甘蔗姊妹系对低钾胁迫响应的差异分析
2021-05-19曾巧英吴嘉云冯晓敏张南南吴自林齐永文
曾巧英 吴嘉云 冯晓敏 张南南 吴自林 齐永文


摘 要:以来源于同一甘蔗杂交组合的姊妹系‘粤糖55号和‘粤糖00-236为材料,通过对长期低钾胁迫下甘蔗生长、钾含量、钾利用效率和钾吸收效率以及短期低钾胁迫下钾含量、抗氧化酶活性变化的研究,探讨甘蔗姊妹系耐低钾能力的差异。研究结果显示:在长期低钾胁迫下,2个品种的株高、茎径和生物量存在显著差异,‘粤糖55号表现出显著的下降,而‘粤糖00-236仅根系生物量显著下降。2个品种的钾含量显著下降,‘粤糖00-236下降幅度高于‘粤糖55号,但是低钾胁迫下‘粤糖00-236的钾利用效率和钾吸收效率均高于‘粤糖55号。短时间低钾胁迫下,‘粤糖55号根系钾含量在胁迫48 h后开始下降,而‘粤糖00-236根系的钾含量在胁迫的24 h已经显著低于对照。‘粤糖55号根系的MDA含量在胁迫48 h后显著上升,而‘粤糖00-235則在胁迫的72 h显著上升。‘粤糖55号的SOD和POD活性在低钾胁迫下表现为下降,而‘粤糖00-236则在48 h后出现上升。由此可见,同一组合的甘蔗姊妹系在耐低钾胁迫能力上存在显著差异,提高钾的吸收、利用效率和根系抗氧化酶活性有助于提高甘蔗的耐低钾胁迫能力。
关键词:甘蔗;低钾胁迫;钾含量;抗氧化酶活性
中图分类号:S566.1 文献标识码:A
Different Responses to Low Potassium Stress Between Two Sugarcane Varieties from the Same Hybrid Combination
ZENG Qiaoying, WU Jiayun, FENG Xiaoming, ZHANG Nannan, WU Zilin, QI Yongwen*
Guangdong Key Lab of Sugarcane Improvement and Biorefinery / Guangzhou Sugarcane Industry Research Institute, Guangzhou, Guangdong 510316, China
Abstract: Two sugarcane varieties (YT55 and YT00-236) from the same hybrid combination were used to analyze the different responses to low potassium (K) stress. The growth parameters, K contents, K utilization and uptake effeciency under longterm low K stress, and the K contents, activites of antioxidative enzymes under shortterm low K stress were investigated. The results revealed that significant differences were found for height, diameter, biomass between the two varieties under longterm low K stress. The height, diameter, shoot and root biomass decreased significantly in YT55 under longterm low K stress, while only root biomass showed noticeable reduction in YT00-236. The K contents in shoots and roots of the two varieties decreased under longterm low K stress. The rate decrease of K content in YT55 was higher than that in YT00-236 under longterm low K stress, but the utilization effeciency and uptake effeciency in YT00-235 were higher than those in YT55. The K content in the roots of YT55 began to decrease after 48 h under low K stress, whereas significant reduction was found for K content in the roots of YT00-236 after 24 h under low K stress. The MDA content in the roots of YT55 showed significant increase after 48 h under low K stress, but after 72 h under low K stress in the roots of YT00-236. The activities of SOD and POD decreased under low K stress in roots of YT55, but notably increased after 48 h under low K stress in the roots of YT00-236. These results indicated that significant difference for low K tolerance existed between the varieties from the same hybrid combination. Improvement in K utilization and uptake effeciency and activities of antioxidative enzymes might help to increase the low K tolerance in sugarcane.
Keywords: sugarcane; low K stress; K content; activities of antioxidative enzymes
DOI: 10.3969/j.issn.1000-2561.2021.03.019
甘蔗是我国重要的糖能兼用作物,主产于广东、广西、云南和海南4省(区)。这4个省份的土壤为典型的红壤,受土壤的形成和气候条件影响,甘蔗主产区的土壤中有效钾往往缺乏[1]。由于甘蔗的生物量高,在甘蔗生长期内需要吸收大量的钾元素以满足其生长的需要,据估计,每生产1 t甘蔗大约需要吸收1~2.5 kg的K2O[2]。钾素作为植物重要的必需元素,在植物生长发育中起重要作用,它能影响植物的膜电位、离子平衡、酶活性,蛋白和糖的转运等生理过程[3]。研究显示,植物在缺钾条件下,植株钾含量的下降[4]、光合系统受抑制[5]、植株体内的活性氧(ROS)平衡受到破坏形成氧化胁迫等[6],最终导致植株生长受到抑制,表现出生物量下降、根系生长受到影响[7-8]。在对甘蔗的研究中同样发现,在缺钾条件下,甘蔗生长、光合作用受到抑制,导致生物量下降,蔗茎的产量下降[9-11]。因此,在甘蔗生长期内需要大量施用钾肥,而大量钾肥的施用既增加了成本,同时也增加环境风险。培育和筛选耐低钾甘蔗品种是最有效解决这些问题的方法。
由于甘蔗是典型的非整倍多倍体,基因组极其复杂[12],杂交后代分离严重,同一杂交组合的后代从形态、产量、糖分等性状上均存在不同的分离类型[13]。在低钾胁迫下,不同甘蔗品种间生物量、钾含量、钾吸收和利用效率,光合参数、抗氧化酶活性等指标表现出显著的差异[10]。但是目前还没有关于甘蔗姊妹系对低钾胁迫响应差异的报道。本研究选用了2个甘蔗来自于同一甘蔗杂交组合‘粤农73-204/CP72-1210的姊妹系品种‘粤糖55号和‘粤糖00-236。2个品种在产量、糖分、植株外形等方面均存在显著的差异。通过分析长期和短期低钾胁迫对甘蔗生长、钾含量、钾吸收和利用效率、抗氧化酶等指标的差异,探讨来源于同一组合品种的耐低钾胁迫能力的差异情况,为选育耐低钾甘蔗品种提供依据。
1 材料与方法
1.1 材料
以2个来源于同一甘蔗杂交组合‘粤农73-204/CP72-1210的2个姊妹品种‘粤糖55号和‘粤糖00-236为试验材料,试验材料采自广东省科学院生物工程研究所资源保育圃。
1.2 方法
1.2.1 试验处理 试验均在广东省科学院生物工程研究所温室大棚中进行。试验分为以下2个部分:(1)长期低钾胁迫对甘蔗姊妹系的影响。选取均匀一致的种茎,砍成单芽,蔗种用5%的多菌灵消毒,然后整齐排布于装有石英砂的方盆中进行发芽,保持石英砂湿润。待幼苗长出4片叶片以后,选取生长一致的幼苗移到装有10 L营养液中进行预培养2周。期间在幼苗上的根系长出,将种茎剪除。设置2个处理:正常钾(CK,钾浓度3 mmol/L)和低钾处理(LK,钾浓度100 ?mol/L),设5次重复。营养液采用改进的1/2浓度Hoagland营养液[9]。每隔2 h通气1 h,每一周换一次营养液,培养时间为60 d。测定株高、茎径、地上部分干重、根系干重、地上部分和根系的钾含量。
(2)短时间低钾胁迫对甘蔗钾含量和根系抗氧化酶活性影响。甘蔗种茎发芽处理和幼苗的预培养与试验一相同。设置2个处理:正常钾(CK,钾浓度3 mmol/L)和低钾处理(LK,钾浓度100 ?mol/L),5次重复,营养液采用改进的1/2浓度Hoagland营养液[9],每隔2 h通气1 h。在处理的24、48、72 h和15 d取样测定钾含量、丙二醛含量、SOD、POD和CAT酶活性。
1.2.2 指标测定 钾含量测定:植株样品置于烘箱中105 ℃杀青,70 ℃烘干至恒重。用植物粉碎机将植株样品粉碎过20目筛。称取0.2 g样品,在消煮炉上H2SO4-H2O2湿灰化法进行消煮,消煮后定容至250 mL。钾含量采用Model 425火焰光度计(Sherwood, UK)进行测定。计算各处理甘蔗植株的养分含量。钾吸收和利用效率根据Damon等[14]的方法进行计算:
植株钾利用效率=全株干重(g)/植株地上部钾积累量(g);
植株钾吸收效率=整株钾积累量(g)/根干重(g)。
抗氧化酶活性的测定:植株样品取样后,用液氮速冻,置于?80 ℃进行保存,抗氧化酶SOD、POD和CAT酶活性的测定参照Li等[15]的方法進行。抗氧化酶提取液为50 mmol/L Tris-HCl缓冲液(pH 7.0,内含1 mmol/L EDTA,1 mmol/L ASA,1 mmol/L DTT,1 mmol/L GSH,5 mmol/L MgCl2和20%甘油),植株样品采用液氮研磨,称取样品约0.5 g,加入5 mL的提取液,在4 ℃下12 000 r/min离心10 min,取上清液并分装,液氮冷冻后保存在?80 ℃,待测定时取出。SOD,CAT和POD活性测定根据Li等[15]的方法采用分光光度计进行测定,其中测定SOD和CAT酶活加入50 ?L酶提取液,测定POD酶活性时加入10 ?L酶提取液。
丙二醛(MDA)含量测定:植株样品用液氮粉碎后,取0.5 g加5 mL 10%三氯乙酸(TCA)进行提取,然后在4 ℃下12000 r/min离心10 min,取上清液。吸取离心的上清液2 mL(对照加2 mL蒸馏水),加入2 mL 0.6%硫代巴比妥酸(TBA,用10%的三氯乙酸配制)溶液,混匀物于沸水浴室反应15 min,迅速冷却后再离心。取上清液用紫外可见分光光度计测定532、600和450 nm波长下的消光度。用下列公式计算提取液中MDA浓度,再根据样品质量计算出MDA含量。MDA
含量的单位为nmol/g(以鲜重计)。
C(μmol/L)=6.45(OD532?OD600)?0.56OD450
叶绿素含量的测定:新鲜植物叶片用液氮粉碎,称取样品约0.1 g,加入95%的乙醇10 mL,置于黑暗条件下。浸泡至植物组织发白,无绿色为止,测定提取液在波长665和649 nm下测定吸光度。葉绿素a和叶绿素b的浓度(Ca、Cb,mg/L)按以下公式进行计算,再根据样品重量计算叶片叶绿素含量。
Ca=13.95A665?6.88A649;
Cb=24.96A649?7.32A665。
1.3 数据分析
所有试验数据的方差分析均采用DPS 7.05软件进行,以LSD法进行多重比较(P<0.05)。
2 结果分析
2.1 长期低钾胁迫对甘蔗姊妹系生长影响的差异
由图1可知,2个甘蔗品种对低钾胁迫的响应存在显著差异。在低钾胁迫60 d后,‘粤糖55 号的株高,茎径、地上部分干重、根系干重均显著低于正常钾处理(P<0.05),与正常钾处理相比分别下降了17.44%、12.70%、35.472%和36.57%。而‘粤糖00-236仅根系干重显著下降了21.85%,株高、茎径和地上部分干重虽有所下降,但与对照相比无显著差异。由此可见,2个甘蔗姊妹系长期低钾胁迫的耐性存在显著的差异,其中‘粤糖00-236对低钾胁迫具有较高的耐性。
2.2 长期和短期低钾胁迫对甘蔗钾含量的影响
由图2可知,长期低钾胁迫下,2个品种的地上部分钾含量和根系的钾含量均显著下降(P<0.05),其中‘粤糖00-236下降幅度高于‘粤糖55号。在低钾胁迫下,‘粤糖55号的地上部分和根系的钾含量分别下降了67.59%和71.98%,而‘粤糖00-236的地上部分和根系钾含量下降了74.75%和81.62%。
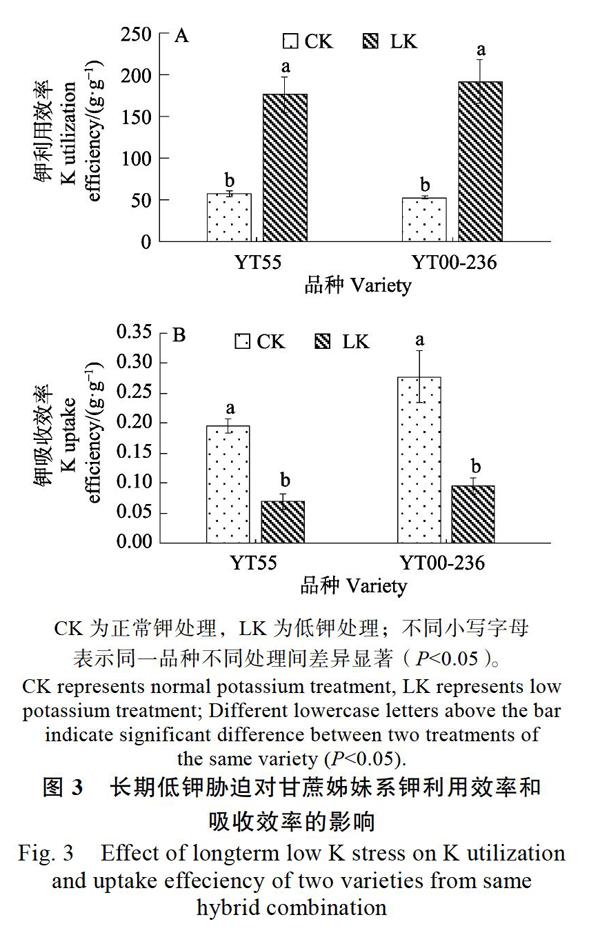
由图3可知,对2个品种的钾吸收和利用效率分析显示,低钾胁迫下2个品种的钾利用效率显著提高(P<0.05)。其中‘粤糖55号的钾利用效率是对照的3.1倍,‘粤糖00-236的钾利用效率是对照的3.65倍。‘粤糖00-236低钾胁迫下的钾利用效率比‘粤糖55号高8.7%。正常钾条件下,‘粤糖00-236的钾吸收效率显著高于‘粤糖55号。低钾胁迫下,2个品种的钾吸收效率显著下降,分别下降了64.28%和65.93%。虽然‘粤糖00-236钾吸收效率在低钾胁迫下下降的幅度高于‘粤糖55号,但是低钾条件下,其绝对值仍然高35.70%。
对15 d内低钾胁迫2个姊妹系地上部分和根系的钾含量变化进行分析(图4),结果显示,二者在短时间内的钾含量变化也存在显著的差异。‘粤糖55号的地上部分钾含量在胁迫的72 h开始下降,15 d后显著低于对照,根系则在胁迫的48 h开始下降,但是在48 h和72 h与对照无显著差异,15 d后显著下降。‘粤糖00-236的地上部分和根系钾含量在低钾胁迫处理24 h开始就已经显著低于对照,到处理的15 d钾含量大幅度下降。
2.3 短期低钾胁迫对甘蔗姊妹系抗氧化系统的影响
由表1可知,在低钾胁迫24 h后,甘蔗姊妹系根系的丙二醛含量上升,其中‘粤糖55号上升了20.43%,‘粤糖00-236上升了10.67%,但是与对照无显著差异。到胁迫48 h,‘粤糖55号的MDA含量显著上升,‘粤糖00-236仍然与对照无显著差异。胁迫72 h后,‘粤糖00-236的根系MDA含量才与对照存在显著差异。胁迫15 d后,根系的MDA含量与对照均不存在显著差异。
由表2可知,低钾胁迫下,甘蔗姊妹系的SOD酶活性发生了变化,‘粤糖55号根系SOD酶活性在胁迫的各时间点均低于对照,但与对照不存在显著差异。‘粤糖00-236的SOD酶活性在胁迫的24 h表现出活性的下降,但与对照无显著差异。而到了胁迫的48 h,SOD酶活性表现出明显上升,比对照高21.22%,胁迫的72 h和15 d SOD酶活性也均高于对照,但与对照不存在显著差异。
由表3可知,低钾胁迫下的POD酶活性变化与SOD酶活性变化类似,‘粤糖55号的根系POD酶活性在胁迫的24、48、72 h,POD酶活性均低于对照,但是与对照不存在显著差异。而‘粤糖00-236的POD酶活性在胁迫的24 h,根系POD酶活性低于对照,但与对照无显著差异;到胁迫48 h,POD酶活性比对照高9.34%,胁迫的72 h显著高于对照。
由表4可知,2个品种的根系CAT酶活性在低钾胁迫下均低于对照。2个品种的CAT酶活性均在胁迫的48 h显著低于对照,其中‘粤糖55号CAT酶活性下降了24.58%,而‘粤糖00-236下降了19.00%。其他时间点,2个品种低钾胁迫下的CAT酶活性均与对照无显著差异。
3 讨论
由于钾是植物必需的元素之一,低钾胁迫严重影响植物生长,往往表现出生物量的下降[8]。不同作物和同一作物的不同品种间对低钾胁迫的耐性存在显著差异,耐低钾品种的生长受低钾胁迫影响较敏感品种小,生物量等性状下降幅度小[16]。在对不同甘蔗品种对低钾胁迫响应的研究显示,低钾胁迫下耐低钾品种的株高、茎径、生物量等性状下降幅度低于敏感品种[17]。本研究中,来源于同一组合的2个品种对低钾胁迫耐性各不相同,‘粤糖00-236表现出较高的耐低钾胁迫能力,在低钾胁迫下除了根系生物量显著下降外,其他性状与对照无显著的差异,而‘粤糖55号各性状均表现出显著下降。由此可见,来源于同一甘蔗杂交组合的后代对低钾胁迫的响应是存在显著差异的,这为利用优异的甘蔗杂交组合筛选耐低钾品种提供了依据。
在低钾胁迫下,植株的钾含量出现显著的下降,植物通常情况下通过提高植物对钾的吸收和利用效率来减轻低钾胁迫对植物生长的影响,这是植物应对低钾胁迫的耐性机制之一[16, 18]。但是,不同作物在应对低钾胁迫策略上存在显著的差异。在对大豆的报道中显示耐性品种在低钾胁迫下,植株钾含量下降幅度要小于钾敏感品种,植株通过增加钾的吸收来提高植株对钾的耐性[19]。然而在玉米的研究中发现,耐性品种钾含量下降幅度高于敏感品种,玉米通过提高钾利用效率来提高耐低钾胁迫能力[20]。在对甘蔗姊妹系的研究中,2个品种的钾含量对低钾胁迫的响应存在差异。在长期低钾胁迫下2个品种的钾含量都出现了不同程度的下降,‘粤糖00-236的钾含量下降的幅度显著高于‘粤糖55号,并且在短时间的低钾胁迫下,‘粤糖00-236在胁迫24 h开始,根系和叶片钾含量下降早于‘粤糖55号。但是,对比2个品种钾吸收效率时可见,长期低钾胁迫处理中,无论是在正常钾水平还是低钾胁迫下,‘粤糖00-236钾吸收效率均高于‘粤糖55号,而且在短时间钾胁迫下,‘粤糖00-236地上部分的钾含量也始终高于‘粤糖55号。另外,尽管在正常钾条件下,2个品种的钾利用效率相当;在低钾胁迫下,2个品种的钾利用效率提高。但是,‘粤糖00-236钾利用效率提高幅度高于粤‘糖55号。因此,‘粤糖00-236相对于‘粤糖55号更耐低钾胁迫的机制之一是提高植株对钾的吸收和利用效率。
根系植植物主要吸收水分、养分等器官,胁迫对根系的伤害直接影响到植物对养分的吸收。在低钾条件下,植物体内代谢活动受到影响,活性氧(ROS)的平衡受到破坏,植物体内ROS含量增加,形成氧化胁迫。ROS引起植物细胞膜脂的过氧化,从而影响到细胞膜的稳定性,影响植物养分的吸收和运输[21]。作为植物氧化胁迫的指标之一丙二醛是膜脂过氧化的产物,在低钾脅迫下,丙二醛含量显著上升[6, 22]。植物的抗氧化酶在清除ROS,保持植物体内的ROS的平衡起重要的作用。SOD酶是植物清除ROS的第一道防线,可以将活性较高的O2-歧化为活性较低的H2O2[23],随后H2O2被POD、CAT、APX等酶降解为水和氧气,从而消除ROS对细胞的影响[24],因此提高植物体内的抗氧化酶活性是植物对抗逆境胁迫诱导产生氧化胁迫的重要机制之一[6]。在斑茅研究中发现,低钾胁迫下叶片和根系的SOD、POD和CAT酶活性均高于高钾处理[25],提高抗氧化酶活性可以提高斑茅耐低钾胁迫的能力。本研究中对短时间低钾胁迫下的抗氧化系统分析中发现,2个品系的MDA含量出现不同程度的上升,但是2个品种间存在显著的差异。粤糖55号的MDA在胁迫的48 h出现显著的上升,而‘粤糖00-226则是在胁迫的72 h出现显著的上升,说明‘粤糖00-236受到的氧化胁迫的时晚要于‘粤糖55号。这个结果也在抗氧化酶活性分析得到验证,‘粤糖00-236的SOD和POD酶活性在24 h下降,但是随后在48和72 h出现显著上升,而‘粤糖55号的SOD和POD酶活性在72 h内均表现为下降。‘粤糖00-236根系中SOD和POD酶活性上升有助于清除根系过多的活性氧,减轻根系的氧化胁迫,保持根系的膜结构的稳定性,进而保持根系对钾的吸收,提高其在低钾胁迫下的钾吸收效率和耐低钾胁迫的能力。
综上所述,同一组合2个姊妹系间耐低钾能力存在显著的差异。低钾胁迫下,敏感型的品系生长受到抑制,表现出生物量、株高和茎径的下降,而耐性品系受影响较小。长期低钾胁迫下甘蔗的钾含量下降,但是耐性品种的钾吸收和利用效率高于敏感品种。短期低钾胁迫下,耐性品种提高根系的SOD和POD酶活性提高,降低根系氧化胁迫,提高植株耐低钾能力。
参考文献
[1] 江 永, 黄忠兴. 我国蔗区土壤主要养分的分析研究[J]. 甘蔗糖业, 2001, 1(5): 5-10.
[2] Gopalasundaram P, Bhaskaran A, Rakkiyappan P. Inte-grated nutrient management in sugarcane[J]. Sugar Tech, 2012, 14(1): 3-20.
[3] Wang M, Zheng Q, Shen Q, et al. The critical role of potassium in plant stress response[J]. Int J Mol Sci, 2013, 14(4): 7370-7390.
[4] 郑 超, 李奇伟, 黄振瑞, 等. 不同品种甘蔗对钾素吸收差异性的研究[J]. 热带作物学报, 2011, 32(12): 2221-2225.
[5] 杜 琪, 王 宁, 赵新华, 等. 低钾胁迫对玉米苗期光合特性和光系统Ⅱ性能的影响[J]. 核农学报, 2019, 33(3): 592-599.
[6] 况 帅, 冯 迪, 宋 科, 等. 低钾胁迫对烟草幼苗活性氧及抗氧化酶系统的影响[J]. 中国烟草学报, 2018, 24(02): 48-54.
[7] 刘 宁, 于海秋, 王晓磊, 等. 不同耐性玉米自交系苗期根系对低钾胁迫的生物学响应[J]. 玉米科学, 2012, 20(6): 85-88, 93.
[8] 郭 泽, 李子绅, 代晓燕, 等. 低钾胁迫下外源生长素对烟草根系生长及钾吸收的影响[J]. 植物营养与肥料学报, 2019, 25(7): 1173-1184.
[9] 李奇伟, 卢颖林, 周文灵, 等. 低钾胁迫对甘蔗不同品系生长和光合特性的影响[J]. 甘蔗糖业, 2011(6): 1-5.
[10] 周文灵, 卢颖林, 敖俊华, 等. 不同基因型甘蔗耐低钾能力、光合特性和抗氧化酶系统的差异[J]. 广东农业科学, 2015, 42(24): 41-46.
[11] 江 永, 敖俊华, 卢颖林, 等. 湛江市甘蔗“3414”肥料效应试验[J]. 广东农业科学, 2011, 38(19): 69-72.
[12] 董廣蕊, 石佳仙, 侯藹玲, 等. 甘蔗基因组研究进展[J]. 生物技术, 2018, 28(3): 296-301.
[13] 阙友雄, 黄文华, 许莉萍, 等. 甘蔗杂交后代遗传变异评价及高糖和低糖池构建[J]. 热带作物学报, 2009, 30(6): 811-816.
[14] Damon P M, Osborne L D, Rengel Z. Canola genotypes differ in potassium efficiency during vegetative growth[J]. Euphytica, 2007, 156(3): 387-397.
[15] Li Z, Yuan L, Wang Q, et al. Combined action of anti-oxidant defense system and osmolytes in chilling shock-induced chilling tolerance in Jatropha curcas seedlings[J]. Acta Physiologiae Plantarum, 2013, 35(7): 2127-2136.
[16] 徐 云, 袁 青, 胡华冉, 等. 低钾胁迫下不同大麻品种的耐性差异研究[J]. 中国麻业科学, 2016, 38(4): 156-161.
[17] 曾巧英, 江 永, 黄 莹, 等. 不同甘蔗品系对养分胁迫响应的差异研究[J]. 广东农业科学, 2015, 42(12): 38-43.
[18] 杜 琪, 赵新华, 王华杰, 等. 低钾胁迫对玉米干物质和养分积累与分配的影响[J]. 沈阳农业大学学报, 2017, 48(3): 257-264.
[19] 王晓光, 李春红, 赵新华, 等. 低钾胁迫下不同大豆品种钾吸收利用效率的差异分析[J]. 沈阳农业大学学报, 2013, 44(1): 7-12.
[20] 吴宇佳, 张 文, 肖彤斌, 等. 缺钾对不同基因型香蕉根系分泌物产生及土壤钾活化的影响[J]. 西南农业学报, 2017, 30(3): 624-628.
[21] Berglund A H, Quartacci M F, Liljenberg C. Changes in plasma-membrane lipid composition: a strategy for acclimation to copper stress[J]. Biochem Soc Trans, 2000, 28(6): 905-907.
[22] 王晓光, 王 岩, 李兴涛, 等. 低钾胁迫对大豆叶片膜脂过氧化及保护酶活性的影响[J]. 中国油料作物学报, 2010, 32(4): 512-517.
[23] Fatima R A, Ahmad M. Certain antioxidant enzymes of Allium cepa as biomarkers for the detection of toxic heavy metals in wastewater[J]. Sci Total Environ, 2005, 346(1-3): 256-273.
[24] Bhaduri A M, Fulekar M H. Antioxidant enzyme responses of plants to heavy metal stress[J]. Reviews in Envi-ronmental Science and Bio/Technology, 2012, 11(1): 55-69.
[25] 卢颖林, 曾巧英, 黄 莹, 等. 低钾胁迫对斑茅抗氧化酶活性和脯氨酸含量的影响[J]. 甘蔗糖业, 2018(3): 9-14.
责任编辑:白 净
收稿日期 2020-03-19;修回日期 2020-05-06
基金项目 广东科学院建设国内一流研究机构行动专项资金项目(No. 2020GDASYL-20200302005);国家甘蔗产业技术体系(No. CARS-20-1-4);广东省重点领域研发计划项目(No. 2019B020238001)
作者简介 曾巧英(1973—),女,博士,副研究员,研究方向:甘蔗抗逆生理与遗传机制。*通信作者(Corresponding author):齐永文(QI Yongwen),E-mail:yongwen2001@126.com。
