Variation analyses of controlled pollinated families and parental combining ability of Pinus koraiensis
2021-04-30ShuchunLiJiaqiLiYanyanPanXiangeHuXuesongNanDanLiuYueLi
Shuchun Li · Jiaqi Li · Yanyan Pan · Xiange Hu ·Xuesong Nan · Dan Liu · Yue Li
Abstract To evaluate and select elite Pinus koraiensis parent combinations and off spring families, 34 full-sib families were evaluated. Variance analysis of tree height, diameter at breast height and volume showed no signif icant diff erences among each block, the interaction of block, male and female. The family heritability of all traits were high(> 0.9). Phenotypic and genotypic coeffi cients of variance of height, diameter at breast height, and volume showed that the phenotypic coeffi cient of variation of volume in the same combination was higher than height and diameter at breast height, indicating that volume was the main factor determining excellent single plant selection. The female and male trees with the best general combining ability indicated that the optimal parental hybridization was not necessarily the optimal combination. In addition, the least parents of hybrid off spring performance were always relatively poor and should not be selected as hybrid parents. Additionally,we found there was no signif icant diff erence among the Effects of female and male parents by random modelling,but there were signif icant diff erences by f ixed modelling.The most signif icant Effect of female- and male interaction,and the selection of better female or male parents in the test parent group should be effi cient as well as the results of signif icant variation among them by f ixed modelling. For the evaluated breeding population of P. koraiensis, the backward selection of excellent parent combinations or forward selection of excellent individuals as progeny should be the major breeding strategies.
Keywords Controlled pollination · Progeny testing ·Pinus koraiensis · Parental combining ability
Introduction
Genetic tests play an important role in tree improvement programs and accelerating breeding cycles used to understand genetic structure and composition of quantitative traits, such as tree height, diameter at breast height, seed production.Genetic testing is involved in three broad programs associated with def ining genetic architecture to understand the quantitative genetic makeup of a population: (1) To evaluate the genetic worth of specif ic parents within a population by progeny testing; (2) To establish an advanced-generation base population from a breeding population; and, (3) To quantify the gains to estimate progress made in successive steps of the program. Based on enhancing steps in the breeding cycle, genetic tests have great use for advancing tree improvement programs. The genetic value of specif ic parents in the population are evaluated and the base population for the selection of superior germplasm for higher order improvement is constructed (White et al. 2007).
Controlled pollination is an important genetic test for evaluating elite progeny of coniferous species, and can provide a clear relationship for higher order improvement. In addition, the general combining ability (GCA) and special combining ability (SCA) of parents can be estimated by controlling pollination, which provides a basis information for backward selection. The analysis of controlling pollination and genetic combining ability is of signif icance for exploring the genetic variation of seed generation of species, and for the improvement of later generations.Pinus koraiensisSiebold & Zucc. is a member of the family Pinaceae widely spread in northeastern China, eastern Russia, the Korean peninsula and Japan (Vidakovic 1993; Barnes et al. 1992).The species can endure severe drought and persist on barren soil (Yang 2013) and is commonly used for ecological restoration in arid mountain landscapes of northern China(Zhang et al. 2014). In addition,P.koraiensisis a species with relatively high ecological and economically value (Lim 2012; Deng et al. 2015). It has excellent timber, straight trunk and edible pine nuts (Imbs et al. 1998; Nergiz and Donmez 2004; Zhang et al. 2015). Genetic improvement ofP.koraiensisbegan in the 1980s in China, and mainly based on natural forests. More than 20 clonal seed orchards have been established (Xia et al. 2016). Tissue culturing and other asexual propagation methods are diffi cult forP. koraiensismaterials, selecting excellent trees and establishing seed orchards are the main way to produce genetic improved seedlings ofP. koraiensis.Information on the genetic Effects of parents and combinations forP. koraiensisoff spring is not available. Furthermore, studies on the controlled pollinated progeny ofP. koraiensisand its parental combining ability are insuffi cient.
In this study, 14P.koraiensisclones were randomly selected from a breeding population and 34 full-sib families obtained by the factorial mating design. Tree height (H) and diameter at breast height (DBH) of 18-year-old trees were recorded. The specif ic objectives were to: (1) Understand the source of variation, genetic structure and parental inf luence of full-sib family growth traits; (2) Examine the interplant variation in full-sibs families; (3) Evaluate the general and special combining ability of parents and combinations,respectively; and, (4) Develop strategies forP.koraiensisbreeding improvement.
Materials and methods
Experimental sites
The experiment and progeny testing was conducted in the Naozhi forestry seed orchard (41° 05′ N, 126° 06′ E), with an average elevation of approximately 510 m, located in the western hillsides of Changbai Mountain, Linjiang City,Jilin Province. This seed orchard is in a temperate monsoon climate, mean annual temperature is 8.2 °C, precipitation 744 mm, and average frost-free days 128.
Materials and methods
One hundred and thirtyP.koraiensisclones were selected from local natural forests for measuring and the parents of 14 clones were randomly selected to estimate genetic variation of off spring. A factorial mating design was used to control pollination, and included seven male and seven female clones. The experiment was carried out in the spring of 1995, and seeds of 34 families collected in the autumn of 1996 (Table 1). Thirty-four full-sib off spring families were obtained and met the requirements of full-sibs. Each parent mated with the opposite parent three to six times which satisf ies the estimation of parental GCA.
All full-sib families were sown in 1997, and seedling management conditions were consistent. Seedlings were planted out in spring 2001. Planting sites were gentleslope, dark brown forest soil more than 50 cm deep. The experiment was randomized complete blocks with threereplications by a completely randomized block design in row plots containing six seedlings at 2.0 × 3.0 m spacing.
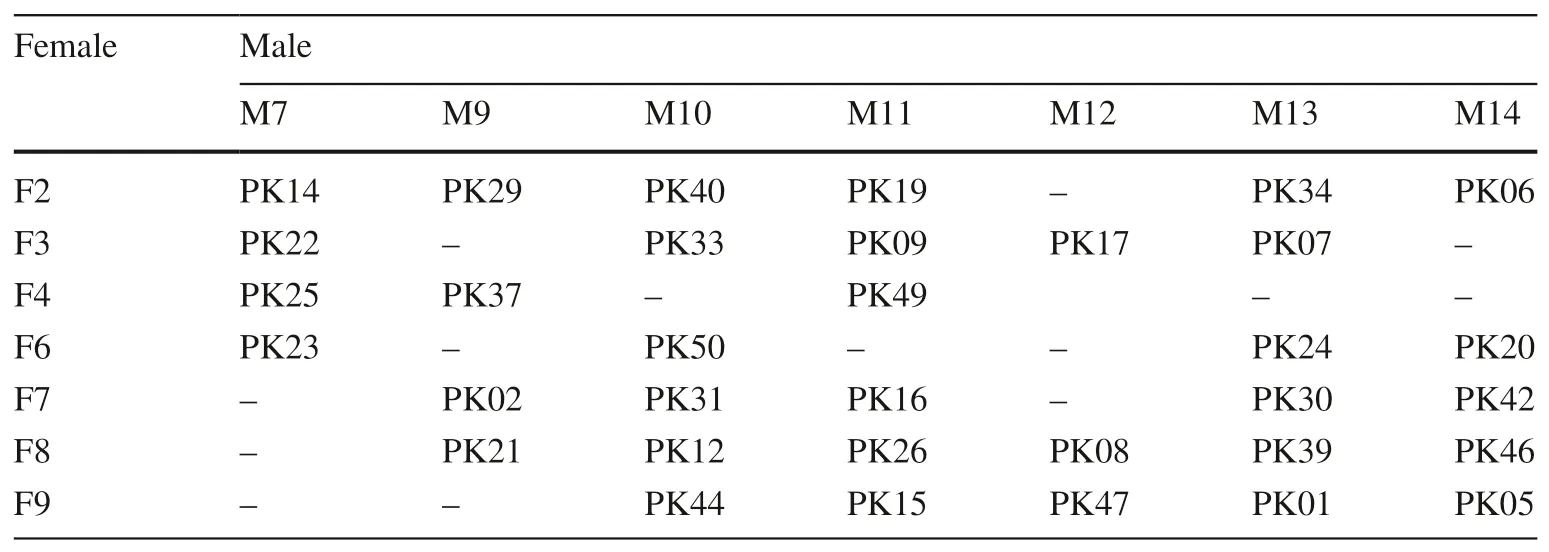
Table 1 Combination of 34 full-sib P. koraiensis families
Determination of index
Tree height (H) and diameter at breast height (DBH) of all families were measured in 2015 (19-year-old trees). Volume (V) was calculated following the method of Zhang et al.( 2016).
Statistical analysis
Statistical analysis of the growth indicators were surveyed with the linear model:
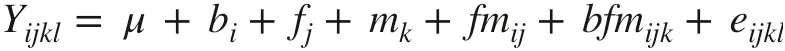
whereY ijklis the observed value of each experiment;μis the average value of the test;b jis the block Effect;f jis the female Effect;m kis the male Effect;fm ijis the combination Effect of parents;bfm ijkis the interaction Effect between block and combination;e ijklis the error within the plot. The female,male and full-sib Effects were tested by random and f ixed models, respectively. Additionally, the variance component was estimated according to Hansen and Roulund ( 1996).
Phenotypic coeffi cient of variation (PCV) and genotypic coeffi cient of variance (GCV) were estimated following Jonah ( 2011) using the formulas:
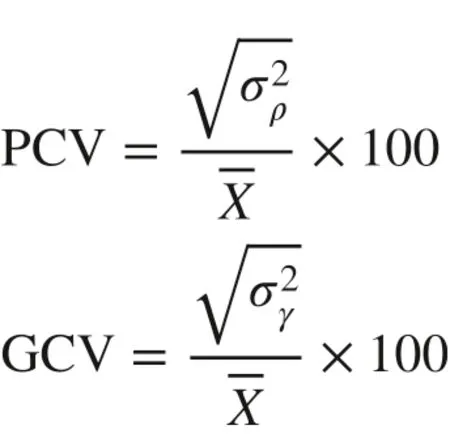


The estimation of heritability for a single plant was calculated as:
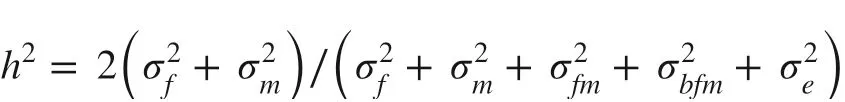
All data analyses were performed in SAS software (v.9.2).
Results
Genetic variation of traits
The ANOVAs of growth traits H, DBH and V were shown in Table 2. The estimated variance components of female × male were more than 70%, and the maximum gain could be obtained by selecting an excellent full-sibscombination, indicating the combination selection of the tested parent and full-sibs can obtain a degree of improvement gain.

Table 2 ANOVAs of tree height (H), diameter at breast height (DBH), volume (V) among diff erent variation resources
The results of random model analysis showed no signif icant diff erences in growth traits between male and female parents. The main genetic variance components were still derived from the combined interaction of female and male.The interaction of female and male had a higher variance component. The results showed that the genetic variation of the population mainly came from the full-sib Effect.
Genetic variations between combinations of various traits
Based on the analysis of variance results, H, DBH and V of diff erent hybrid combinations were analyzed (Table 3).The results show signif icant diff erences among hybrid combinations (P< 0.01); the families’ heritability of all traits was above 0.90, indicating high heritability. The phenotypic coeffi cient of variation (PCVs) of H, DBH and V of the families were 12.10%, 10.37% and 24.40%, respectively. The genotypic coeffi cient of variation (GCVs) of H, DBH and V of the families were 11.09%, 8.77% and 9.90%, respectively. PCVs and GCVs of H were higher than DBH; PCVs of V were twice as high as H and DBH. The relatively high amplitude of volume variation provided abundant resources of variation for selection among combinations, and higher genetic gain could be obtained by genetic selection based on the phenotype of volume traits.
The estimated heritability of H, DBH and V ranged from 0.30 to 0.39. There was a degree of genetic variation in the full-sib families which can be used selectively.The heritability of full-sib single plants was signif icantly lower than that of full-sib family, indicating that there was greater potential for improvement based on the performance of full-sibs.
Estimation of relative combining ability
The estimated results of relative general combining ability(RGCA) of all traits were shown in Table 4. Among the seven female trees, the H with positive dominance relative to RGCA was 3, DBH and V were 4, the best performance were F2 and F4 parents, and the poorest performers were F6, F7 and F8 parents. Among the seven male trees, the RGCA of M7 and M13 were the best performers, M9 and M12 parents were the poorest. The optimal H, DBH and V of the female trees were F4, F3 and F2, the poorest tree H and DBH was F9, the lowest volume was F6. The optimal H, DBH and V of the male trees was M11, the best DBH and V was M7,and the poorest was M12.
The results of relative special combining ability(RSCA) were shown in Table 5. The RSCA of H, diameter, DBHand V ranged from − 29.13 to 30.23, − 25.83 to 18.85, and − 48.00 to 58.22, respectively. The H, DBH and V of F9 × M14 in the full-sib family had the largest RSCA,and the minimum RSCA families were F2 × M14 (H) and F7 × M9 (DBH, V).

Table 3 ANOVAs of traits of P. koraiensis full-sib families
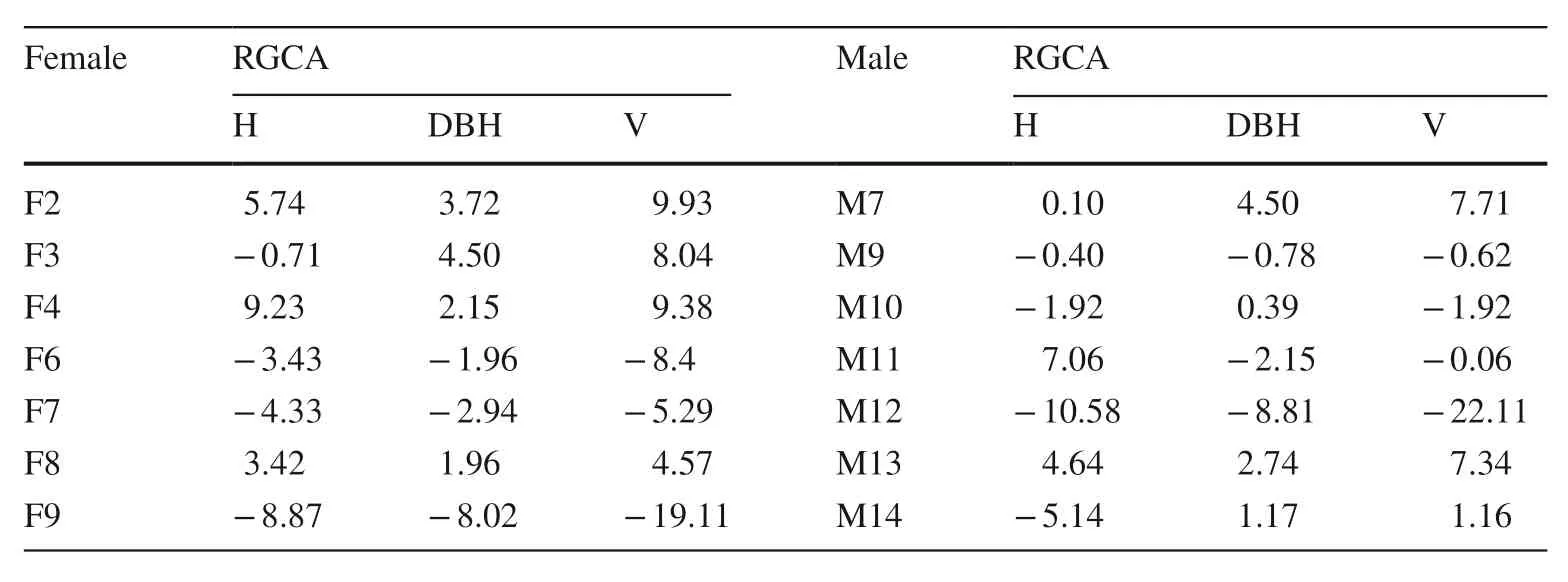
Table 4 RGCA of H, DBH and V of diff erent parents

Table 5 RSCA of H, DBH and V of diff erent full-sib families
Comprehensive evaluation of multiple traits
The Qi values of 34 families were shown in Table 6. With a selection rate of 10% based on growth traits, families 22, 5 and 40 were selected as elite families. The genetic gains of height, diameter and volume were 14.9%, 12.4%, and 34.9%,respectively.

Table 6 Comprehensive evaluation of multiple traits of all the families
Discussion
In this study, 34 full-sib families of aP. koraiensisbreeding population were selected and the variation and special combining ability between the full-sib combinations were identif ied as the main characteristics of the breeding population. Variation analysis is an important method to evaluate the range of variations in tree breeding (Zhao et al. 2015),and heredity and variation are the main components of forest tree breeding (Mwase et al. 2008). The coeffi cient of genetic variation ref lects the variation degree of the population and is an Effective indicator to measure the genetic variation potential of related growth traits (Gao 2010). In this study, there were no signif icant diff erences among female and male parents by random modelling, but there were signif icant diff erences by the f ixed model. The most signif icant female/male interactions and the selection of better female or male parents in the test parent group should be effi cient.The results of signif icant variation by f ixed modelling, and for the evaluated breeding population ofP. koraiensisthe backward selection of excellent parent combinations or forward selection of excellent individuals in progeny should be the breeding strategy.
To obtain the advantage of variation brought by parents, inter-provenance hybridization should be adopted; the question to be addressed in the next step. The phenotypic coeffi cient of variation (PCV) and the genotypic coeffi cient of variation (GCV) of height were higher than diameter at breast height, which is contrary to previous results on the evaluation of superb families ofLarix(Jia et al. 2016). This may be related to the rapid growth of height relative to diameter at breast height at this age. The phenotypic coeffi cient of variation was volume > height > diameter at breast height;the genotypic coefficient of variation, which accounted for 40.4%, 91.6% and 84.6% of the PCV, respectively. The results show that the variation of height and diameter are inf luenced by genetic factors; the selection potential for height and diameter at breast height was higher than that for volume, height, and diameter and were suitable factors for selection ofP. koraiensis. This result was not consistent with research onLarix kaempferi(Lamb.) Carr. (Pan et al. 2018) and may be due to diff erent species. The family heritability of height and diameter was higher than 0.90 which is signif icantly higher than that forPinus massoniana(Huang 2017), indicating that there are abundant genetic variations in height and diameter ofP. koraiensisfull-sib families which could be stable to the next generation. In addition, high phenotypic coeffi cients of variation and heritability values indicate that there would be less inf luence by environmental Effects and benef icial for the selection and evaluation of elite families (Maniee et al. 2009). Genetic gain is larger and the selection Effect better under certain conditions (Chen 2008).
Controlled pollination is the recombination of parent genes. The general combining ability (GCA) and special combining ability (SCA) of parents can be estimated by identif ication of parental traits in cross combinations and can provide a reference for the selection of parent combinations and the determination of parent genetic design (Qin et al.2011). From the results of the relative general combining ability (RGCA) and the relative special combining ability(RSCA), the female and male trees with the best RGCA were F2, F4 and M7, M13. But the combined off spring of RSCA is not the best, indicating that the optimal parental hybridization is not the optimum combination. In addition,the weakest parents of the hybrid off spring performance is relatively poor and should not be selected as hybrid parents;the parental hybridization with the best character was not necessarily the best combination. These results are in agreement with previous studies (Xu et al. 2015), for example, in cross-breeding. Parents with high GCA (at least one parent with high GCA) should be selected to obtain greater heterosis (Chen et al. 2016).
Conclusions
The tested breeding population ofP. koraiensishad higher genetic variation and special combining abilities and could promote genetic gain by backward selection for superior parental combination and forward selection for excellent progeny individuals. The single plant heritability in the fullsib family provides a scientif ic basis for forward selection of the improved generation. This study has increased information on the genetic determination of controlled coniferous pollination and has provided a scientif ic basis for developing forest management strategies forP. koraiensisseed orchards.
Publisher’s NoteSpringer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affi liations.
杂志排行

Journal of Forestry Research的其它文章
- Sacred groves of India: repositories of a rich heritage and tools for biodiversity conservation
- Relationship between H 2 O 2 accumulation and NO synthesis during osmotic stress: promoted somatic embryogenesis of Fraxinus mandshurica
- Changes in leaf stomatal traits of diff erent aged temperate forest stands
- Somatic embryogenesis and plant regeneration in Betula platyphalla
- Hydrogen peroxide as a systemic messenger in the photosynthetic induction of mulberry leaves
- Production and quality of eucalyptus mini-cuttings using kaolin-based particle f ilms