复叶千姿百态的奥秘
2021-04-29王超群贺亮亮陈江华
王超群,贺亮亮,陈江华†
①中国科学院西双版纳热带植物园,昆明 650223;②中国科学院大学,北京 100049
1 单叶和复叶的发育
叶是植物进行光合作用的主要部位,在形态上表现出极大的多样性,因此是研究植物复杂形态建成和演化的很好模型。自然界中的叶片包括单叶(simple leaf)和复叶(compound leaf)两种最基本的类型。单叶包含一个叶片单元;复叶由多个叶片单元,即小叶(leaflet)构成。从生物学功能上来说,每一个小叶等同于一个单叶,因此在植物演化历程中,复叶的出现被认为带来了一定的优势,如提高光合效率、减少植食性动物的损伤等[1]。根据复叶中小叶的数目和排列方式,复叶又可分为羽状复叶、掌状复叶、三出复叶和单身复叶等。此外,还有各种各样的多级复叶,如二回三出复叶、多回羽状复叶等。
叶的发育过程可以分为三个连续和交叉的事件:起始(initiation)、初级形态建成(primary morphogenesis)和次级形态建成(secondary morphogenesis)[2-3]。单叶和复叶都起始于茎顶端分生组织(shoot apical meristem, SAM)的侧翼。单叶的初级形态建成是一个相对短暂和简单的形态改变过程:建立叶最基本的结构框架,包括三维极性的建立,即近轴端-远轴端(也称基-顶轴向,proximo-distal)、近轴面-远轴面(也称背-腹轴向,adaxial-abaxial)、中-侧轴(mediolateral),以及叶各部分结构(叶柄、叶片和叶边缘等)初级形态的形成[2]。相比之下,复叶的初级形态建成包括一个特殊而复杂的发育过程,即小叶的形成,这一过程决定了复叶中小叶的数目和排列方式。不同植物中,保守或特异的遗传因子协同调控复叶的初级形态建成和小叶的时空起始,最终导致了复叶形态的复杂性和多样性(图1)。小叶起始后,利用与单叶发育类似的机制来建立其三维极性,并分化出各部分结构。次级形态建成过程是指在已分化出的叶的基本结构框架上,细胞进一步特化出具有功能的组织结构,如表皮细胞、栅栏组织、海绵组织和维管束组织等。次级形态建成在单叶和复叶中是比较保守的。下面着重探讨复叶初级形态建成的分子机制。
2 KNOXI与复叶发育
复叶发育的分子机制研究最早始于番茄(Solanum lycopersicum)。早在20世纪90年代,Hareven 等人将玉米KNOXI基因(class I Knotted1-like homeobox)KN1在番茄中错位表达(misexpression)导致了超级复叶(supercompound leaf)的发生[4]。最近20多年中,美国加利福尼亚大学Neelima Roy Sinha等人深入研究了KNOXI基因在不同物种复叶发育中作用的广泛性和保守性[5-6]。目前的研究表明,KNOXI调控复叶发育途径是存在于包括番茄在内的大多数植物中的保守机制,KNOXI的表达与复叶的形态之间有很高的相关性(图1)。KNOXI基因首先是SAM的特征基因,它通过在SAM区域的特异表达来维持其无限生长(indeterminancy)属性。单叶植物叶发育过程中,KNOXI基因在叶原基中检测不到表达;但是复叶植物中,叶原基从顶端分生组织侧翼起始后,KNOXI在复叶原基中的表达激活了其无限生长属性,维持复叶原基的形态发生活性(morphogenetic activity),从而促进小叶原基的发生。以色列希伯来大学的Naomi Ori和Yuval Eshed等人以茄科植物番茄为模式材料,鉴定出许多控制复叶形态发生活性的关键遗传因子,同时也揭示了生长素、细胞分裂素和赤霉素等重要植物激素对番茄复叶的可塑性发育调控十分重要[7-9]。这些研究极大地加深了对复叶形态多样性形成机制的认识。
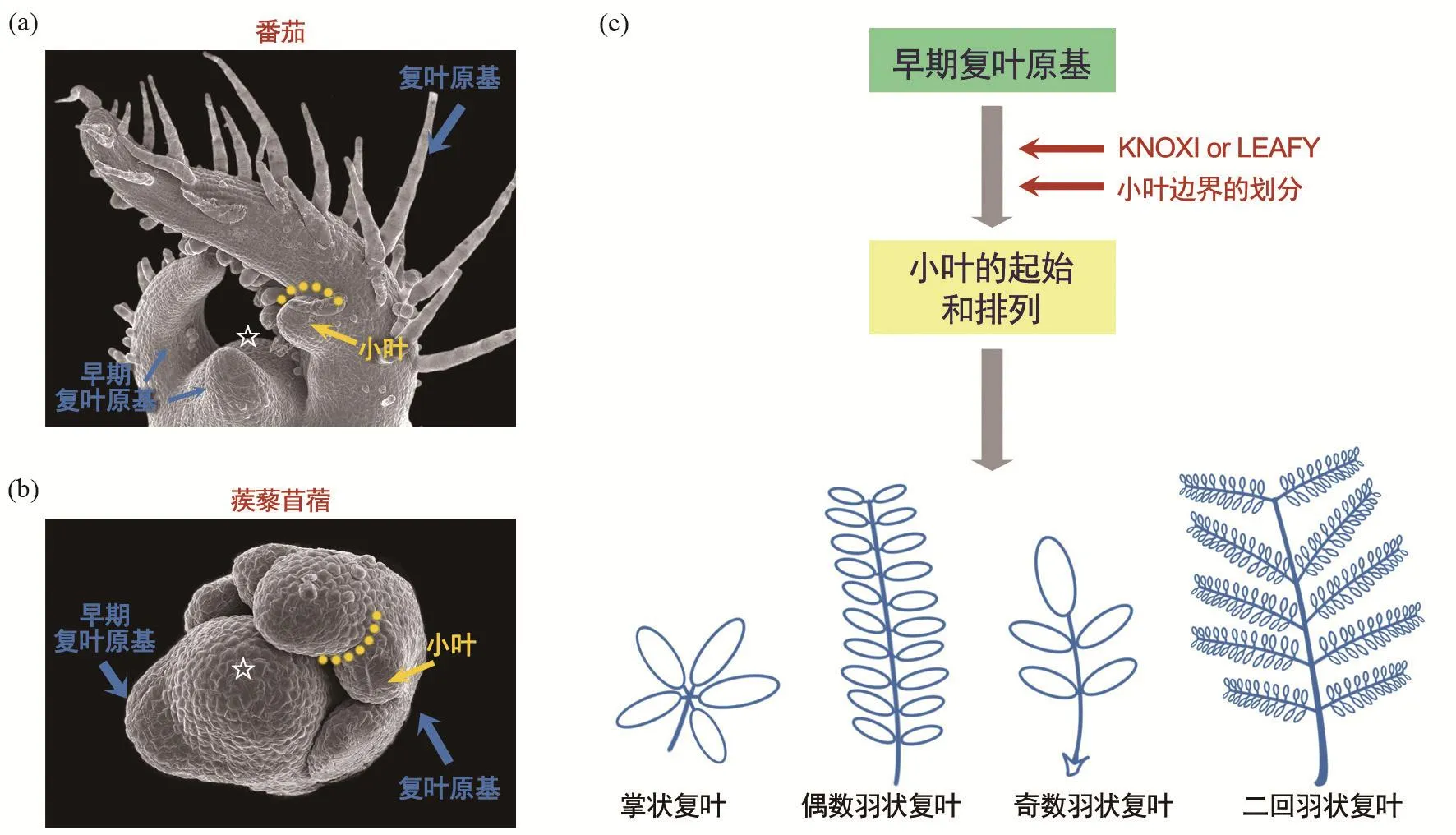
图1 复叶的发育过程:(a)番茄不同发育时期的复叶原基。(b)蒺藜苜蓿不同发育时期的复叶原基。箭头所示复叶原基和小叶原基;☆代表顶端分生组织;虚线代表小叶和小叶之间的边界。(c)复叶发育过程示意图
植物都会利用一些保守的因子来调控自身特有复叶形态的形成,但是有些植物也发展了一些物种特异因子来调控这个过程。近一二十年来,德国马克斯·普朗克植物育种研究所Angela Hay和 Miltos Tsiantis等人对十字花科拟南芥(Arabidopsis thaliana)和其近缘复叶物种碎米荠(Cardamine hirsuta)的叶发育进行深入和系统的比较研究[10-12]。碎米荠复叶形态建成也是由保守的KNOXI途径调控,但是其物种自身特异因子如HD-ZIP I Homeodomain家族基因REDUCED COMPLEXITY(RCO)也被招募来调控这个过程[13]。同时,他们结合经典的遗传学和最新的基因组学、植物成像和建模技术,展示了基因的活性如何通过影响细胞生长来调控叶器官形态建成,以及演化过程中如何改变器官的发育模式来决定复叶的最终形态的发生。这些研究进一步促进人们从多维角度,包括分子、细胞和演化等层面来理解复叶千姿百态形成的奥秘。
3 豆科植物的复叶发育
豆科植物呈现极其丰富的复叶形态多样性,几乎囊括了所有类型的复叶模式,并且有许多植物表现为单叶结构,如紫荆族分支和山蚂蝗属的部分植物[14]。豆科植物叶形态的多样性是系统分类学、形态发生学、进化发育生物学、遗传学以及古生物学等学科的重要研究方向之一。同时,豆科植物在农业和生态系统中发挥着举足轻重的作用,一系列重要农作物如大豆(Glycine max)、花生(Arachis hypogaea)等是人类和动物主要的植物蛋白及油脂来源。豆科多个模式植物已被用于复叶发育的研究,其中豌豆是最经典的用于复叶发育研究的遗传材料。在20世纪末至21世纪初,英国约翰英纳斯中心Julie Hofer和 Noel Ellis等人在豌豆(Pisum sativum)复叶发育的研究上做了系统的工作,克隆了很多叶形态突变体的基因,并提出豌豆复叶模式建成机制,为豆科植物的复叶发育机制研究奠定了基础[15-17]。在20世纪末,我国豆科植物的功能基因研究始于植物学家罗达先生,他为这一领域培养了一批优秀人才。
虽然KNOXI途径在大多数植物的复叶发育中具有广泛性和保守性,但在豆科植物的复叶发育调控机制上出现了分化[6,18]。目前已知,在属于豆科IRLC(inverted repeat-lacking clade)分支的复叶发育中,KNOXI基因的作用完全被LFY同源基因UNIFOLIATA(UNI)和SINGLE LEAFLET1(SGL1)所代替[15,19]。而一些non-IRLC豆科植物,KNOXI和LFY对于维持复叶原基的形态发生活性都发挥着重要的作用[6,20-21]。但是我们也要知道,目前对于non-IRLC豆科植物,只有LFY基因的功能缺失突变体,而没有KNOXI基因的功能缺失突变体报道,所以对于KNOXI基因在其复叶发育中究竟有何作用还不清楚。因此,探索KNOXI途径和LFY途径在豆科植物复叶形态多样性形成中的作用机理是非常有趣的科学问题。
豆科植物的复叶具有许多特化结构,如位于叶柄基部包裹着叶柄的托叶、在叶或小叶最基部的叶枕以及豌豆复叶中的叶卷须。这些特化结构都具有十分重要的生物学功能,为植物的生存提供了一些特殊的途径,如托叶起到保护叶芽的作用,叶枕是豆科植物重要的运动器官,而叶卷须适应了攀援运动的需要。对于这些特化结构的发育控制,植物往往进化出特有的调控途径,目前已取得部分研究进展[22-25]。未来对这些特化结构的发生与其生物学功能的分子机制的解析及其进化研究将是豆科生物学研究的热点。
4 蒺藜苜蓿的复叶模式建成研究
紫花苜蓿(Medicago sativa)是世界上最著名的优质牧草,也一直是奶业和畜牧业的核心原料,素有“牧草之王”的美称。四倍体紫花苜蓿的基础研究受到其本身遗传特性的限制,一直进展缓慢。最近,中国科学院西双版纳热带植物园陈江华团队联合西北工业大学王文团队和邱强团队完成的新疆大叶紫花苜蓿的全基因组测序和基因编辑,为未来紫花苜蓿的分子定向育种打开了一个新的窗口[26]。作为其近缘种的二倍体蒺藜苜蓿(Medicago truncatula)是豆科生物学研究的主要模式植物,它原产于地中海地区,为豆科苜蓿属一年生植物。野生型蒺藜苜蓿产生的第一片真叶是单叶,称为非成熟态叶;从第二片真叶开始均为成熟态叶,呈现三出复叶结构,其远轴端由一对侧小叶和一个顶小叶构成,近轴端为一对托叶包裹叶柄,侧小叶与顶小叶的基部均有感应昼夜节律变化的运动器官,即叶枕。自21世纪初,由于蒺藜苜蓿遗传转化效率高、基因组小并完成了全基因组的精确测序、自花授粉、生长周期短、结实率高并具有丰富的突变体资源等优势,其成为豆科生物学及叶发育生物学研究的模式材料[27]。
蒺藜苜蓿复叶发育的详细过程已被解析[19]。根据叶原基所处的叶间期(plastochron)可以定义叶的发育时期:最晚出现的叶原基为P1,次晚为P2……以此类推,将要出现的下一个叶原基为P0。不同时期叶原基具有特定的形态特征。在P1期,叶奠基细胞在SAM上的周边分生区凸起形成复叶共同原基,随后发育出托叶原基;在P2期,一对侧小叶原基和顶小叶原基分化出来,小叶原基之间形成边界,彼此分离;在P3和P4期,表皮毛陆续从小叶原基远轴面发生,而小叶原基远轴面生长发育速度快过近轴面,使小叶原基在近轴面形成凹陷,并进一步发育成折叠状;在P5期和随后的时期,托叶和侧小叶原基之间形成叶柄原基,侧小叶和顶小叶之间接着形成叶轴原基,至此建成三出复叶的基本结构。在此发育过程中,复叶的形态发生活性是如何维持和调控的,一直是人们关注的核心问题。
研究发现:蒺藜苜蓿LFY同源基因SGL1被定义为叶形态建成的无限生长决定因子(indeterminancy factor),其维持早期复叶原基的形态发生活性,同时也决定一对侧小叶的起始和发育;sgl1突变体侧小叶不能形成,突变体表现为单叶结构[19]。C2H2锌指蛋白PALMATE-LIKE PENTAFOLIATA1(PALM1)是叶形态建成的有限生长决定因子(determinancy factor),它特异地在侧小叶区域发挥作用,通过负调控SGL1的时空表达来抑制原基的形态发生活性;palm1突变体的5个小叶聚集在叶柄顶部,形成类掌状复叶模式[28]。最近发现的一个BEL1-like(BELL)亚家族同源异型结构域(homeodomain)蛋白PINNATELIKE PENTAFOLIATA1(PINNAI)也是叶形态建成的有限生长决定因子,它也是通过直接抑制SGL1的转录活性来调控叶形态发生活性。pinna1突变体的5个小叶以羽状方式排列,额外增生的两片小叶位于顶端小叶的基部,形成类羽状复叶模式;pinna1 palm1双突变体产生二级复叶,最多可形成13片小叶(图2)。PINNA1和PALM1通过协同作用(synergy)来调节SGL1基因在复叶发育过程中的时序表达,从而实现对复叶形态发生活性的精细控制,最终决定蒺藜苜蓿的三出复叶模式[18](图3)。

图2 蒺藜苜蓿不同突变体背景下的复叶模式。从左至右的突变体株系分别为elp1 sgl1双突变体、elp1单突变体、palm1 elp1双突变体、pinna1 elp1双突变体、pinna1 palm1 elp1三突变体
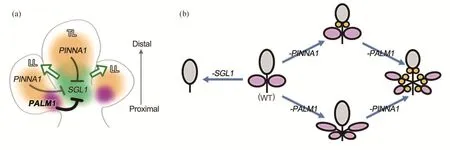
图3 蒺藜苜蓿复叶模式建立模型[18]
此外,蒺藜苜蓿的复叶模式建成过程中也涉及一些保守的信号途径和关键因子的调控,包括叶背腹极性控制基因和生长素等。最近研究人员发现控制叶片背腹极性的HD-zip III家族的REVOLUTA转录因子和控制生长素合成的YUCCA酶,也同时参与蒺藜苜蓿的复叶发育[29-30]。这些研究预示着复叶形态发生活性的维持和叶原基背腹极性的维持及激素信号途径之间有着千丝万缕的联系。
5 小叶边界划分的分子机制
复叶以共同原基的形式从SAM周缘区起始,而KNOXI或LFY是维持早期复叶原基分生能力的关键因子,决定小叶原基的起始能力。从小叶原基的起始到形成最终的完整复叶形态,还涉及一个关键的生物学过程——小叶原基与共同原基间、小叶原基与小叶原基间边界的划分。其中,NAM/CUC转录因子保守地调控不同植物复叶发生过程中的小叶原基间边界区的形成,相关基因的沉默或突变都能导致小叶不同程度的融合以及小叶数目的减少[31]。NAM/CUC基因在转录水平上受到miRNA164A的调控,碎米荠par突变体的miRNA164A功能缺陷,NAM/CUC基因的过量表达使小叶数目增多[32]。蒺藜苜蓿和番茄中都发现M类KNOX蛋白参与调控小叶边界的划分和小叶数目的维持[33-34]。由此可以理解:在复叶的形态建成过程中,小叶数目的维持和小叶边界划分之间存在着千丝万缕的联系;单叶和复叶的差异,不仅仅在叶原基分生组织活性的控制上,也在叶原基侧缘不同区域边界发育的控制上,植物有着精细而复杂的调控机制来控制复叶的不同形态。如果能进一步理解并阐明小叶数目的维持和小叶边界划分间的关系,将帮助我们更好地理解复叶形态建成的内在过程。
6 不同物种复叶发育的比较
对不同植物复叶发育过程的研究,可以看出不同植物复叶形成既有相同之处,又各具特点。复叶发育的核心事件包括早期复叶原基的建立、复叶原基形态发生活性的维持、小叶原基的起始、叶片三维极性的建成等。小叶原基是按特定的方式沿着复叶基-顶轴向有序起始的。根据小叶起始方式不同,复叶发育可以分为向基式(basipetally)、向顶式(acropetally)和分支式(divergently)三种发育模式[35-36](图4)。向基式发育是指复叶中靠近顶部的小叶原基最先起始,相邻的小叶再起始,最后是基部的小叶起始。典型的有羽状复叶的番茄复叶发育过程。向顶式发育是指复叶中靠近基部的小叶原基最先起始,最后是顶部的小叶起始。如:罂粟科花菱草,其基部侧小叶原基先起始,沿着基-顶轴向逐渐起始顶部的侧小叶原基;豌豆复叶的小叶形成也是向顶式发育,但是其顶端的卷须是向基式发育。分支式发育是向顶式发育和向基式发育的结合形式,如菊科蓍(shī)属植物,其复叶初级侧小叶为向基式发育,而二级侧小叶为向顶式发育。目前对于这些不同的小叶发育模式研究只有一些形态学和组织学方面的工作,未来深入研究相关的分子调控路径非常有意义,也能帮我们更好地理解复叶形态多样性的形成机制。
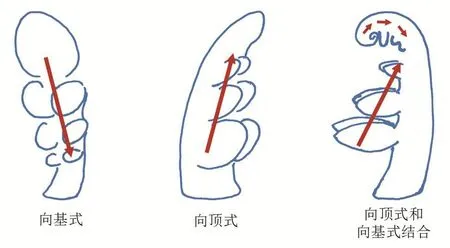
图4 复叶中小叶的发育方式,其中箭头代表小叶起始的顺序(修改自[36])
7 复叶的起源和演化
陆生维管植物叶的出现和早期演化是影响到整个地球生命的重要事件[37]。所有叶器官无一例外从茎顶端分生组织周缘区隆起发生,成熟后成为茎轴上特定形态的侧生器官。它是一种有限的器官,与理论上无限生长的茎顶端不同。叶原基的顶端生长在个体发育的早期就停止了,接着是组织的特化、增大并成熟。因此,人们要问:在植物演化的历史上,无限生长的茎顶端与有限生长的叶侧生器官之间的不同是如何产生的?换句话说,有限生长的叶器官是如何从无限生长的茎顶端起源的?
齐默尔曼(Walter Zimmermann)1930年提出的顶枝学说(telome theory)是迄今为止解释现代陆生维管植物叶(大型叶,megaphyll)起源的一个较好的学说,该学说认为大型叶起源于三维的分枝系统。原始的陆生植物为二叉分枝,两个分叉出的顶枝束是大小、形态相似的,其中一个顶枝束经历漫长的演化历程发展成片化的叶器官(图5)。一方面,KNOXI对维持顶枝束的无限生长能力是必需的,有限生长的叶器官的发生则必须抑制KNOXI基因的活性。有研究认为,ARP基因拮抗KNOXI基因的活性在叶的起源和演化过程中发挥着保守而关键的作用[38]。另一方面,BELL、KNOXII与KNOXI蛋白来自于共同的祖先,KNOXII与BELL互作拮抗KNOXI的功能对有限生长器官的属性维持很重要,这一机制可能对顶枝向叶器官的演化有着重要意义[39]。
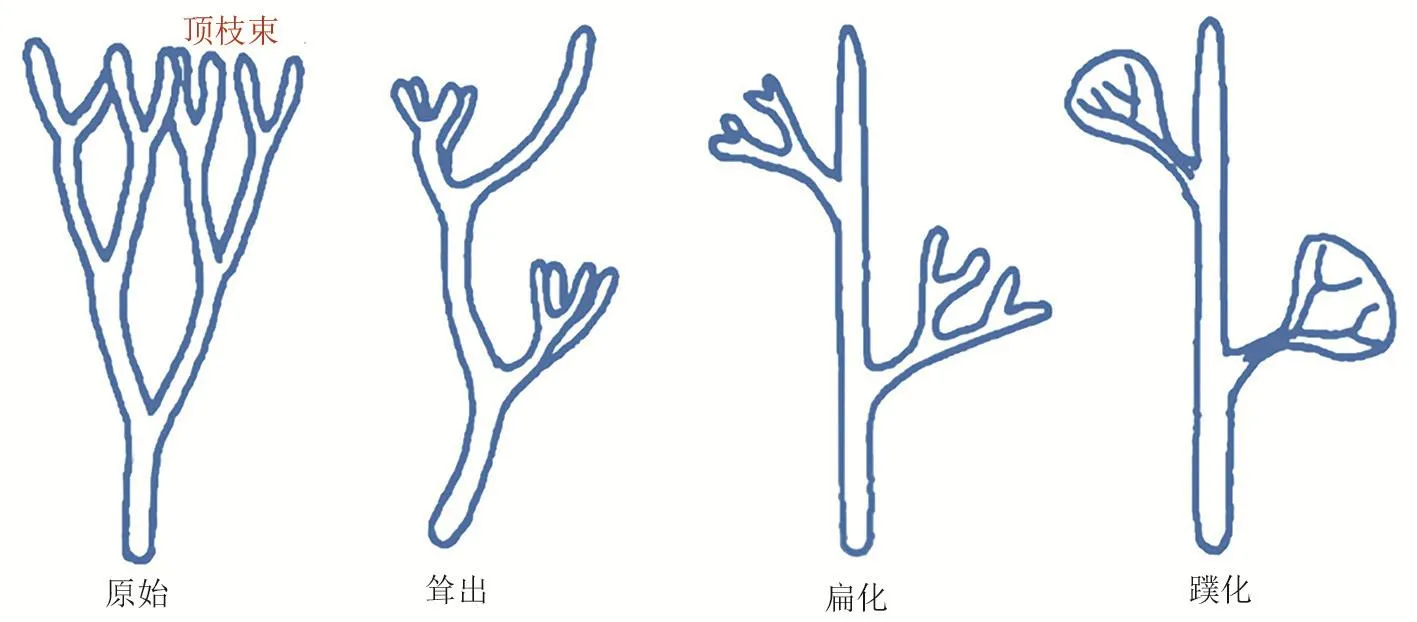
图5 原始的陆生植物都是二叉分枝的,两个分叉出的顶枝束是大小、形态相似的;在耸出的过程中,其中一束变长变粗而演化成主轴,相应地另外一束缩短成为侧枝;原本朝向空间不同方向的顶枝在演化的过程中逐渐生长到同一个平面上(扁化);位于同一平面内的各个顶枝以及中干被由薄壁细胞组成的一层组织连接到一起,形成一个独立的器官(蹼化)(图片修改自[37])
对于复叶的起源,化石证据表明被子植物的祖先的叶形态是单叶结构,复叶是在单叶的基础上多次演化的结果,并且一些复叶还会返回祖先的单叶形态。目前有两种假说来解释复叶和单叶的演化关系[40]:第一种假说认为复叶中的每一片小叶都等同于一片完整的单叶,复叶被认为是一个同时具有茎和叶特性的部分未决定的结构,因此,复叶可看作是一个退化的茎分枝;第二种假说认为整个复叶等同于一片完整的单叶,每片小叶被看作是单叶上的突起,因此复叶被认为是单叶的一种修饰(图6)。基于“顶枝学说”,Bharathan和Sinha等提出在被子植物进化过程中,其祖先的单叶原基由于建立了KNOXI或LFY基因的表达,使其属性由有限生长态(determinacy)转变为部分无限生长态,导致叶原基某些区域具有部分分生组织活性,这是复叶中小叶发生的基础[41]。然而,对自然界中丰富多样的复叶形态而言,仅仅依靠KNOXI或LFY基因的功能可能是难以完全实现的。如上所述,物种保守因子工作机制的多样性和物种特异因子的多样性都可能是复叶形态多样性形成的原因。
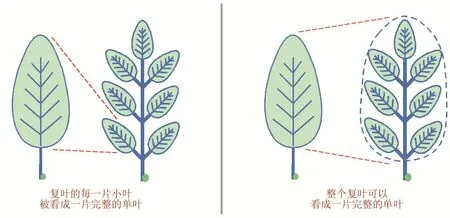
图6 复叶和单叶的进化关系假说,其中叶基部的绿色圆点代表腋芽分生组织(修改自[40])
在许多复叶植物中发现,一些基因的突变同时会影响到复叶和花的形态。LFY是花分生组织的特征基因,其在豆科植物复叶发育中的作用不言而喻,同时还发现LFY对碎米荠和番茄的复叶形态有重要作用[42-43]。UFO基因编码的F-box蛋白可以结合LFY蛋白,并导致LFY泛素化降解,因而对花的发育十分关键。无论碎米荠、豌豆、百脉根(Lotus japonicus)还是苜蓿,UFO都可能参与到复叶发育的调控中[17,43-44]。最近在蒺藜苜蓿中报道的一个AGFL基因也是同时控制了花和叶的发育[45-46]。从进化的观点来看,花被认为是变态的叶,这给我们一个启示,叶和花器官可能具有很多共同的机制来控制其复杂的形态建成。
8 展望
植物复叶的模式建成是一个极其复杂的过程,受内外环境因素、植物激素信号和植物发育过程中关键基因共同调控。基于已知的单叶发育和复叶发育中的分子机制的保守性和多样性认识,目前对于茄科植物番茄、十字花科植物碎米荠和豆科植物的复叶发育的研究取得较大的进展,但是与单叶发育研究相比较,复叶发育的研究仍处于初级探索阶段,其深入的遗传调控网络仍然很不清楚。特别是豆科植物,不同物种间其叶形态相差甚远,呈现出丰富的叶模式。如在豆科植物中既有完全单叶结构(如肾叶山蚂蝗Desmodium renifolium)、类单叶结构(如羊蹄甲等),又有羽状和掌状复叶,同时还有众多的多级复叶模式等,这给复叶发育的研究既带来挑战又带来众多有趣的科学问题。随着众多的植物基因组测序的完成以及新一代基因组编辑技术的不断成熟,相信叶片复杂形态建成的分子机制研究将得到极大发展。
