取食不同寄主植物的灰飞虱对药剂敏感性和解毒酶活性的影响
2021-05-14温胜芳王雪婷薛燕楠张国福夏晓明
丁 杰,温胜芳,王雪婷,薛燕楠,张国福,金 岩,夏晓明*
(1. 山东农业大学植物保护学院,山东泰安 271000;2. 山东省农药检定所,济南 251000)
灰飞虱(small brown planthopper,SBPH)Laodelphaxstriatellus(Fallén)属半翅目飞虱科,主要分布在亚洲和欧洲的温带和亚热带地区,在我国长江流域和黄淮地区为害严重。灰飞虱世代周期短,1年可发生5~6代,各世代可在不同寄主间转移危害,除取食水稻外,还可危害麦类、玉米等禾本科植物,同时田间禾本科杂草也可以为灰飞虱提供充足的营养条件(汪恩国,2007;乔慧等,2009)。灰飞虱除直接为害造成损失以外,主要传播水稻条纹叶枯病(rice stripe virus, RSV)和黑条矮缩病(rice black-streaked dwarf virus, RBSDV)等病毒病而造成水稻、小麦和玉米产量严重损失(Otukaetal.,2010;Zhangetal.,2010)。
由于寄主植物体内的次生物质对植食性昆虫体内的解毒酶具有诱导作用(王沫等,2003),同时不同寄主所含有的营养成分和次生代谢物质不同,所以当昆虫取食不同寄主植物时,其体内的羧酸酯酶(CarE)、谷胱甘肽-S-转移酶(GST)和多功能氧化酶(MFO)等解毒酶活性会发生变化(吕朝军等,2007;李晓磊和刘长明,2009;张睿等,2011;尹飞等,2013;Dermauwetal.,2018)。当昆虫以不同寄主植物为食时,会影响昆虫体内这些酶的活性以扩大寄主植物范围(Wangetal.,2010)。除酶活外,昆虫解毒酶基因的表达水平也受不同寄主植物的影响,且因寄主植物种类而异(Huangetal.,2018;Daietal.,2019)。害虫体内参与植物次生物质代谢和对杀虫剂起解毒作用的酶系是相同或相似的(姚洪渭等,2002;谢佳燕等,2007),因此,寄主植物介导的解毒酶基因表达和解毒植物次生代谢物和化感物质所涉及的酶活性的变化可能影响昆虫的适应性,并影响昆虫对杀虫剂的敏感性(Taoetal.,2012)。并在小菜蛾Plutellaxylostella(尹飞等,2013)、斜纹夜蛾SpodopteralituraFabricius(Xueetal.,2010;Karuppaiahetal.,2016)、B型烟粉虱BemisiatabaciGennadius(Castleetal.,2009;Xieetal.,2011)、朱砂叶螨Tetranychuscinnabarinus(戴宇婷,2013)等多种害虫中得到证实。
为探索取食不同寄主植物对灰飞虱的影响,本研究选取了灰飞虱的常见寄主水稻(紫香糯2315和武育粳3号)、小麦(农大26和济麦22)及稗草,将灰飞虱在这5种寄主上连续饲养3代后,测定了其对不同药剂的敏感性以及常见解毒酶活性的变化。研究结果可为灰飞虱的抗性发展研究提供依据,并为灰飞虱的田间科学用药和综合治理策略的制定提供指导。
1 材料与方法
1.1 供试虫源
灰飞虱种群2017年5月采集自山东省鱼台县水稻良种繁育基地的小麦田,于RXZ智能型人工气候培养箱中,在塑料盒(35 cm×25 cm×15 cm)中用未接触农药的紫香糯2315水稻饲养繁殖。水稻苗每10~15 d更换一次,以确保灰飞虱可以获得充足的营养。试验进行之前灰飞虱已在实验室在不接触药剂的条件下饲养20代以上。饲养条件为温度27±1℃,相对湿度75%±5%,光周期L ∶D=16 h ∶8 h。
1.2 供试寄主植物
供试5种寄主植物包括2个水稻OryzasativaL.品种(紫香糯2315和武育粳3号),由山东省水稻研究所提供;2个小麦TriticumaestivumL.品种(农大26和济麦22),由山东农业大学农学院提供;1种禾本科杂草:稗草EchinochloacrusgalliBeauv.,采自山东省鱼台县水稻良种繁育基地。5种寄主植物均为灰飞虱的常见寄主。将灰飞虱分别转接到供试的5种寄主植物上,按照1.1的饲养条件连续饲养3代后,选取3龄灰飞虱若虫进行药剂敏感性和解毒酶活性测定。
1.3 供试药剂及试剂
94%溴氰虫酰胺原药(cyantraniliprole,上海杜邦农化有限公司),96.4%吡虫啉原药(imidacloprid,山东潍坊润丰化工股份有限公司),96%噻虫嗪原药(thiamethoxam,山东潍坊润丰化工股份有限公司),10%三氟苯嘧啶悬浮剂(triflumezopyrim,美国杜邦公司),丙酮(天津市大茂化学试剂厂),α-乙酸萘酯(α-NA)(上海麦克林生化科技有限公司),固蓝RR盐(上海麦克林生化科技有限公司),对硝基苯甲醚(p-NA)(上海麦克林生化科技有限公司),1-氯-2, 4-二硝基苯(CDNB)(上海麦克林生化科技有限公司),还原型谷胱甘肽(GSH)(北京索莱宝科技有限公司),还原型辅酶Ⅱ(NADPH)(北京索莱宝科技有限公司),NaH2PO4(天津市凯通化学试剂有限公司),Na2HPO4(天津市凯通化学试剂有限公司),乙二胺四乙酸(EDTA)(Amresco公司),苯甲基磺酰氟(PMSF)(BBI公司),苯基硫脲(PTU)(北京精细化工厂),BCA蛋白浓度测定试剂盒(上海碧云天生物技术有限公司)、全自动酶标仪 680 型(Bio-Rad 公司)、Neofuge 13 R高速冷冻离心机(上海力申科学仪器有限公司)。
1.4 生物测定
参照班兰凤等(2015)和张凯伦等(2020)报道的稻苗浸渍法。用丙酮(10%三氟苯嘧啶悬浮剂用去离子水稀释)将供试药剂溶解,配制成1×104mg/L的母液。根据预试验结果,将上述母液用0.1%吐温80依次稀释成一系列浓度。将6日龄左右的水稻苗连根一起在系列浓度的药液中浸10~15 s,取出沥至无液体滴下,自然晾干后以湿脱脂棉包住根部放入透明的试管中。用吸虫器将试虫移入试管(2 cm×20 cm)中,每管15头,每个浓度3个重复,共45头,然后用纱布封口,以防湿度过大增加死亡率和灰飞虱逃逸。待试虫全部爬上稻苗或试管壁后,剔除机械损伤的个体,并补足15头。接虫后把试管放入温度27±1℃,光周期为16 h ∶8 h(光照 ∶黑暗)的培养箱中饲养。处理3 d后,检查死虫数,以不含药剂的处理做空白对照。
1.5 酶活测定
1.5.1酶液制备
取不同寄主上的3龄灰飞虱若虫20头,用1 mL磷酸缓冲液(0.1 mol/L,pH7.6,含1 mmol/L EDTA,1 mmol/L PMSF,1 mmol/L PTU和20% 甘油)冰浴匀浆,于4℃,10 000 rpm条件下离心15 min,上清液即待测酶液。
1.5.2酯酶(esterases,ESTs)活力测定
参照Han等(1998)的方法,并略做修改。将20 mg固蓝RR盐和0.2 mL 100 mmol/L α-乙酸萘酯(α-NA)加入到10 mL 0.2 mol/L, pH6.0的磷酸缓冲液中,振荡混匀,过滤得到底物和显色剂的混合液。在96孔酶标板中每孔加入20 μL经0.1 mol/L pH7.6磷酸缓冲液稀释10倍的酶液和200 μL底物和显色剂的混合液。用酶标仪在波长450 nm下记录光密度值,酶促反应在27℃下进行,每隔30 s记录一次,反应15 min。
1.5.3谷胱甘肽-S-转移酶(glutathione-S-transferases,GSTs)活力测定
参照Kao等(1989)的方法。在96孔酶标板中每孔加入50 μL 0.6 mmol/L 1-氯-2,4-二硝基苯(CDNB)和100 μL 6 mmol/L还原型谷胱甘肽(GSH),最后加入100 μL酶液。用酶标仪在波长340 nm下记录光密度值,酶促反应在27℃下进行,每隔20 s记录一次,反应15 min。
1.5.4多功能氧化酶(mixed-function oxidase,MFO)活力测定
参考Shang and Soderlund(1984)的方法。在96孔酶标板中每孔加入50 μL 1.0 mmol/L 对硝基苯甲醚(p-NA)和50 μL 1.0 mmol/L还原型辅酶Ⅱ(NADPH),最后加入100 μL酶液。用酶标仪在波长405 nm下记录光密度值,酶促反应在27℃下进行,每隔20 s记录一次,反应15 min。
1.5.4蛋白质含量的测定
使用BCA蛋白浓度测定试剂盒制作标准曲线,根据标准曲线和使用的样品体积计算出样品的蛋白浓度。
1.6 数据处理
使用SPSS 13.0软件进行统计分析,计算出供试药剂对不同寄主上的灰飞虱的毒力回归方程式、LC50值及其95%置信限、b值及其标准误,并采用Tukey’s test比较不同寄主植物上灰飞虱的解毒酶活性差异。
2 结果与分析
2.1 取食不同寄主植物灰飞虱对药剂敏感性的影响
采用稻苗浸渍法,测定取食不同寄主植物的灰飞虱3龄若虫对三氟苯嘧啶、吡虫啉、噻虫嗪和溴氰虫酰胺的敏感性,结果见表1。从表1可以看出,连续取食不同寄主植物3代后,灰飞虱3龄若虫对三氟苯嘧啶的敏感性无差异,但对吡虫啉、噻虫嗪和溴氰虫酰胺的敏感性存在一定差异。其中,取食2个品种小麦的灰飞虱对吡虫啉和溴氰虫酰胺最敏感,取食2个水稻品种和稗草的灰飞虱对吡虫啉和溴氰虫酰胺敏感性相对较低。取食水稻武育粳3号的灰飞虱对噻虫嗪最敏感;取食2个品种小麦的灰飞虱对噻虫嗪相对较敏感,取食稗草和水稻紫香糯2315的灰飞虱对噻虫嗪敏感性最低。
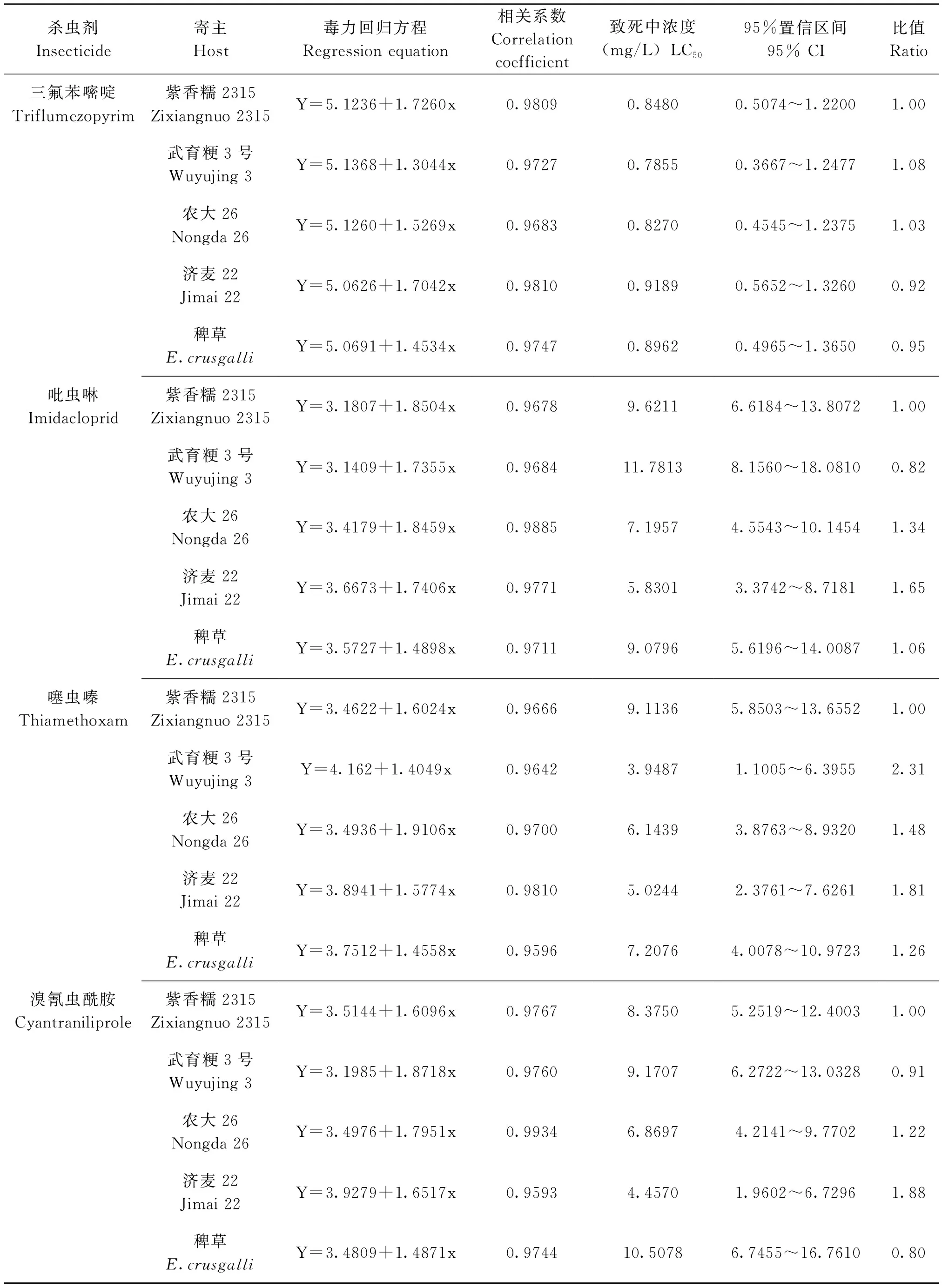
表1 取食不同寄主的灰飞虱对4种杀虫剂的敏感性
2.2 取食不同寄主植物灰飞虱的解毒酶活性比较
连续取食不同寄主植物3代后,灰飞虱3龄若虫体内的3种解毒酶活性存在明显差异(表2)。取食稗草的灰飞虱解毒酶活性要明显高于取食小麦的灰飞虱解毒酶活性。其中,取食稗草的MFO和ESTs活性均最高,取食2个品种小麦的MFO和ESTs活性均最低;取食2个品种小麦的ESTs活性差异显著,但MFO活性没有差异,取食2个品种水稻的MFO和ETSs活性差异显著(P<0.05)。GST活性从大到小依次为紫香糯2315>稗草>济麦22>武育粳3号>农大26,不同寄主之间差异显著(P<0.05)。

表2 取食不同寄主灰飞虱的3种解毒酶活性
3 结论与讨论
寄主植物诱导的昆虫对药剂敏感性变化通常有3种情况:敏感性上升、下降或基本保持不变(Wangetal.,2010;Xueetal.,2010)。已有研究发现,取食不同寄主后的朱砂叶螨对联苯菊酯的敏感性无变化(戴宇婷等,2013)。本研究也发现,连续取食水稻、小麦和杂草3代后,灰飞虱3龄若虫对三氟苯嘧啶的敏感性无变化,对吡虫啉、噻虫嗪和溴氰虫酰胺的敏感性变化也较小。但也有很多研究表明,取食不同寄主植物后,害虫对药剂的敏感性变化较大(Karuppaiahetal.,2016)。这种变化与寄主植物的营养条件、次生代谢物质的种类和含量、昆虫种类与发育阶段以及外界环境温度等条件有关(姚洪渭等,2002;张睿,2011),与供试药剂的种类也有关系(Xieetal.,2011)。本研究也发现,取食不同寄主植物后灰飞虱对不同药剂敏感性变化也存在不同。此外,诱导时间较短,这可能是寄主植物对灰飞虱药剂敏感性影响较小的一个原因。
寄主植物在影响害虫生长发育的同时也可以诱导昆虫体内的解毒酶系统发生变化(吕朝军等,2007;吕敏等,2012)。ESTs、GSTs和MFO均是昆虫体内重要的解毒酶,能够被不同寄主植物诱导,在对次生代谢物质和外源化合物的解毒代谢和对杀虫剂的抗性机制中起着重要的作用。本研究发现,用不同寄主植物饲养灰飞虱3代后,灰飞虱3龄若虫体内的ESTs、GSTs和MFO 3种解毒酶活性存在明显差异,取食稗草后的灰飞虱体内3种解毒酶活性要明显高于取食小麦的灰飞虱。尽管发现取食不同寄主植物后的灰飞虱对药剂的敏感性变化较小,但取食小麦后的灰飞虱对药剂的敏感性均要高于稗草的趋势。因此,这种变化趋势很可能是不同次生代谢物质诱导解毒酶活性变化导致。但是哪种解毒代谢酶起主要作用,还需进一步研究。
在包括中国的东亚地区,小麦和水稻轮作系统中的小麦、水稻和杂草均可为灰飞虱提供足够的食物(Huangetal.,2018;Lietal.,2019)。本研究结果表明,取食水稻、小麦和稗草后的灰飞虱的解毒酶活性存在差异,同时灰飞虱对杀虫剂的敏感性也发生了一定的变化。研究结果不仅对灰飞虱的田间综合防治具有重要的意义,也有助于我们更好的理解灰飞虱对寄主植物的生理生化适应机制。
