高羊茅FaFT基因表达、蛋白互作及生物学功能分析
2021-05-14罗维牟琼舒健虹吴佳海王小利
罗维 牟琼 舒健虹 吴佳海,3 王小利
(1. 贵州大学动物科学学院 高原山地动物遗传育种与繁殖教育部重点实验室,贵阳 550025;2. 贵州省农业科学院草业研究所,贵阳 550006;3. 贵州省农业科学院果树科学研究所,贵阳 550006)
开花是植物从营养到生殖阶段的关键时刻,是一个复杂的发育调控过程,并受内外因素的影响。植物需要适当的外界环境条件,如适宜的光周期和温度,在一定时间内刺激开花[1]。温度、光照等条件适宜时,植物叶片产生的成花物质或信号传递到茎尖的生长点,在顶端分生组织内(shoot apical meristem,SAM)启动开花基因的表达,促使植物进入成花决定态[2]。
基于对模式植物拟南芥的研究表明,在植物的成花转变中有多种途径,包括赤霉素途径、自发途径、春化途径及光周期途径[3]。不同途径能感受不同的外界信号刺激,共同调控植物开花过程。FLOWERING LOCUS T(FT)是重要的开花促进因子,研究证明 FT基因编码的蛋白质产物是长距离转运的成花素,属于磷脂酰乙醇胺结合蛋白基因(phosphatidylethanolamine binding protein,PEBP)家族,该家族成员在植物中是高度保守的调控因子,在被子植物和裸子植物中发现PEBP家族进化出开花抑制子和活化子。在适宜的光照下,FT基因受光周期诱导在叶片中表达,其表达产物经维管束转移到茎顶端分生组织调控下游基因表达促进开花。FT蛋白到达SAM后,与bZIP转录因子FLOWERING LOCUS D(FD)相互作用,FT /FD复合系统激活了许多MADS盒基因,包括APETALA1(AP1),FRUITFULL(FUL) 和 SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1(SOC1)过度表达CONSTANS(CO)刺激花器官的启动[4]。FT基因位于开花促进基因CO基因的下游,在光周期途径中,CO基因受到光周期节律GIGANTEA(GI)基因的诱导,CO基因表现出节律变化,从而激活 FT基因的转录和表达[5]。春化途径中,FLOWERING LOCUS C(FLC)基因是春化途径的关键基因,它是一个开花抑制子,既能在叶片中抑制 FT 基因的表达,也在茎顶端生长点抑制 FD 和 SOC1基因的表达,能影响茎顶端分生组织对来自叶片 FT蛋白信号的应答能力。但春化后,FLC基因的表达被抑制,这时在长日照条件下,CO基因通过上调 FT基因表达,进而促进SOC1和AP1基因 mRNA的积累而启动开花[5-6]。在甜菜(Beta vulgaris)中,春化涉及到两个相互拮抗作用的FT基因,BvFT1抑制BvFT2基因的表达,BvFT2 基因促进植物开花。春化后,BvFT1 基因的表达被抑制,BvFT2 基因表达促进植物开花[7]。在小麦(Triticum aestivum)和大麦(Hordeum vulgare)中,春化作用调控开花抑制子[8],使得 VERNALIZATION1(VRN1)基因表达上调,VERNALIZATION2(VRN2)基因表达下调,进而使得 FT基因表达促进植物开花[9-10]。FD基因同样参与调控植物开花进程,研究发现丧失功能的拟南芥fd突变体表现出晚花,当FD过量表达时可以帮助植物正常开花。FD基因能促进FT发挥作用,表现为fd突变体过量表达FT基因并没有早花表型[11]。
目前FT基因已经在多种植物中被分离出来,如拟南芥(Arabidopsis thaliana)[12]、水稻(Oryza sativa)[13]、洋葱(Allium cepa)[14]、二穗短柄草(Brachypodium distachyon)[1]、 陆 地 棉(Gossypium hirsutum)[15]等,而在高羊茅的成花机制研究甚少。高羊茅由于耐热性好、抗逆性强,现已广泛应用于草坪种植和牧草栽培,对草坪业和畜牧业有重要意义。本研究从前期高羊茅转录组数据中克隆出FaFT基因并对其进行生物信息学和亚细胞定位分析;运用qRT-PCR分析该基因在不同光照处理下及不同发育时期下的表达情况;应用酵母双杂交方法对互作蛋白FD进行探究,为进一步探讨高羊茅的成花机制提供依据。
1 材料与方法
1.1 材料
本实验选用的“黔草1号”高羊茅为贵州省农业科学院草业研究所2005年培育品种(国审:299)。TRIzol购自美国Invitrogen公司,cDNA合成试剂盒购自TaKaRa公司,所用相关载体由实验室保存,其他试剂为国产分析纯。
1.2 方法
1.2.1 FaFT基因在高羊茅基因组中扩增与序列分析 所取样品参照TRIzol说明书提取总RNA,cDNA第一链合成参照TaKaRa试剂盒。根据“黔草1号”基因组,用Primer 5.0设计扩增引物FaFT-F1/FaFT-R1进行目的序列扩增(表1),PCR反应程序:4 μL dNTP Mixture(0.0025 mol/L)、5 μL 10×PCR Buffer(Mg2+)、2 μLcDNA、3 μL MgCl2(0.025 mol/L)3 μL、0.25 μL TaKaRa Taq 酶、上下游引物FaFT-F/FaFT-R 各 2 μL, 补 ddH2O 至 50 μL。 反 应条件为预变性94℃ 2 min,35个循环(94℃ 10 s,55℃ 30 s,72℃ 30 s),72℃ 延伸 10 min。待反应结束后用1%的琼脂糖凝胶检测PCR扩增产物,并胶回收试剂盒收回目的片段,筛选纯化后送宝生物工程(大连)有限公司测序,测序结果用Blast数据库分析。

表1 引物信息Table 1 Primer sequence
1.2.2 FaFT基因表达分析 筛选大小相近的高羊茅种子经消毒后平铺在1/2MS固体培养基上,待长出子叶后移栽至装有蛭石和土壤的小花盆中(蛭石和土壤比例为1∶1),将小花盆放入不同光照培养箱。长日照(22℃,16 h/8 h)、短日照(22℃,8 h/16 h),最后分别在长日照和短日照基础上转入光照/黑暗与黑暗/光照循环模式转入黑暗/光照循环模式。以GAPDH(glyceraldehyde-3-phosphate dehydrogenase)为内参基因,将保存好的cDNA稀释20倍用于qRT-PCR分别检测高羊茅叶片中FaFT基因在长日照和短日照、长日照转、短日照转连续黑暗和连续光照、昼夜颠倒处理下以及长日照下不同生长发育阶段的表达量。每个样品3个技术重复,3个生物学重复,采用2-ΔΔCt方法分析数据。
1.2.3 亚细胞定位分析 利用双酶切将FaFT的编码区克隆到1300-GFP表达载体上,构建FaFT-1300-GFP重组质粒。将FaFT-1300-GFP重组质粒转化到农杆菌感受态细胞GV3101,筛选阳性菌落接种于LB(含 100 μg/mL Rif)28℃培养基震荡培养 24 h,离心后收集菌体。再加入0.1 mol/L的乙酰丁香酮2 μL和 0.5 mol/L MES 100 μL,28℃摇床培养OD值约1.0左右。4 000 r/min常温离心10 min,15 min收集菌体,用含0.01 mol/L MgCl2重悬菌体至OD值为1.0,以每毫升菌液加入2 μL 0.1 mol/L AS,静置3 h以上。将侵染液装入5 mL注射器内,用拇指按压注射器反板将液体从叶片下表皮注射到烟草叶片内,72 h后,取样通过激光共聚交荧光显微镜检测荧光信号。GFP激发光为488 nm,DAPI激发光为405 nm。
1.2.4 酵母双杂交 构建FaFD-pGADT7和FaFTPGBKT7载体,利用PEG/LiAc介导法将阴性对照pGBKT7和pGADT7、阳性对照pGADT7和pGBKT7-53和实验组FaFD-AD和FaFT-BD同时转入酵母菌株AH109。将转化后的酵母细胞加入0.9%NaCl溶液悬浮后的菌液涂布于SD/-Leu/-Trp(缺陷型-亮氨酸-色氨酸)培养基,待SD/-Leu/-Trp培养基上长出克隆后,挑取克隆至SD/-Ade/-His/-Leu/-Trp(缺陷型腺嘌呤-组氨酸-亮氨酸-色氨酸)上。30℃恒温箱培养3-5 d后进行X-α-Gal检测。
1.2.5 双分子荧光互补验证(bimolecular fluorescence complementation,BiFC) 将 FaFT基因的编码区构建至pSPYCE-35S载体,所用引物为FaFTC-F :5′-GCCTGGCGCGCCACTAGTGGATCCA-3′,FaFT-C-R:5′-CATCCCGGGAGCGGTACCCTCGAG-3′;同样将FD基因编码区构建至pSPYNE-35S载体。将FaFT-pSPYCE-35S和FD-pSPYNE-35S转入农杆菌GV3101中,将携带表达载体的农杆菌菌株注入烟草叶片,培养72 h后激光共聚焦显微镜下观察黄色荧光蛋白(YFP)信号。
1.2.6 FaFT基因生物学功能验证 FaFT基因表达载体构建:构建pCAMBIA1300-35S-FaFT 过量表达载体:根据终载体pCAMBIA1300-35S的图谱设计Kpn I-XbaI为插入位点并合成引物并扩增,操作步骤按试剂盒说明书进行。PCR反应体系及条件:2×PCR Buffer for KOD FX Neo 25 μL,0.01 mol/L dNTP Mix 1 μL,KOD FX Neo(1U/μL)1 μL,cDNA 第 一链 5 μL,primer F(10×)1.5 μL,primer R(10×)1.5 μL,PCR-Grade Water 15 μL ;预变性 98℃ 5 min,35个循环(98℃ 10 s,55℃ 30 s,68℃ 30 s),68℃ 延伸10 min。经检测合格后与pCAMBIA1300-35S载体进行体外连接转化,再将转化出来的单菌落通过摇菌后用空载体的通用引物进行菌液PCR鉴定,最后将经过菌液PCR鉴定的阳性菌液进行测序。
农杆菌介导的拟南芥遗传转化:常规方法种植野生型拟南芥Ler和突变体ft拟南芥:选取饱满的野生型拟南芥种子,先用70%的无水乙醇消毒,震荡离心后用无菌水冲洗,重复5次。再用0.7%的次氯酸钠溶液震荡混匀后静置15 min,离心吸除上清液,继续用无菌水冲洗3次。消毒完毕后将种子晾干,均匀点播在MS培养基上,封口。将MS培养基置于光照培养箱中(光照16 h/d,黑暗8 h/d,温度22℃)。待生长一周后,拟南芥幼苗转移到营养土与蛭石比为1∶1的花盆中。定期浇水。将农杆菌菌液扩大培养60 mL。4 500 r/min,10 min离心菌体,用侵染液重悬菌体并使得菌体的OD600到达0.8左右。将pCAMBIA1300-35S-FaFT过量表达载体转化野生型拟南芥Ler和突变体ft获得FaFT基因过量表达植株和FT基因功能补偿转基因植株。取生长状况良好的野生型拟南芥(花序较多的植株,未结荚)的花序浸入到菌体侵染液中3 min。结束后让侵染株系在暗环境下处理1 d。次日转移到正常生长条件下。相隔一周后侵染第2次。正常生长浇水等待收获T0代种子。每个处理及对照各9株,记录转基因株和对照株开花时间,莲座叶数量。
教师在写英语作文评价时要有针对性,应该因人而异,因材施教。学生的英语基础不一样,衡量的标准也应不同,教师一定要把握好评价的尺度。做到既要纠正学生的错误,也要鼓励学生继续写作。不同水平的学生也应该采取不同的评价标准,对于英文写作水平较高的学生,要表扬中有指正,对于中等水平的学生,教师要以鼓励为主,对于那些不擅长写英语作文的学生,要尽量从内容或者形式上,找出其闪光点,比如对几个准确的词语,几句符合语法规则的句型做出标识,并加以赞扬,激起学生写作的兴趣。
转基因植株的阳性初步筛选:收获的T0代种子经过烘干无菌消毒洗涤后均匀铺具有潮霉素抗性(40 mg/L)的MS培养基上,春化3 d后转移到正常条件下生长。观察根的生长。一般阳性株会正常发芽、生根、叶片舒展,而非转基因株系只有发芽,无根、无叶片。挑取选择培养基上的正常生长的幼苗转移到不加抗性的MS培养基上,待植株生长3-5 d左右可移栽到小花盆土中筛选T1代植株种子,常规种植T1代转基因拟南芥种子,继续筛选获得T2代植株。每个处理及对照各9株都置于培养箱中培养(光照16 h/d,黑暗8 h/d,22℃),记录转基因株和对照开花时间和莲座叶数量。
2 结果
2.1 FaFT基因的序列
测序结果(图1)显示,高羊茅FaFT基因有完整的cDNA序列,全长1 073 bp,有长度为537 bp的开放阅读框,编码178个氨基酸残基。

图1 FaFT基因的核苷酸序列及氨基酸序列Fig. 1 Nucleotide sequence and amino acid sequence of FaFT
2.2 高羊茅开花FaFT基因的表达分析
2.2.1 FaFT基因在长日照和短日照条件下昼夜表达节律分析 从图2-A可知,FaFT基因在长日照(Long-day,LD)条件下具有规律的昼夜震荡且周期约为24 h。2个光周期内,在开灯后ZT4-ZT8这一阶段表达量上升,到ZT8达最大值随后逐渐下降。FaFT基因在短日照(short-day,SD)下的表达与LD下表达模式基本相似,表达高峰出现的时间都在ZT8,但表达量较LD低(图2-B)。

图2 长日照(A)、短日照(B)下高羊茅FaFT基因的表达水平Fig. 2 Expression level of FaFT gene in tall fescue under long day (A) and short day (B)
2.2.2 长日照和短日照条件下转入连续光照和连续黑暗条件下FaFT基因的昼夜表达分析 与LD相比,把高羊茅从长日照转入连续光照后(LD-LL),FaFT基因的表达量急剧下降,在第1个光周期内ZT4-ZT12表达量很低,几乎检测不到,但仍维持一定的昼夜节律。不同的是表达峰值出现在ZT0,2个光周期内的表达模式基本维持一致(图3-A)。转入连续黑暗(LD-DD),与(LD-LL)的表达模式基本相近,FaFT基因表达量下调,表达节律减弱,2个光周期内表达峰值同样出现在ZT0(图3-B)。短日照转入连续光照SD-LL下(图3-C),FaFT基因表达的昼夜节律不明显。第1个光周期的表达量高于第2个光周期且有2个表达峰值,分别是ZT4和ZT20。第2个光周期的表达峰值出现在ZT12,时间晚了4 h。SD-DD(图3-D)下,FaFT基因表达模式与SD相似,但表达峰值出现时间较之晚了4 h,出现在ZT12,且表达量也基本相近。
2.2.3 光照/黑暗与黑暗/光照循环模式转入黑暗/光照循环模式FaFT基因的昼夜表达分析 由图可知,光照/黑暗处理(图4-A),高羊茅FaFT基因在第一个光周期有2个表达高峰,分别是ZT8和ZT20,第2天的表达量有所上调,但在两个光周期的变化趋势相似。由光照/黑暗处理转入黑暗/光照后(图4-B),其表达量急剧降低,两个光周期的表达高峰出现的时间分别在ZT16和ZT20。D/L下(图4-C),FaFT在两个周期的表达形式相似,第1个光周期ZT0-ZT8表达量很小,几乎检测不到,但都在ZT20达到表达高峰。由D/L转入L/D后(图4-D),FaFT的表达高峰在ZT4,较其他3种情况早。
2.2.4 不同生长发育阶段FaFT基因的昼夜表达节律分析 长日照条件下,FaFT基因在不同发育时期叶片中的表达模式基本相同,在各个发育时期叶片中都有表达。苗期(图5-A)、分蘖期(图5-B)、孕穗期(图5-D)3个时期的表达规律基本一致,表达量相近,并且2个光周期内表达高峰出现的时间都出现在ZT4。抽穗期(图5-C)FaFT基因在孕穗期叶片中的表达量高于其他3个时期,2个光周期内表达高峰出现在ZT12、ZT16,比其他3个时期的表达高峰出现的时间晚。总的来说,FaFT基因在长日照条件下4个不同时期的表达模式基本相同,呈现一定的昼夜节律性。
2.3 FaFT的亚细胞定位分析
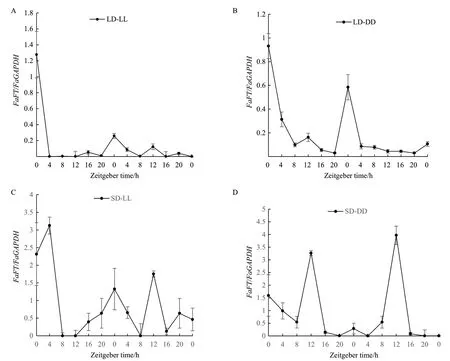
图3 长日照转连续光照(A)和连续黑暗(B)、短日照转连续光照(C)和连续黑暗(D)高羊茅FaFT基因的表达水平Fig.3 Expression level of FaFT gene of tall fescue from sunlight to continuous light (A) and continuous darkness (B) based on long day, continuous light (C) and continuous darkness (D) based on short day
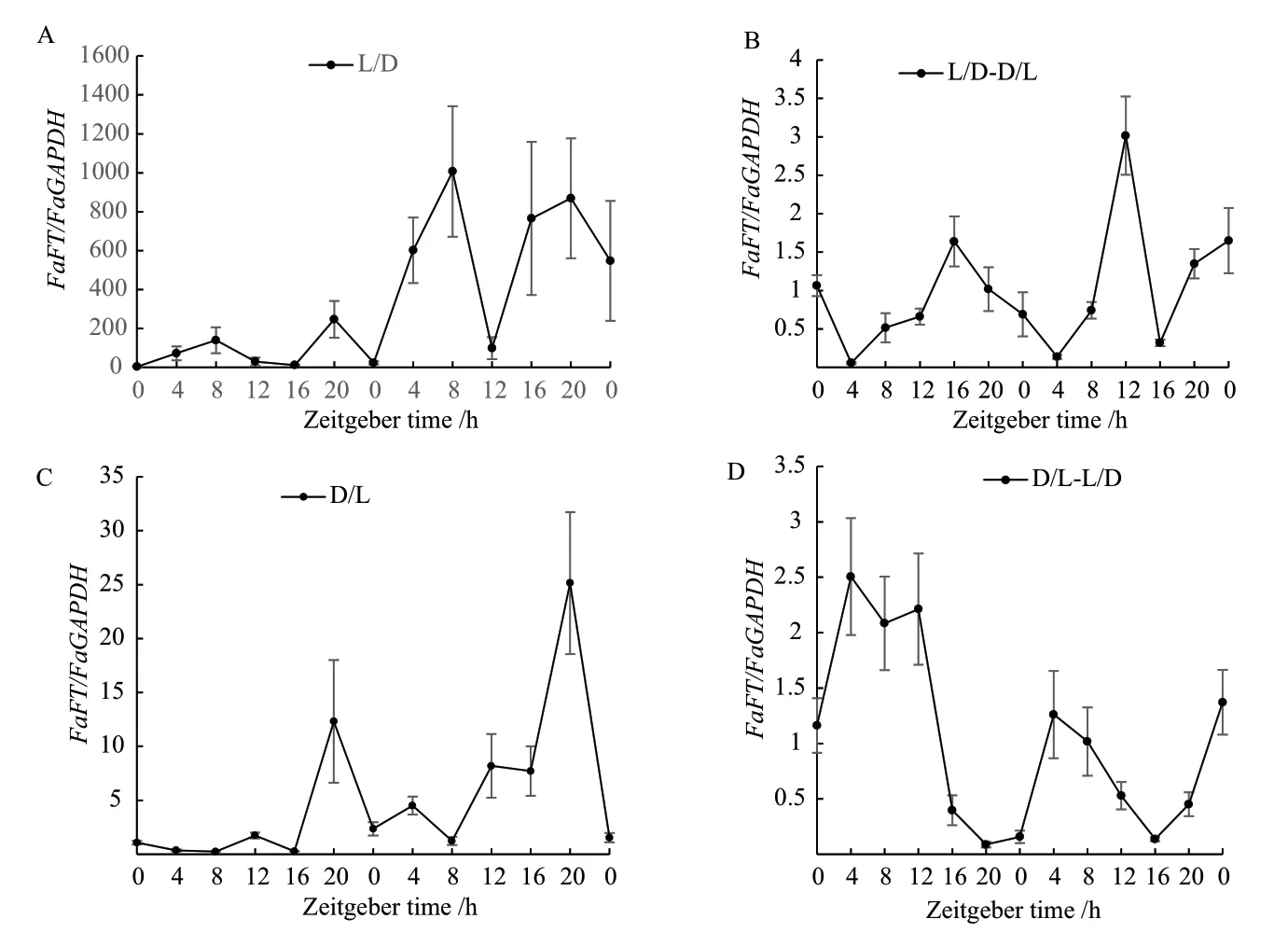
图4 高羊茅在光照/黑暗(A)、光照/黑暗-黑暗/光照(B)、黑暗/光照(C)、黑暗/光照-光照/黑暗(D)下FaFT基因的表达水平Fig.4 Expression level of FaFT gene in tall fescue under light/dark (A), light/dark-dark/light (B),dark/light (C), dark/light-light/dark (D)

图5 高羊茅FaFT基因在苗期(A)、分蘖期(B)、孕穗期(C)、抽穗期(D)的表达分析Fig. 5 Expression analysis of the FaFT gene of tall fescue at the seedling stage (A), tillering stage(B), booting stage (C) and heading stage (D)
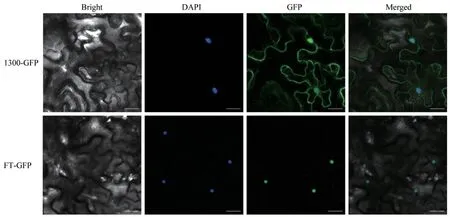
图6 FaFT在烟草表皮细胞中的表达Fig.6 FaFT expression in tobacco epidermal cells
2.4 FaFT互作蛋白FD的初步验证
FT一般与FD形成复合物发挥作用,利用酵母双杂交技术探索FT与FD之间的相互作用,并用相应的培养基来检测是否转化成功。如图7所示,酵母双杂交结果显示所有共转BD和AD载体的AH109菌株在二缺培养基上都长出了克隆,说明共转化成功;将SD/-Leu/-Trp培养基上的克隆挑取至SD/-Ade/-His/-Leu/-Trp平皿中培养发现,除了共转化阳性对照质粒的AH109长出克隆外,而FaFD/FaFT实验组也长出克隆;将SD/-Leu/-Trp上的克隆挑取至含有 X-α-Gal的 SD/-Ade/-His/-Leu/-Trp 上培养发现除了共转化阳性对照质粒变蓝,FaFD/FaFT实验组也变蓝,此实验结果说明FaFD蛋白和FaFT蛋白有潜在互作。

图7 FaFT与FD相互作用的筛选Fig.7 Screening of interaction between FaFT and FD
2.5 BiFC验证
利用BiFC技术进一步验证FaFT与FD互作的真实性。将FaFT-pSPYCE-35S与YFP的N-端片段(1-155aa)融合,将FD-pSPYNE-35S与YFP的C-端片段(156-239aa)融合,通过农杆菌介导转化在烟草表皮细胞表达。结果如图8所示,当FaFT与FD共表达时,在表皮细胞中可观察到黄色荧光信号,仅有FaFT或FD时并未检测到黄色荧光信号,说明FaFT蛋白和FD蛋白能发生相互作用。该结果与酵母双杂交结果一致。
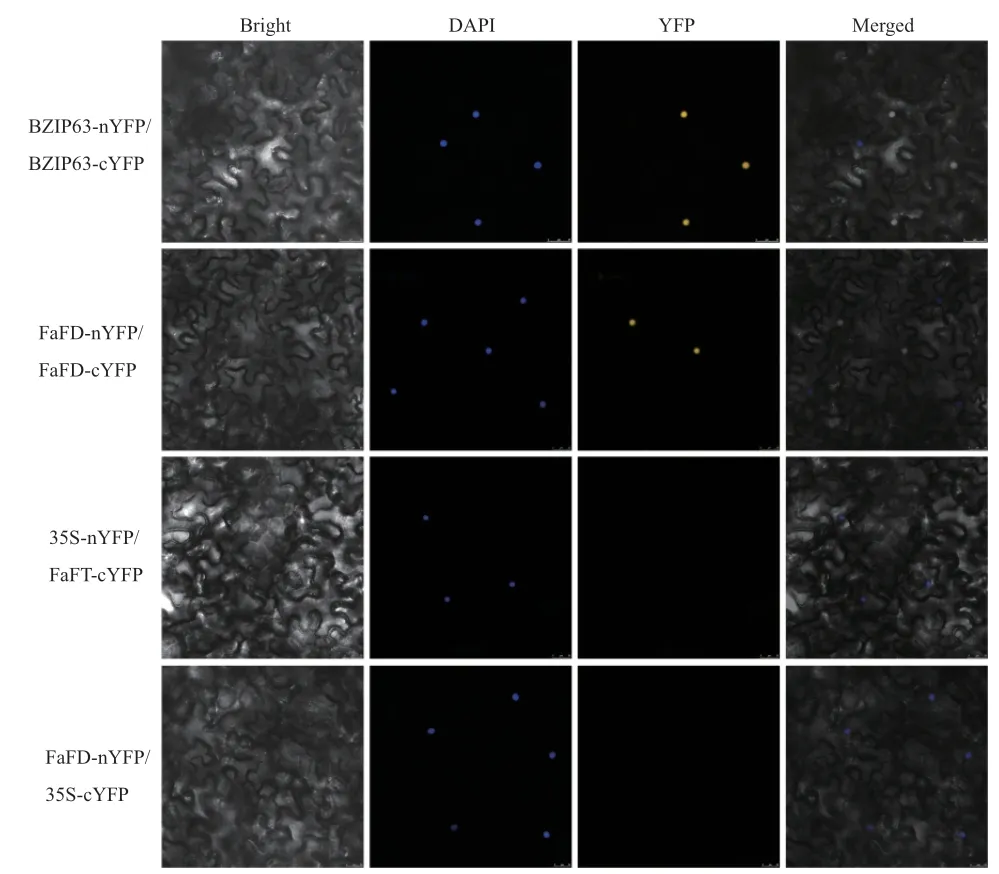
图8 FaFT与FD的BiFC实验验证Fig. 8 BiFC experimental verification of FaFT and FD
2.6 高羊茅FaFT基因促进拟南芥开花
构建pCAMBIA1300-35S-FaFT过量表达载体,转化拟南芥野生型Ler和突变体ft,长日照条件下,FaFT基因过量表达拟南芥植株表现出早花,功能补偿后的突变体ft则恢复了野生型的开花表型,突变体ft本身没有开花现象,一直停留在营养生长期,这表明FaFT基因促进拟南芥提前开花,恢复了ft突变体的晚花表型。
将拟南芥野生型和转基因植株种植在长日照条件下,结果(图9)显示:在长日照条件下,转FaFT基因拟南芥比野生型Ler开花提前约4 d,比ft突变体功能补偿植株早3 d左右;与野生型莲座叶9.89±1.7相比,过表达FaFT基因拟南芥莲座叶数量为11.2±1.41,转基因突变体ft莲座叶数达到9.1±0.73植株开花。这也表明FaFT基因能促进拟南芥开花。
3 讨论
FT蛋白是促进植物由营养生长到生殖生长转变的关键因子,能够整合多条开花途径。学者对拟南芥[16]、水稻[17]、二穗短柄草[18]和小麦[2]开花基因FT的研究发现,FT蛋白的基本功能非常保守,主要参与植物开花的诱导。高羊茅FT基因cDNA全长1 073 bp,开放阅读框537 bp,编码178个氨基酸残基,其与黑麦草、节节麦等禾本科牧草的氨基酸序列相似性较高,有很近的亲缘关系[19]。通过亚细胞定位得出高羊茅FT基因编码的蛋白定位在细胞核,在细胞核中发挥生理作用,也说明FaFT基因进化保守,与黑麦草、节节麦等禾本科牧草FT基因同源,可能有相似的功能。

图9 转基因拟南芥的开花时间、莲座叶数量及表型Fig.9 Flowering time, rosette leaf number and phenotype of transgenic Arabidopsis
光照是影响植物开花转变的重要因素,在合适的光照下FT基因受CO基因的诱导在叶片中表达。拟南芥在LD下能启动开花机制,长日照下FT基因的mRNA的表达量随着发育时间而逐渐增加,Kong等[20]在大豆中克隆了10个FT同源基因,其中GmFT2a和GmFT5a的表达量最高且受生物钟调控,表现出一定的昼夜节律。这两个基因在短日照条件下光照后ZT4时表达量最大,长日照开花时间晚于短日照,但二者都能促进开花。在小麦中,存在3个春化基因,VNR1、VRN2和VRN3,VRN3是与拟南芥FT基因同源,受春化和长日照的诱导从而促进开花;水稻Hd3a是拟南芥FT的同源基因,在LD下的表达量随着春化时间逐渐上升[2]。本研究对高羊茅不同光照处理下的转录水平分析得出,FaFT基因在长日照、短日照下的表达量不同,长日照下表达量高于短日照,但表达高峰都出现在有光照时期;在LD-LL和LD-DD两种条件下,FaFT基因的表达量相比LD、SD下有所降低,但表达高峰出现的时间一致(ZT0);SD-LL下,FaFT基因的表达出现2个表达高峰(ZT4和ZT0),SD-DD下的表达高峰延迟在ZT12;L/D、L/D-D/L、D/L、D/L-L/D昼夜颠倒后,虽然FaFT基因的表达高峰出现的时间不一致,但都能维持一定的规律,表现为“双峰型”。这也说明高羊茅FaFT基因受光周期的调控。同样,LD下,FaFT基因在不同发育时期的叶片中都有表达,在孕穗期叶片中的表达量最高。如果改变光照时间,FaFT基因叶片中的表达情况是否一致,这有待于进一步研究。
FT蛋白在叶片合成后经叶脉到达顶端分生组织(SAM)与bZIP转录因子FD相互作用,FT/FD复合物激活许多MADS盒基因启动开花。14-3-3蛋白是动植物中保守的一个调节分子家族,可以与水稻顶端芽中的FT同源Hd3a直接相互作用,形成功能性复合物,转运至细胞核[21]。因此,14-3-3蛋白可以充当支架蛋白,从而将FT与FD结合以触发开花。本研究为验证FT蛋白FD的作用,用酵母双杂交和BiFC验证了高羊茅FaFT蛋白和FD蛋白发生相互作用的真实性。
蔡宇鹏[22]在大豆中发现ft2a突变体无论是长日照还是短日照均表现为晚花,过表达GmFT2a植株开花时间都比野生型和突变体植株早,ft2a突变体的表达模式与野生型相似呈“双峰型”,只是表达量很低。当把野生型植株先置于短日照13 d后转入长日照,野生型植 株开花时间比LD下早,晚于SD,即短日照诱导产生的开花促进物质能在长日照下促进开花。同样光照处理的突变体植株并未恢复晚花表型。为验证FT基因的功能,本文成功构建FaFT基因过表达载体并转入野生拟南芥Ler和突变体ft,均成功获得转基因植株。LD下,FaFT基因过量表达拟南芥植株表现出早花,突变体ft恢复了野生型的开花表型,这表明FaFT基因促进拟南芥提前开花,恢复了ft突变体的晚花表型,成功验证了该基因的功能。
4 结论
从高羊茅转录组数据中成功克隆得到FaFT基因,该基因在不同光照处理下的表达能维持一定的昼夜节律,受光周期的调控。亚细胞定位发现FaFT基因编码的蛋白定位在细胞核;通过酵母双杂交和双分子荧光互补实验得出高羊茅FaFT蛋白能和FD蛋白发生相互作用;FaFT基因过表达在长日照下能促进拟南芥提前开花,且恢复突变体拟南芥开花。
