzoviella insularis(Si)、棉铃虫Helicoverpa armigera(Ha) 的氨基酸序列一致性分别高达83.67%、57.39%、52.61%(表2)。

表2 梨小食心虫CYP6AE123 的BLAST 比对
多序列比对如图3 所示,梨小食心虫CYP6AE123具有细胞色素P450 的特征区域,包括螺旋I、螺旋C、螺旋K、Meander 区以及血红素结合区。

系统发育分析结果显示(图4),进化树中多为各物种的CYP6AE 家族,表明克隆得到的基因属于CYP6AE 亚家族;且梨小食心虫CYP6AE123与烟草天蛾CYP6AE129聚在一起,表明二者亲缘关系最近,可能具有相似功能。

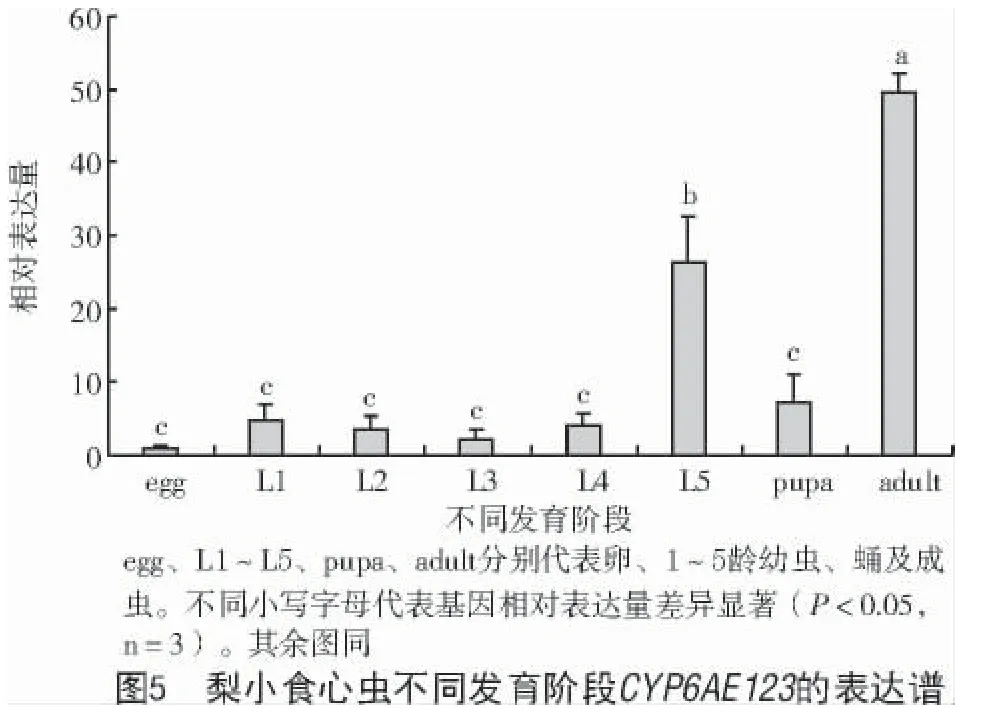
不同发育阶段表达模式如图5 所示,CYP6AE123在梨小食心虫所有发育阶段均有表达,成虫期和五龄幼虫相对较高,且差异达显著水平(P<0.05);其他龄期表达相对较低,且各阶段之间无明显差异。其中,卵期表达量最低,成虫期表达量是卵期的49.20 倍,五龄幼虫时期表达量是卵期的26.92 倍。
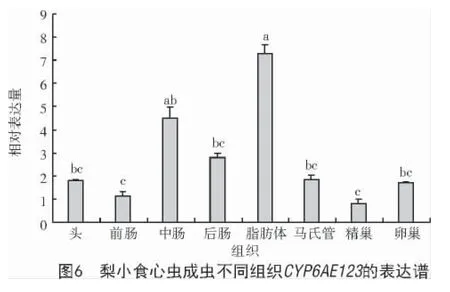
不同组织表达结果表明,CYP6AE123在成虫的脂肪体中表达量最高,中肠次之;前肠和精巢中表达量最低;脂肪体中CYP6AE123表达量为精巢中的8.81 倍、前肠的6.35 倍;中肠表达量是精巢的5.42 倍、前肠的3.90 倍(图6)。
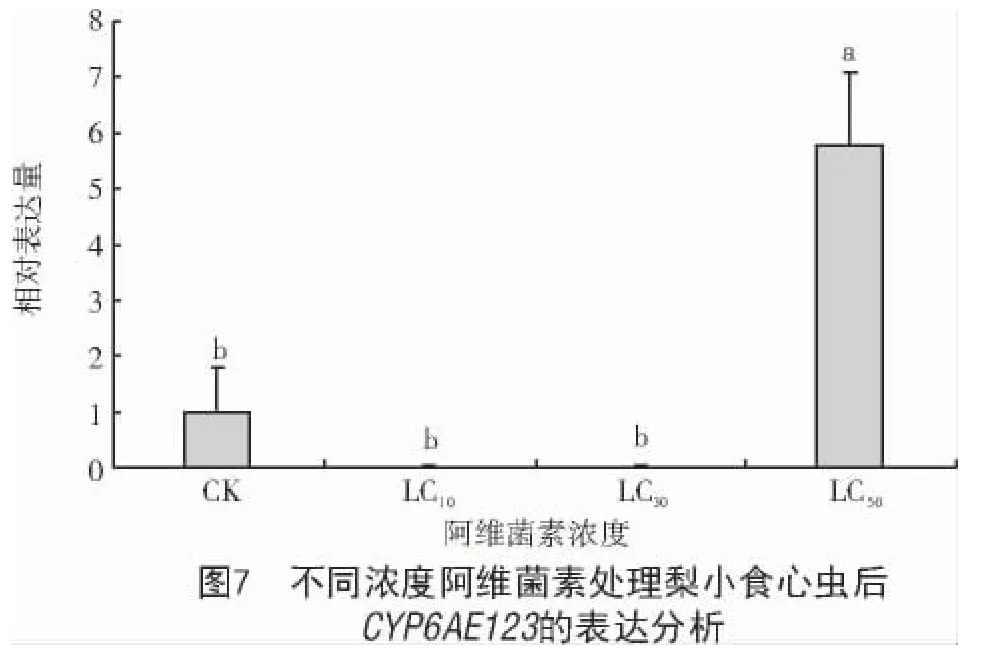
采用不同浓度的阿维菌素对梨小食心虫成虫进行处理,结果表明,低浓度阿维菌素(LC10、LC30)处理后,CYP6AE123表达量较CK 降低,但二者间无显著变化;当浓度增加到LC50(179.66 μg/mL)时,CYP6AE123表达显著增加,为CK 的5.80 倍,且二者间差异达显著水平(P<0.05)(图7)。
3 结论与讨论
本研究对梨小食心虫CYP6AE123的多序列比对结果表明,该基因的氨基酸序列与苹果蠹蛾、小线角木蠹蛾、棉铃虫的氨基酸序列一致性分别高达83.67%、57.39%、52.61%,且均有细胞色素P450 的保守区域,这些保守区域常被用作鉴定P450 的主要特征[17]。
本研究为了进一步研究CYP6AE123的分子特性和生物功能,对不同发育阶段与不同组织中CYP6AE123的表达水平进行了表达模式研究,结果显示,梨小食心虫CYP6AE123在成虫期和五龄幼虫期表达量较高,因为成虫期是梨小食心虫为数不多暴露于环境中的时期,也是农民防治的关键期[18],在这个发育阶段CYP6AE123基因表达量上调,相似的结果在斜纹夜蛾中也有体现[19]。不同组织中基因表达量显示,CYP6AE123基因在脂肪体中表达量最高,中肠次之,可能是因为脂肪体主要用于储存能量以及对外源有毒物质的代谢[20],而中肠是昆虫的消化器官,暗示该基因可能与梨小食心虫外源物质代谢有关。研究发现,杀虫剂对昆虫的诱导表达与基因代谢外源物质密切相关。例如,在氯虫苯甲酰胺处理甜菜夜蛾后,CYP9 家族基因被诱导,进一步验证Unigene0035508 基因参与代谢氯虫苯甲酰胺[21];苹果黄蚜细胞色素P450 基因AcCYP6CY14表达可增强吡虫啉抗性[22]。本研究发现,高浓度阿维菌素(179.66 μg/mL)诱导梨小食心虫CYP6AE123表达,但该基因是否参与梨小食心虫对阿维菌素的代谢尚不明确,下一步将对其进行RNAi,进一步明确其功能。
本研究克隆了梨小食心虫CYP6 家族基因CYP6AE123,该基因属于CYP6AE 亚家族,且包含细胞色素P450 的识别特征;表达分析研究表明,CYP6AE123在成虫期和脂肪体高表达,且被高浓度的阿维菌素诱导表达,推测其可能参与梨小食心虫对阿维菌素的代谢,研究结果可为进一步探究梨小食心虫CYP6AE123基因的功能奠定基础。
