喀纳斯泰加林群落火成演替中草本层与林冠层 物种的连锁关系*
2021-05-21潘存德余戈壁李贵华邹卓颖
郭 珂 潘存德 余戈壁 李贵华 张 帆 邹卓颖 刘 博
(1.新疆农业大学林学与园艺学院 乌鲁木齐 830052; 2.喀纳斯国家自然保护区 布尔津 836600; 3.新疆林业和草原局 乌鲁木齐 830000)
森林群落中的草本层与林冠层的物种关系一直是植被生态学领域研究的热点问题(Barbieretal., 2008; Martinetal., 2011; Songetal., 2014; Rawliketal., 2018; Guoetal., 2019)。充分认识草本层与林冠层的物种关系不仅是探索森林生态系统结构与功能的关键,同时也可为揭示森林生态系统演替过程,实现森林生态系统的稳定和可持续发展提供科学依据(Tilmanetal., 1994)。研究表明,森林群落的草本层与林冠层的物种连锁(linkage)可能有2种方式: 一种是林冠层与草本层物种之间循环往复地相互影响,即林冠层的物种多样性可能通过增加生境异质性或创造有利于更多草本植物物种生存的环境条件来增加草本层的物种多样性(Beatty, 2014; Jagodzińskietal., 2018),包括土壤酸碱度和养分、光照环境或林冠下的凋落物数量与分布等(Hilletal., 2001),同时草本层的物种组成和盖度在一定程度上也决定着林冠层优势树种幼苗的密度和分布(Maguireetal., 1983; Georgeetal., 1999; 杨健等, 2013); 另一种是林冠层和草本层物种对相同环境梯度响应的相似性(Gagnonetal., 1986; Robertsetal., 1988; Hostetal., 1992; Nematietal., 1995),且二者物种之间的连锁与干扰有关,会在森林演替的过程中发生变化(Gilliametal., 1995)。
泰加(taiga)一词最初来自俄语,原指北极苔原以南与苔原接壤的极地针叶林地带(Hoffmann, 1958),现在这一术语泛指寒温带的北方森林(Esseenetal., 1997)。我国真正意义上的泰加林分布区主要包括大兴安岭北部及其支脉伊勒呼里山山地(吴征镒, 1980),以及阿尔泰山(中国)北端的喀纳斯区域(中国科学院新疆综合考察队等, 1978)。喀纳斯区域的森林作为我国唯一的西西伯利亚山地南泰加林生态系统的代表,同分布于世界上寒温带地区的其他山地森林一样,自然火干扰是其群落演替和生生不息的主要驱动力,对森林生态系统以生物多样性为本质体现的生态完整性维持具有不可或缺的作用(刘翠玲, 2009; Chavezetal., 2012; Yuetal., 2013; Schereretal., 2018)。在火干扰驱动下,喀纳斯泰加林群落物种多样性的维持有赖于群落演替过程中草本层物种与林冠层树种的协同共生(Esseenetal., 1997; Reichetal., 2012; Bartelsetal., 2015)。目前,关于喀纳斯泰加林的研究大多集中在对其植被特点及植物区系形成(潘晓玲等, 1994)、森林群落物种多样性特征(张荟荟, 2008)和火成演替森林群落类型(郭珂等, 2019; 杨玉萍等, 2019)等方面,但有关火干扰后,泰加林群落演替过程中草本层与林冠层物种之间是否存在连锁关系——对相同环境梯度响应的相似性,尚未见报道。
笔者以喀纳斯泰加林火成演替群落为对象,在进行数量分类和演替阶段划分的基础上(郭珂等, 2019),采用典范对应分析(canonical correspondence analysis,CCA)和相关分析,探讨不同烈度火干扰后泰加林群落演替过程中草本层与林冠层物种关系及其成因,检验泰加林群落火成演替过程中的草本层与林冠层物种是否存在连锁关系,以及其连锁关系的形成机制,以期加深对泰加林群落草本层与林冠层物种关系的认识。
1 研究区概况
喀纳斯国家级自然保护区位于新疆阿勒泰地区布尔津县境内北端(86°54′—87°54′E,48°35′—49°11′N),为中国与哈萨克斯坦、俄罗斯、蒙古国接壤地带,其西部与白哈巴河县交界,南部与布尔津县的禾木乡相连,是我国唯一的古北界欧洲-西伯利亚动植物区系典型分布区。保护区东西长约74 km,南北宽约66 km,总面积2 201.62 km2,其中核心区面积705.20 km2,缓冲区823.42 km2,科学试验区673.00 km2,本研究的数据采集在科学试验区(87°01′45″—87°33′50″E,48°36′18″—48°38′56″N)进行。保护区具有明显的温带高寒山区气候特征,春秋温暖,冬季严寒漫长,全年无夏季。年均气温-0.2 ℃,极端最高气温29.3 ℃,极端最低气温-37.0 ℃,气温年较差31.9 ℃。年均降水量为1 065.4 mm,年均蒸发量为1 097.0 mm,相对湿度一般为59%~90%,全年日照时数为2 157.4 h,无霜期80~108 天,常年盛行西南风,最大风力可达8级。保护区独特的地貌和气候特征,孕育了其以西伯利亚山地泰加林为主的森林生态系统,森林群落乔木层优势种主要有新疆落叶松(Larixsibirica)、新疆云杉(Piceaobovata)、新疆五针松(Pinussibirica)和垂枝桦(Betulapendula)等; 草本层优势种主要有黑穗薹草(Carexatrata)、老芒麦(Elymussibiricus)、疏花卷耳(Cerastiumpauciflorum)和细叶野豌豆(Viciatenuifolia)等。
2 研究方法
2.1 样地设置与调查
森林群落调查采用典型样地法。分别于2016、2017年的6月中旬—8月中旬,在喀纳斯国家级自然保护区科学实验区尚未受到人为干扰的可识别的历史火干扰林分中,依据火疤木,共设置30 m×30 m的典型样地369个,样地边界距林缘至少50 m。在每个典型样地的4角和中心点嵌套设置5个1 m×1 m的草本样方。为避免因林木火斑造成的自然火干扰误判,典型样地设置的林分条件为面积不小于1.0 hm2,样地及周边存在5株及5株以上新疆落叶松火疤木(fire-scarred tree),且火疤木距离调查年份(2016、2017年)最近一次成疤年龄相同。
对样地中乔木树种(高度>1.3 m)进行每木调查,记录树种名、高度、胸径和生长状况,用统计法测定林冠层郁闭度(crown density,Pc),用LAI-2200C冠层分析仪(美国LI-COR)测定样地林冠层的叶面积指数(leaf area index,LAI),并根据每木调查数据计算林分平均胸径(average diameter at breast height,DBH)、林分株数密度(density of trees,Dt)和林分平均高(average height of stand,HD); 在草本样方中,对草本植物采用每株调查法,记录维管束植物的种名、高度、盖度和数量。土壤采样点设在每个典型样地内,沿坡体方向分别在上、中、下各挖1个土壤剖面,记录土壤剖面特征(分层和结构)。在每个土壤剖面用木质抹刀,按0~10、10~25、25~70 cm的土层采样,确保上下取样均匀; 土样风干后,将每个样地3个剖面的土样分3层混合,用四分法取得土壤样品,去除枯枝、树根、枯叶和石砾等杂物,并研磨过筛(100目)后用于土壤理化性质测定。另外,用100 cm3的土壤环刀,自上述3层剖面每层的中央采样并装入铝盒密封,用于测定土壤密度(soil bulk density,SBD)。
2.2 火干扰因子、地形因子指标获取
火干扰因子包括火烈度(fire severity,Seve)和火后时间(post-fire time,PFT)。火烈度是火干扰造成的森林生态系统地上和地下有机物损耗程度的表征(Keeley, 2009)。本研究通过调查林木火疤的外在属性(长度、高度、深度、成疤部位)和抗火性强的新疆落叶松活立木的残留比例,确定林分历史火干扰的火烈度(强、中、弱),并依据调查时间(年份)和火疤木形成层与木炭层之间的年轮数,推算群落的火后时间(刘翠玲, 2009)。根据调查结果,有强烈度、中烈度、弱烈度火干扰样地64、141、164个; 火后时间在16~179年。地形因子包括海拔(elevation,Elev)、坡度(slope,Slop)和坡向等。用GPS记录样地经纬度和海拔,用罗盘仪测量坡度和坡向。
2.3 土壤理化性质测定
土壤理化性质测定采用国家标准法或行业标准(鲍士旦, 2000),土壤密度(SBD)和孔隙度(soil porosity,Poro)均采用烘干法(NY/T 1121.4—2006); 电导率(conductivity,Con)采用电位法(HJ 802—2016); 酸碱度(pH值)采用电位法(GB 7859—87); 土壤有机质(soil organic,Org)含量采用重铬酸钾氧化-外加热法(GB 7857—87); 全氮(total nitrogen,TN)含量采用半微量开氏法(GB 7173—87); 全磷(total phosphorus,TP)含量采用钼锑抗比色法(GB 7852—87); 全钾(total potassium,TK)含量采用火焰光度法(GB 7854—87); 碱解氮(available nitrogen,AN)含量采用碱解扩散法(GB 7849—87); 有效磷(available phosphorus,AP)含量采用钼蓝法(GB 7853—87); 速效钾(available potassium,AK)含量采用NH4OAc火焰光度法(GB 7856-87); 微量元素钙(GB 7873—87, Ca)、镁(GB 7873—87, Mg)、铁(GB 7873—87, Fe)、锰(GB 7873—87, Mn)、铜(GB/T 17138—1997, Cu)和锌(GB/T 17138—1997, Zn)均采用DTPA-TEA浸提-AAS法。
2.4 数据处理与分析
在对369个样地进行数量分类和演替阶段划分的基础上(郭珂等, 2019),进一步分析不同烈度火干扰后泰加林群落演替过程中的草本层与林冠层物种连锁关系及其成因。
2.4.1 群落多样性指数计算 根据喀纳斯国家级自然保护区泰加林火成演替群落的物种多样性特点(张荟荟, 2008),选取Patrick指数、Shannon-Wiener指数、Simpson指数和Pielou指数,对草本层和林冠层的物种丰富度、多样性、优势度和均匀度进行测度。各指数计算公式如下
1)物种丰富度Patrick指数(S)(Patricketal., 1963):
S=物种数。
(1)
2)物种多样性Shannon-Wiener指数(H′)(Magurran, 1988):
(2)
3)物种优势度Simpson指数(D)(Simpson, 1949):
(3)
4)物种均匀度Pielou指数(E)(Pielou, 1969):
(4)
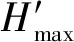
2.4.2 物种连锁关系 CCA除了能生成带有样地和物种位置的排序图外,还能生成来自排序空间中心的环境向量。这些向量的长度表示每个被测环境变量的梯度长度。向量长度与环境梯度在解释物种格局中的重要性时成正比,随向量长度由短到长,表示环境梯度从不太重要转为更加重要。此外,排序空间中的点彼此相距越近,表示样地的物种组成相似性越高。如果样地的草本层与林冠层在第一排序轴的得分值之间存在显著相关性,则表明草本层和林冠层的物种组成在空间变化模式和程度上有密切相似性,即存在基于相同环境变量的连锁关系(Palmer, 1993)。因此,森林群落草本层和林冠层是否以类似的方式响应环境变量,可分别通过对森林群落不同演替阶段草本层和林冠层进行CCA排序来检验(Gilliametal., 2014)。
对不同火烈度干扰后不同演替阶段的森林群落的林冠层和草本层,分别进行单独的CCA排序。在所有情况下,CCA排序第一轴解释了物种多样性数据的最大变异性,因此,CCA排序第一轴得分值被用于比较林冠层与草本层及检验二者间的相关性。所绘制的相关图测量了林冠层和草本层植被排序中沿着第一轴排列的样地的相似性。CCA中的响应变量为草本层和林冠层的物种多样性指数,解释变量由地形、土壤和林分因子组成,共包括26个因子。地形因子中,样地所处坡向在进入CCA前需进行转换。坡向方位角的转换是将0~360°的罗盘测量方位角转换为0~1之间的坡向指标值(transformation of aspect,TRASP)(Robertsetal., 1989),转换公式如下:
(5)
式中,aspect为坡向方位角角度。
通过转换,TRASP的值变化为0~1,0代表北北东方向,1代表南南西方向,即值越大表示生境越干热。CCA中土壤因子数据均采用各层测定数据以各层厚度为权重的加权平均值数据。
应用Pearson相关,计算森林群落在不同火成演替阶段的草本层与林冠层多样性指数间的相关系数,采用双侧方法检验多样性指数间的相关性。采用一元线性回归(简单相关)和F检验,检验不同火烈度和火成演替阶段森林群落的草本层与林冠层CCA排序第一轴得分值的相关性。相关性F检验的显著性水平P值越小,表明连锁关系的紧密程度越高。
数据整理采用Microsoft Excel 2003; 数据分析采用CANOCO Version 5.0和SPSS 20.0软件; 绘图应用Origin 2018软件。
3 结果与分析
3.1 火成演替中的草本层与林冠层物种连锁关系
不同烈度火干扰后,泰加林群落草本层与林冠层存在不同程度的物种连锁关系,但显著水平随火烈度增强而降低(图1)。弱烈度火干扰后,草本层与林冠层的CCA排序第一轴得分值呈正相关,相关系数为0.120,且相关性达到显著水平(P=0.017<0.05); 中烈度火干扰后,草本层与林冠层的CCA排序第一轴得分值呈正相关,相关系数为0.065,且相关性达到显著水平(P=0.043<0.05),但低于弱烈度火干扰情况; 强烈度火干扰后,草本层与林冠层CCA排序第一轴得分值呈负相关,相关系数为-0.116,未达显著水平(P=0.093>0.05)。
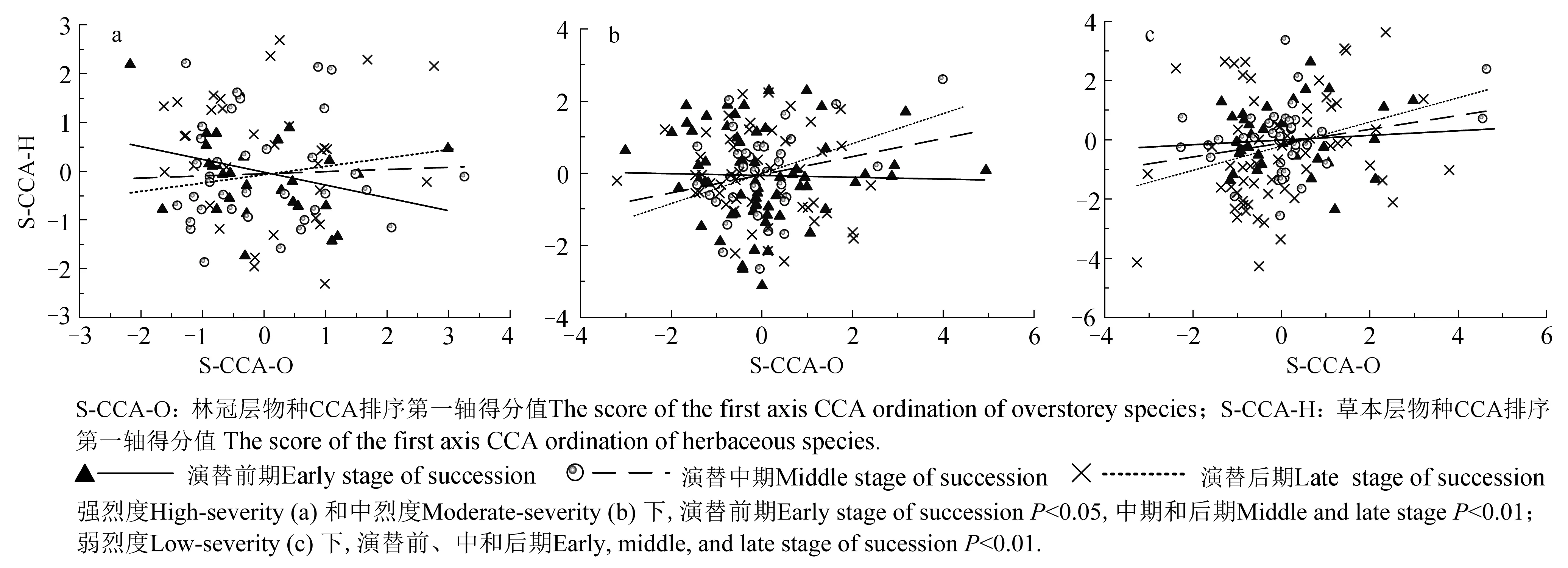
图2 强烈度(a)、中烈度(b)、弱烈度(c)火干扰后不同演替阶段的草本层和林冠层物种连锁关系Fig. 2 The linkage between herbaceous layer and overstorey species at different succession stages after high-severity fire(a), moderate-severity fire (b) and low-severity fire(c)
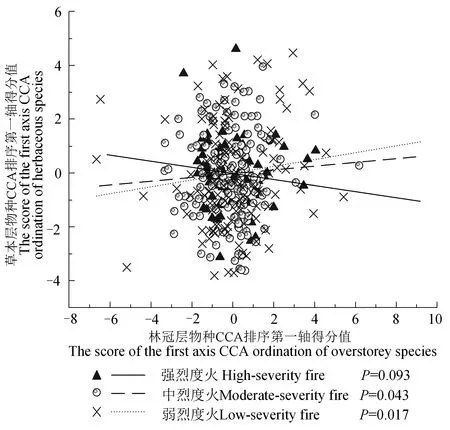
图1 不同烈度火干扰后草本层与林冠层物种连锁关系Fig. 1 The linkage between herbaceous layer and overstorey species after different fire severity
3.2 不同烈度火干扰后不同演替阶段的草本层与林冠层物种连锁关系
强、中、弱烈度火干扰后,草本层与林冠层在不同演替阶段均存在物种连锁关系,且其显著性水平均随演替向前发展逐渐升高(图2)。强烈度和中烈度火干扰后,演替前期,草本层与林冠层CCA排序第一轴得分值呈负相关,相关系数分别为-0.265和-0.027,达到显著水平(P=0.044<0.05;P=0.043<0.05); 演替中期,二者得分值呈正相关,相关系数为0.037和0.301,且达到极显著水平(P=0.008<0.01;P=0.006<0.01); 演替后期,二者得分值呈正相关,相关系数为0.079和0.374,达到极显著水平(P=0.006<0.01;P=0.004<0.01),且显著性水平高于演替中期。弱烈度火干扰后,草本层与林冠层CCA排序第一轴得分值在演替前、中、后期均呈正相关,相关系数分别为0.082、0.188、0.274,均达到极显著水平(P=0.006<0.01;P=0.005<0.01;P=0.002<0.01),且显著性水平增加。
3.3 不同火烈度干扰后不同演替阶段的草本层与林冠层物种连锁关系的成因
对草本层物种多样性有重要影响且相关性达到显著水平(P<0.05)的因子,在强烈度火干扰后的演替前期有Elev、pH值、Poro、Dt和土壤Org、TP、Cu、Mg、Ca含量(图3a、表1),中期有Elev、pH值、Poro、SBD和土壤Cu、Zn、AK、Ca、TK含量(图3b、表1),后期有pH值、Pc和土壤TN、TP、AK、Cu、Fe含量(图3c、表1); 在中烈度火干扰后的演替前期有Elev、pH值、Con、Poro、Pc、LAI和土壤TK、TN、AN、Org、Cu、Mg含量(图5a、表2),中期有Elev和土壤TN、AN、AP、TK、Mg、Ca含量(图5b、表2),后期有Elev、Con和土壤TK、TN、TP、Fe、Cu含量(图5c、表2); 在弱烈度火干扰后的演替前期有Elev、PFT、Dt和土壤TK、AN、Zn、Ca含量(图7a、表3),中期有Elev、Poro、SBD、Dt和土壤Org、TK、Cu、Zn、Mg含量(图7b、表3),后期有Elev、PFT、Poro、HD和土壤TK、AP、Cu、Zn含量(图7c、表3)。
对林冠层物种多样性有重要影响且相关性达到显著水平(P<0.05)的因子,在强烈度火干扰后的演替前期有DBH、HD和土壤TN、AK、AP、Cu、Mn含量(图4a、表1),中期有土壤TN、AK、AP、Cu、Mn含量(图4b,表1),后期有LAI、Pc和土壤TN、TP、AN、Fe、Zn含量(图4c、表1); 在中烈度火干扰后的演替前期有Elev、Poro和土壤TN、Zn、Ca、Fe含量(图6a、表2),中期有Elev、Poro、DBH、HD和土壤TN、AP、Org、Fe、Mg含量(图6b、表2),后期有Elev和土壤Org、TN、TP、TK、AN、AP、Zn、Fe含量(图6c、表2); 在弱烈度火干扰后的演替前期有Elev、Slop、TRASP、PFT、Con、Dt、HD和土壤AK、Fe含量(图8a、表3),中期有Elev、Con、Dt、HD和土壤Org、AN、Cu含量(图8b、表3),后期有Elev、HD和土壤Org、TN、TK、AP、Cu含量(图8c、表3)。
由此可知,草本层和林冠层物种多样性具有一致性响应且达到显著性水平的因子,在强烈度火干扰后的演替前期只有土壤Cu含量1个因子,中期有土壤AK和Cu含量2个因子,后期有Pc和土壤TN、TP、Fe含量4个因子; 在中烈度火干扰后的演替前期有Elev、Poro和土壤TN含量3个因子,中期有Elev和土壤TN、AP、Mg含量4个因子,后期有Elev和土壤TN、TK、TP、Fe含量5个因子; 在弱烈度火干扰后的演替前期有Elev、PFT和Dt3个因子,中期有Elev、Dt和土壤Org、Cu含量4个因子,后期有Elev、HD和土壤TK、AP、Cu含量5个因子。
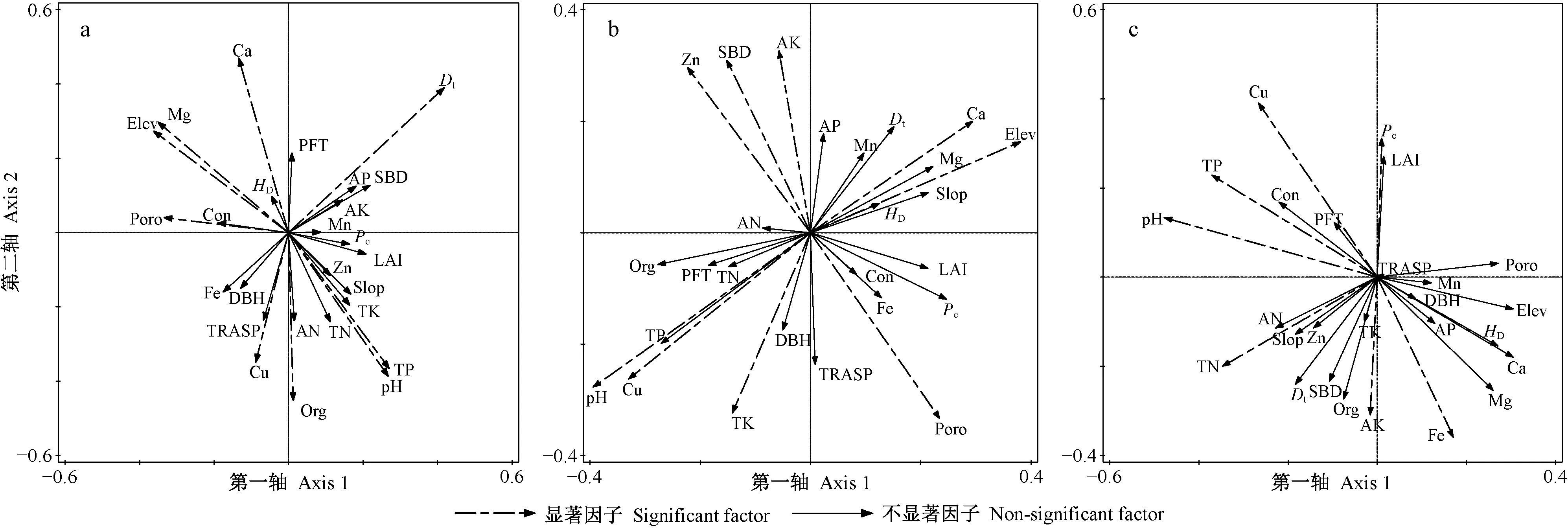
图3 强烈度火干扰后演替前期(a)、中期(b)和后期(c)草本层CCA排序Fig. 3 The CCA sequence diagram of herbaceous layer at early(a), middle(b) and late(c) stage of succession after high-severity fire Elev: 海拔Elevation; Slop: 坡度Slope; TRASP: 坡向Transformation of aspect; PFT: 火后时间Post-fire time; pH: 酸碱度pH value; Con: 电导率Conductivity; SBD: 土壤密度Soil bulk density; Poro: 土壤孔隙率Soil porosity; Org: 有机质Soil organic matter; TN: 全氮Total nitrogen; TK: 全钾Total potassium; TP: 全磷Total phosphorus; AN: 碱解氮Available nitrogen; AK: 速效钾Available potassium; AP: 有效磷Available phosphorus; Fe: 铁含量Fe content; Cu: 铜含量Cu content; Zn: 锌含量Zn content; Mn: 锰含量Mn content; Mg: 镁含量Mg content; Ca: 钙含量Ca content; LAI: 叶面积指数Leaf area index; Pc: 郁闭度Crown density; DBH: 林分平均胸径Average diameter at breast height; Dt: 林分株数密度Density of trees; HD: 林分平均高Average height of stand. 下同The same below.
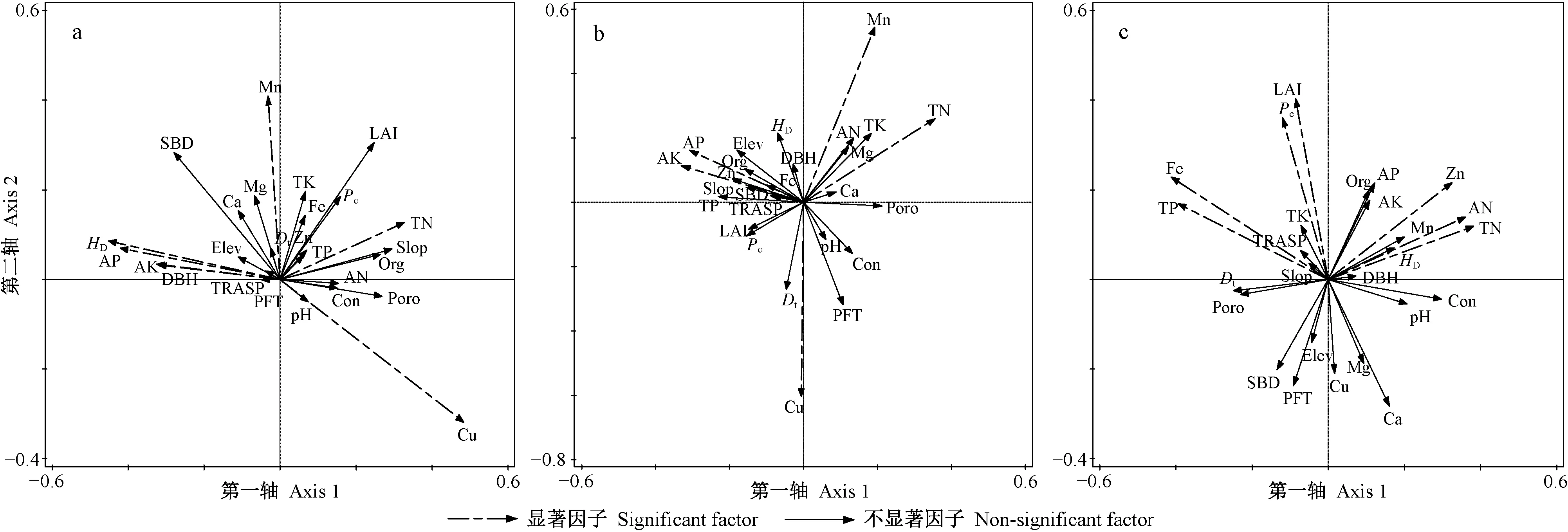
图4 强烈度火干扰后演替前期(a)、中期(b)和后期(c)林冠层CCA排序Fig. 4 The CCA sequence diagram of overstorey at early(a), middle(b) and late(c) stage of succession after high-severity fire
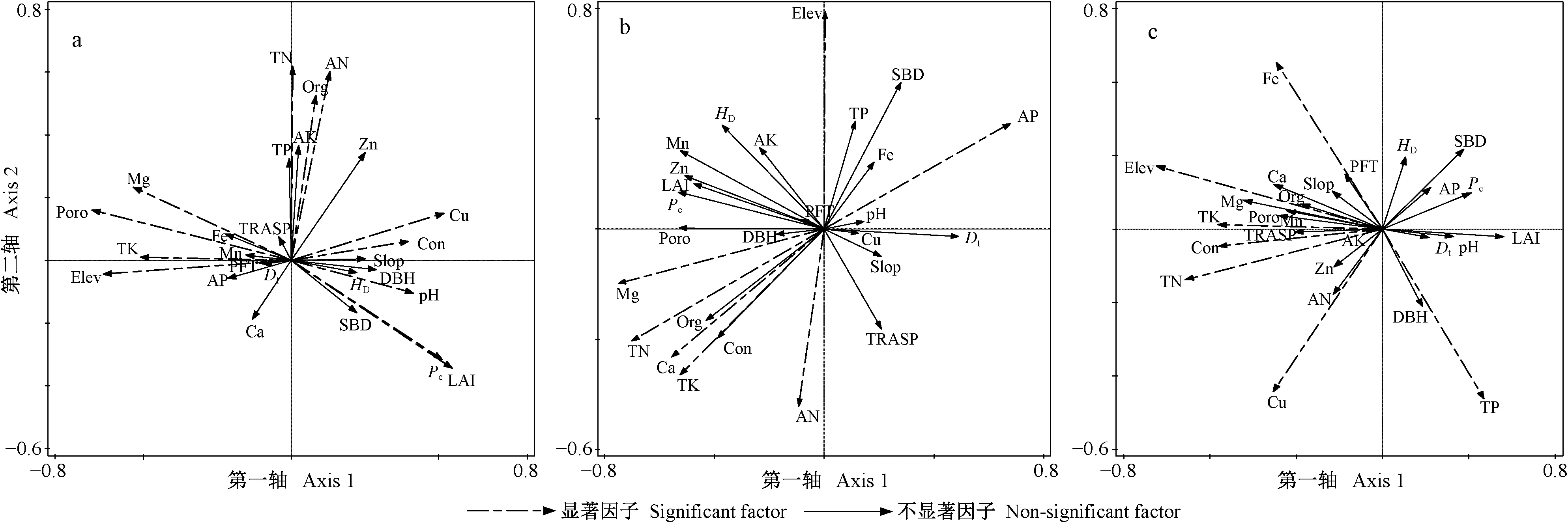
图5 中烈度火干扰后演替前期(a)、中期(b)和后期(c)草本层CCA排序Fig. 5 The CCA sequence diagram of herbaceous layer at early(a), middle(b) and late(c) stage of succession after moderate-severity fire
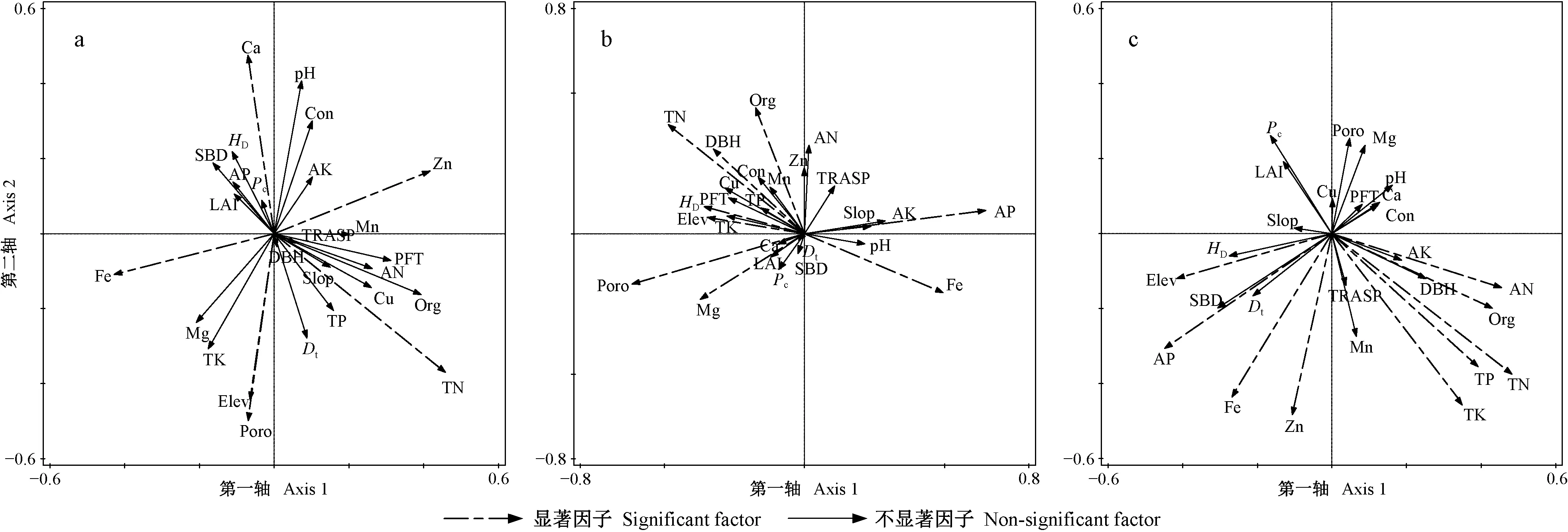
图6 中烈度火干扰后演替前期(a)、中期(b)和后期(c)林冠层CCA排序Fig. 6 The CCA sequence diagram of overstorey at early(a), middle(b) and late(c) stage of succession after moderate-severity fire
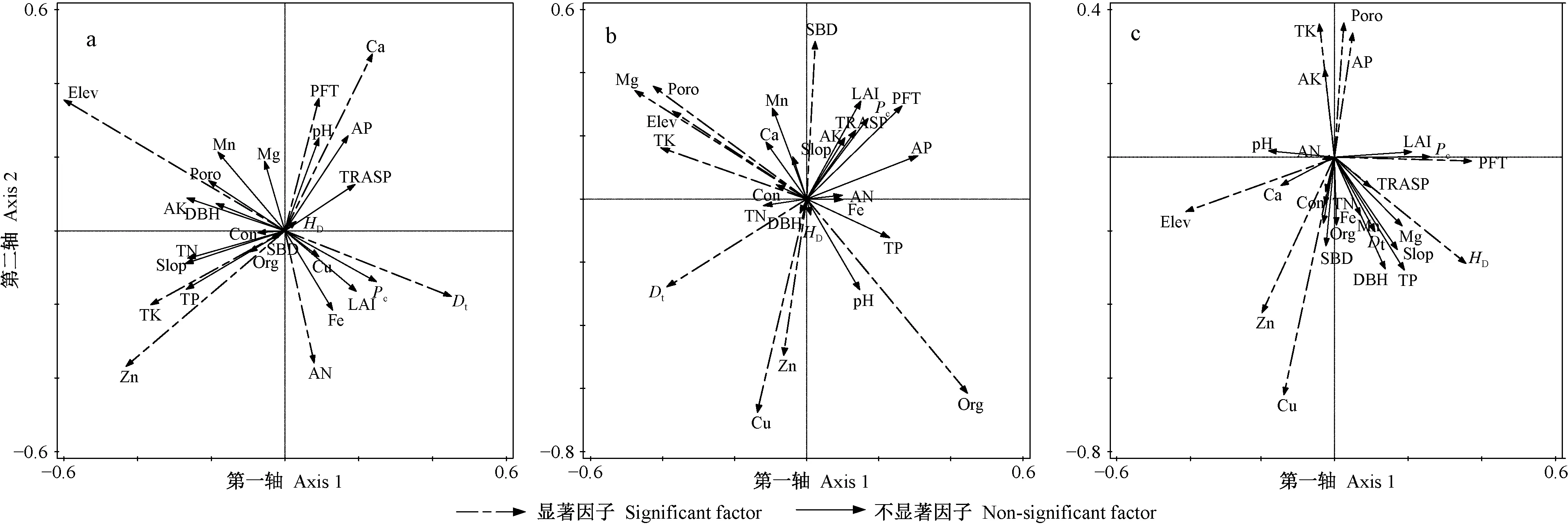
图7 弱烈度火干扰后演替前期(a)、中期(b)和后期(c)草本层CCA排序Fig. 7 The CCA sequence diagram of herbaceous layer at early(a), middle(b) and late(c) stage of succession after low-severity fire

图8 弱烈度火干扰后演替前期(a)、中期(b)和后期(c)林冠层CCA排序Fig. 8 The CCA sequence diagram of overstorey at early(a), middle(b) and late(c) stage of succession after low-severity fire
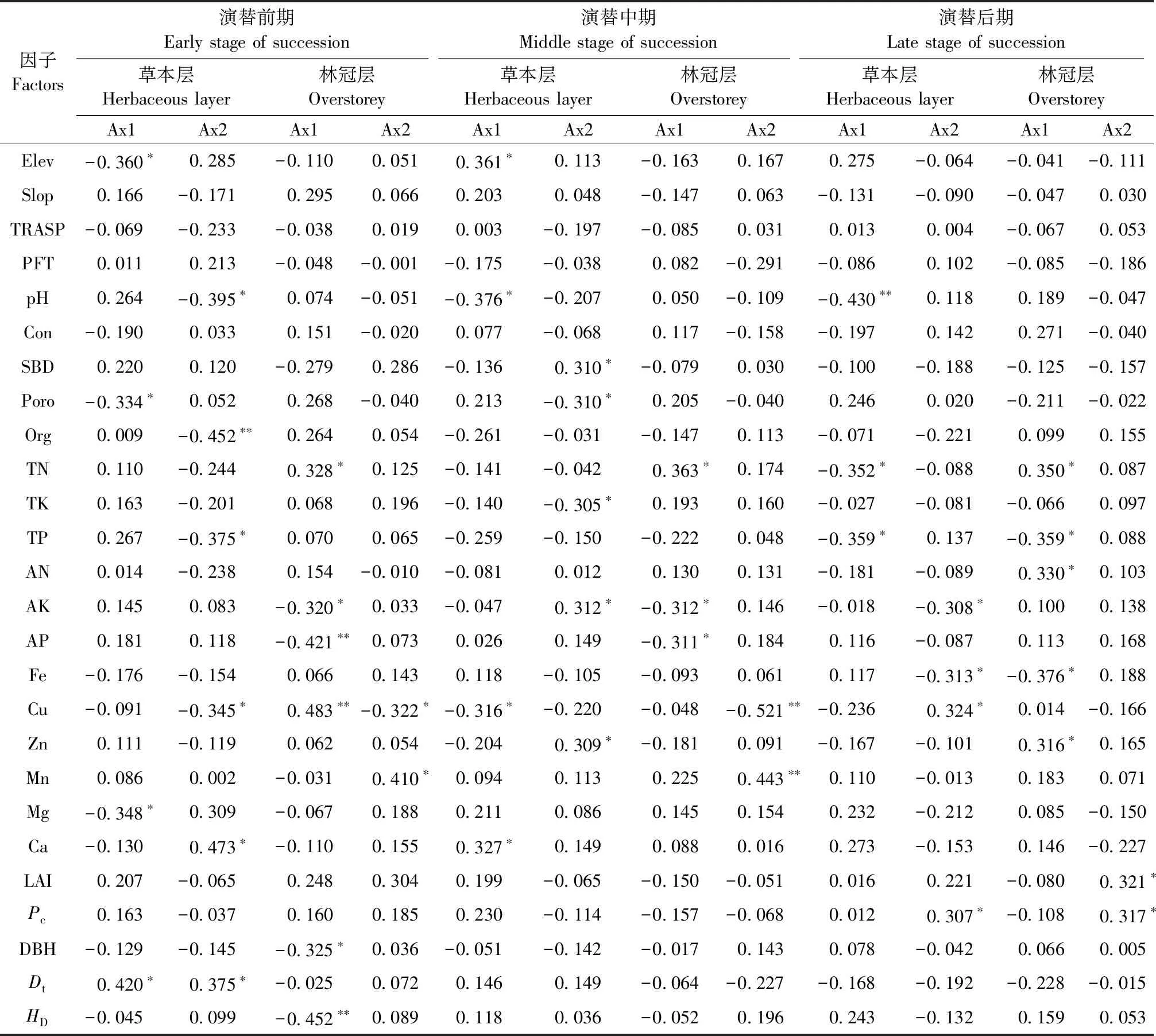
表1 强烈度火干扰后泰加林群落不同演替阶段草本层与林冠层物种CCA排序轴相关系数①Tab.1 Correlation coefficient of CCA ordination axis of herbaceous layer and overstorey species at different succession stages for taiga communities after high-severity fire
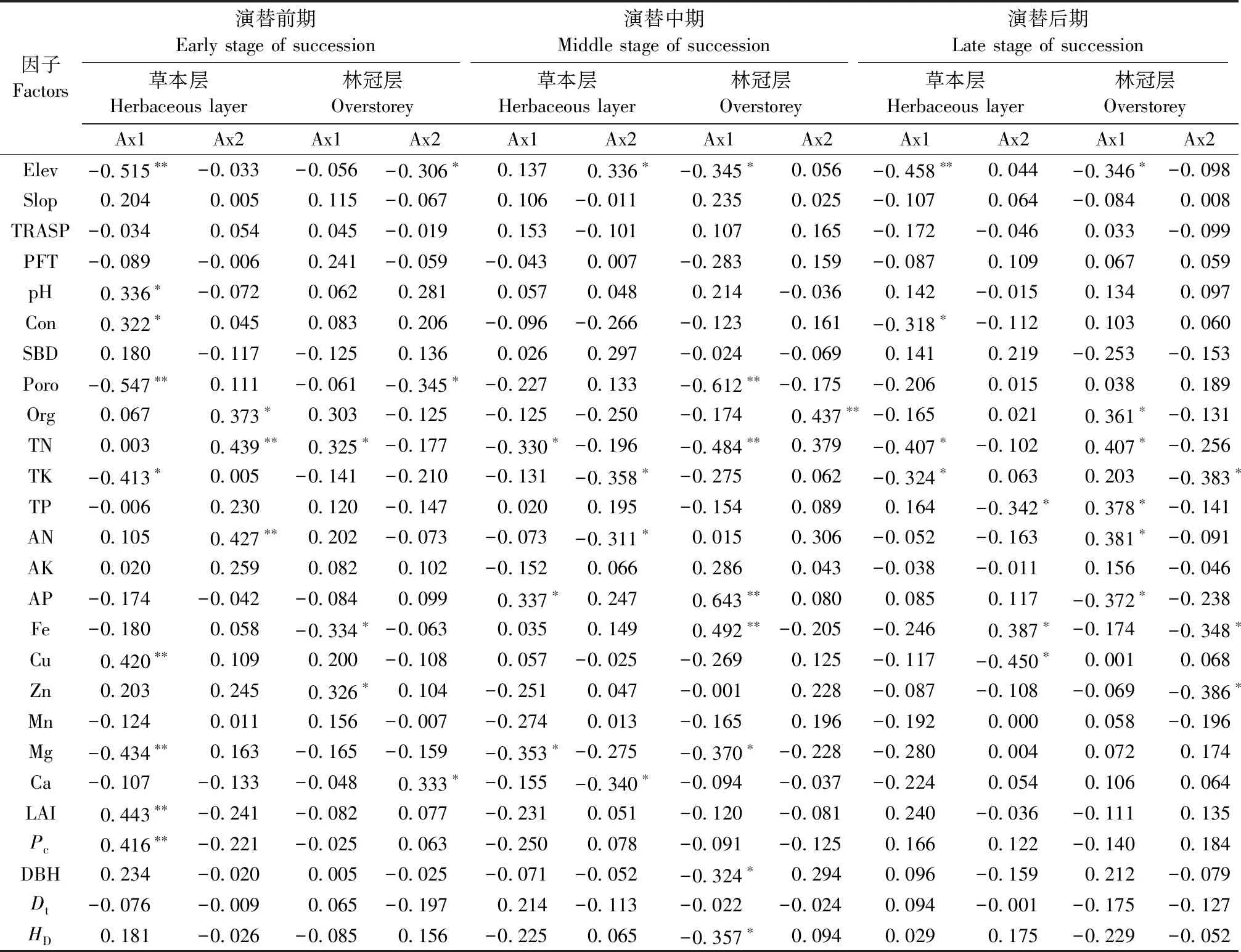
表2 中烈度火干扰后泰加林群落不同演替阶段草本层与林冠层物种CCA排序轴相关系数Tab.2 Correlation coefficient of CCA ordination axis of herbaceous layer and overstorey species at different succession stages for taiga communities after moderate-severity fire
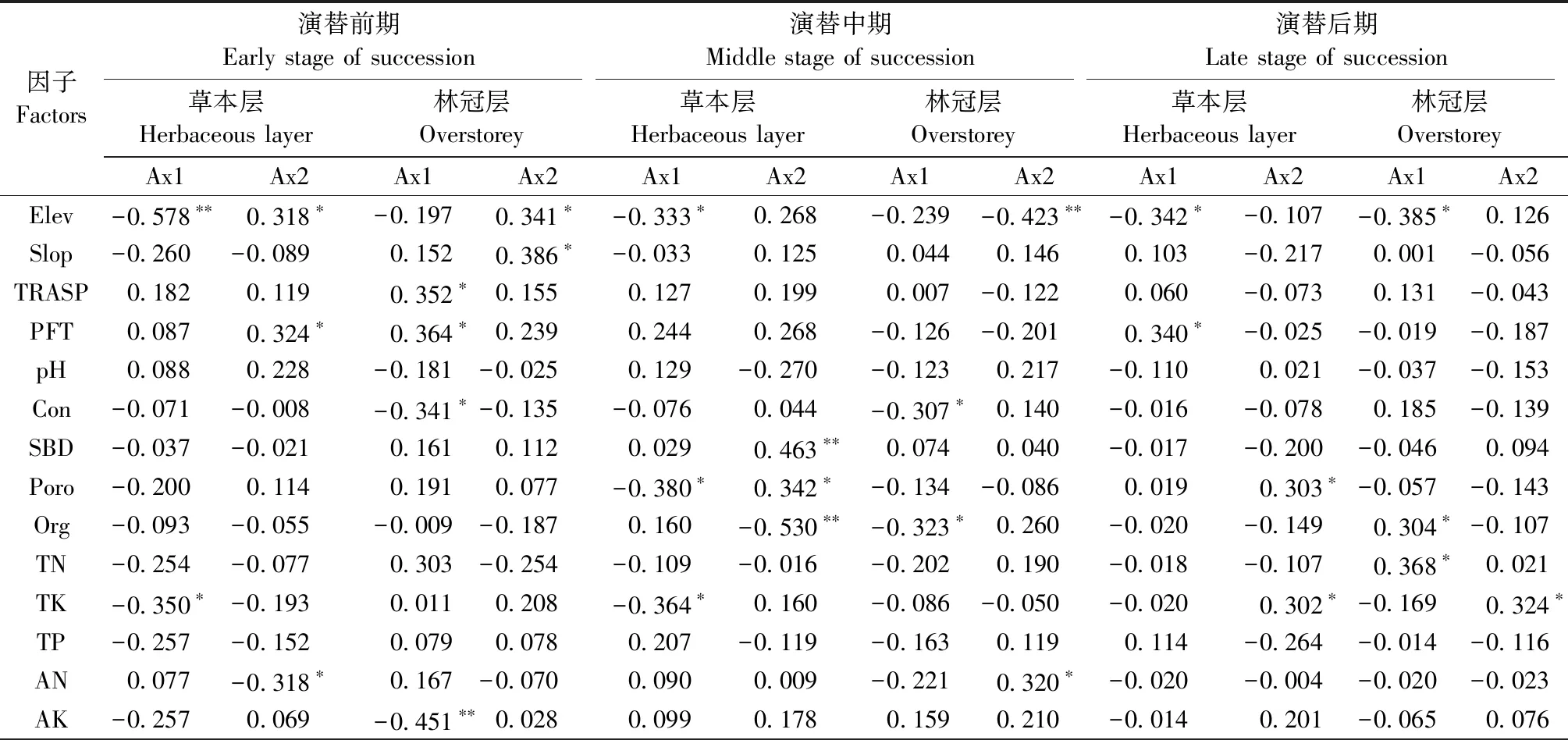
表3 弱烈度火干扰后泰加林群落不同演替阶段草本层与林冠层物种CCA排序轴相关系数Tab.3 Correlation coefficient of CCA ordination axis of herbaceous layer and overstorey species at different succession stages for taiga communities after low-severity fire
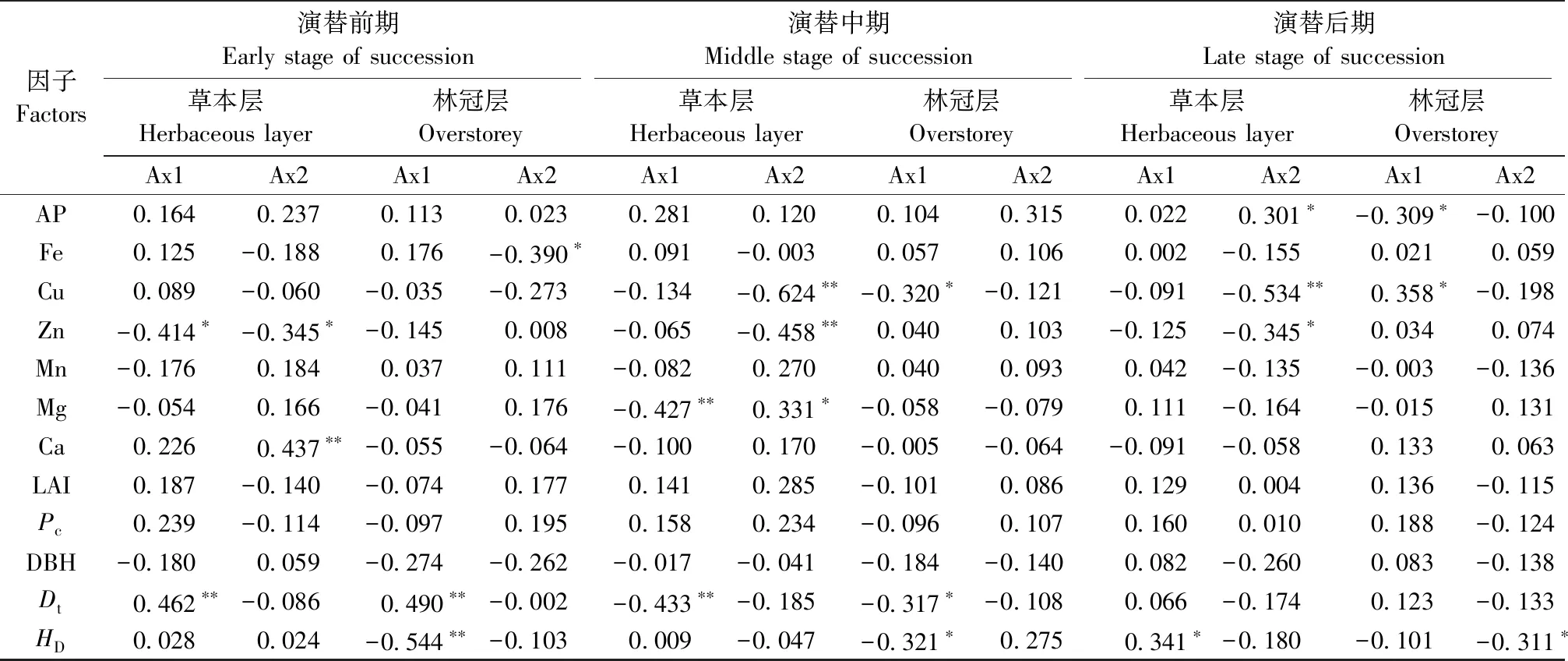
续表3 Continued
4 讨论
4.1 草本层与林冠层物种连锁关系
关于森林群落的草本层与林冠层物种连锁关系,现有研究结果存在争议。有学者认为草本层和林冠层物种很大程度上独立响应环境梯度,可能是因在景观上连续存在的环境梯度在局部尺度上为不连续的,并且草本层和林冠层物种可能以不同方式和速度响应环境梯度(McCuneetal., 1981; Sagersetal., 1997; Lyonetal., 1998); 而另一些学者则用实例说明在森林生态系统中确实存在草本层与林冠层的物种连锁关系,但它发生在比景观尺度小的空间尺度上(Gilliametal., 2014),并且认为物种连锁关系是由多因素引起的,特别是林冠层引起的草本层光照和土壤pH值等条件变化(Nematietal., 1995),除此之外还包括土壤因子和干扰机制(Robertsetal., 1988)。
本文结合地形、土壤、火干扰和林分因子,分析了不同烈度火干扰后泰加林群落在不同演替阶段的草本层与林冠层的物种连锁关系,表明存在连锁关系且其显著性水平随演替阶段进行逐渐升高。这表明火干扰后的连锁关系随森林演替向前发展变得越来越紧密,这与Gilliam等(1995)的研究结果类似。有研究发现,森林群落草本层与林冠层物种多样性之间存在正相关(Ingerpuuetal., 2003; Mölderetal., 2008),但也有研究表明不存在相互影响(Ewaldetal., 2002; Borchseniusetal., 2004; Houle, 2007)。本研究发现在强烈度和中烈度火干扰后,草本层与林冠层物种的连锁关系在演替前期呈显著负相关(P<0.05),在演替中期和后期呈极显著正相关(P<0.01); 而在弱烈度火干扰后,二者连锁关系在演替前、中和后期均呈极显著正相关(P<0.01)。究其原因,相对于弱烈度火干扰,中烈度和强烈度火干扰更大程度地降低了林冠层郁闭度,群落中的光照强度增强,从而增加了林内环境的异质性(孙家宝, 2010)。群落内阳性树种[垂枝桦、五蕊柳(Salixpentandra)]和喜光草本植物得以萌发生长,但阳生树种幼苗的平均高度大于草本层,会先接受到光照和土壤等条件的影响而迅速繁殖生长,从而最先占据生长空间,抑制草本层发育(邢玮等, 2006; 刘景, 2017)。总而言之,中烈度和强烈度林火干扰后,林冠层树种幼苗与草本层植物处于竞争状态,因而林冠层与草本层物种的连锁关系在演替前期为负相关。随着群落演替向前发展,林冠郁闭度增加,耐阴植物种增多,草本层物种的生存一定程度上依赖于林冠层树种创造的微生境(余敏, 2013; McIntoshetal., 2016; Burrascanoetal., 2018),因而表现为正相关。此外,草本层与林冠层物种连锁关系随火烈度增强而减弱,这可能是因不同烈度火干扰对群落物种组成和空间结构的影响有差异,一般火烈度越强,群落恢复所需时间越长(苗庆林等, 2015),因而表现出草本层与林冠层的物种连锁关系随火烈度增强而减弱。
4.2 草本层与林冠层物种连锁关系的成因
草本层与林冠层物种的连锁关系是由于它们对相同环境梯度的响应相似性而产生的,且这种响应会在演替过程中变化(Gilliametal., 1995),本文结果与之类似。强烈度火干扰后,在泰加林群落演替前期,草本层和林冠层具有一致性响应且达显著水平的环境因子仅有土壤Cu含量,演替中期为土壤AK和Cu含量2个因子,而演替后期为Pc和土壤TN、TP、Fe含量4个因子。中烈度火干扰后,在演替前期具有一致性响应且达显著水平的环境因子为Elev、土壤Poro和土壤TN含量3个因子,演替中期为Elev和土壤TN、AP和Mg含量4个因子,演替后期则为Elev和土壤TN、TK、TP和Fe含量5个因子。弱烈度火干扰后,在演替前期具有一致性且达显著性水平的因子为Elev、PFT和Dt共3个因子,演替中期为Elev、Dt和土壤Org、Cu含量4个因子,演替后期则为Elev、HD和土壤TK、AP、Cu含量5个因子。由此可知,不同烈度火干扰后,随群落演替向前发展,草本层与林冠层物种具一致性响应且达显著性水平的因子数在增加,这可能是造成随火后时间推移连锁关系紧密程度在增加的原因之一。此外有研究表明,演替前期草本层物种的生长发育主要受土壤养分影响,而林冠层树种主要受海拔和林分密度影响(Wilsonetal., 1993); 在演替中期和后期,草本层物种生长发育会受林冠层树木密度、郁闭度、平均高等林分因子影响,其物种组成和分布格局对林分结构的响应增强,而此2个阶段的林冠层树种的生长发育也受林分因子影响(Gilliametal., 1993),因而连锁关系更紧密,本文结果与之类似。本研究还表明,海拔对草本层物种和林冠层树种的生长发育有重要影响,这是由于海拔包含了温度、湿度、光照、土壤等多种环境因子变化形成的空间生态梯度,影响着林冠层和草本层物种的分布和生长(Liebermanetal., 1996; 唐志尧等, 2004)。土壤因子中除植物生长发育所需N、P、K等营养元素外,Cu、Mg、Fe含量影响也较大,它们均与叶绿素形成有关,进而影响着植物光合作用。此外,Cu还是植物体内多种氧化酶的组分,参与植物呼吸作用并影响植物对Fe的利用,Mg还是许多酶的活化剂,与碳水化合物的代谢、磷酸化作用、脱羧作用密切相关,Fe还是一些酶的组分,参与细胞呼吸作用,且老叶中的Fe不能被新叶重复利用,因而对泰加林物种的生长发育有重要影响(王忠, 2009)。林分因子中的Pc对林内光照环境有重要影响,Dt和HD对林下生长空间形成制约,影响着林冠层树种幼苗和草本层物种的更新与生长,在群落演替过程中有重要作用(曾晓阳等, 2017)。
本研究验证了泰加林群落火成演替过程中在草本层与林冠层物种之间存在连锁关系,将有助于加深对泰加林火干扰后生态系统演替过程中物种关系的理解,可为进行与泰加林植物多样性要求一致的生物多样性保护和可持续经营提供理论依据。但本研究是基于群落尺度的研究,要全面理解泰加林火成演替过程中的草本层与林冠层的物种连锁关系,还需进一步从不同尺度研究。
5 结论
火干扰后,喀纳斯泰加林群落演替过程中的草本层与林冠层物种之间存在连锁关系,且随火烈度从强到弱,连锁关系由弱变强; 另外,随着演替向前发展,连锁关系的紧密程度增加。产生连锁关系的原因可能是草本层和林冠层物种多样性格局对相同环境梯度的响应具有一致性。
