2份春小麦种质资源成株期抗条锈病基因遗传分析
2021-04-27李秋荣白耀博蒋礼玲魏有海郭青云
赵 珂,李秋荣,侯 璐,*,白耀博,蒋礼玲,魏有海,郭青云
(1.青海大学 青海省农林科学院 青海省农业有害生物综合治理重点实验室,青海 西宁 810016; 2.徐州生物工程职业技术学院 徐州现代农业生物技术重点实验室,江苏 徐州221006)
小麦条锈病由小麦条锈菌(Pucciniastriiformisf.sp.tritici)引起,是世界范围内影响小麦生产的最严重病害之一。从小麦一叶期到成熟期,小麦植株的绿色部位都可以发生病害,且在低温和潮湿条件下易发生[1]。我国西北、西南、黄淮海等冬小麦区和西北春小麦区的生产一直受小麦条锈病的威胁[2]。实践证明,种植抗病品种是防治该病害最为经济安全有效的方法[3]。对抗条锈病春小麦资源中抗病基因的遗传分析为抗病基因的合理使用提供前期理论基础,也为育种家提供抗病材料,加速抗病育种的进程。已有研究表明,我国存在较为丰富的小麦抗性资源,周强等[4-5]研究表明, 绵麦37成株期对条中32号小种的抗性主要受1对显性基因的控制, 同时受另外2对隐性基因的影响,绵麦39对条锈菌条中32的成株期抗性受一对显性基因的控制。邱亨池等[6]研究发现,秦农142成株期抗病性由1对显性基因和1对隐性基因共同决定。苏萍萍等[7]对小麦种质P10078的成株期抗条锈性研究表明,其成株期抗性由1对显性主效基因控制。穆京妹等[8]研究发现,南农790成株期抗条锈性由一个显性单基因控制。曹世勤等[9]研究发现,26个甘肃省春小麦品种(系)中,19个品种(系)含有Yr1及未知抗条锈病基因,其余品种(系)推测含有未知抗条锈病基因。李邦发[10-11]研究发现,西科麦 2028、西麦6号对 CYR31 的抗病性分别由 3 对显性基因和2对显性基因、1对隐性基因控制; 对 CYR32 分别由 2 对显性和 1 对隐性基因控制、3对显性基因(其中2对表现累加作用);对 CYR33分别由1对显性基因、1对显性基因和1对隐性基因控制;对 Su11-4 由 1 对显性和 1 对隐性基因、1对显性基因和1对隐性基因重叠或独立控制,且西麦6号对条锈菌V26抗病性由1对显性基因独立控制。张莹等[12]研究发现,P9897 的成株期抗条锈性是一种数量性状,由一显一隐 2 对基因独立控制或起重叠作用控制。孙建鲁等[13]研究发现,兰天11号等的成株期抗条锈性由2对或3对基因控制。姚强等[14]对春小麦青春39的遗传分析表明,其CYR17的抗病性由1对显性基因控制。而对CYR33的抗病性由1显1隐两对基因独立控制。小麦条锈菌通过有性生殖产生强毒性菌系导致现有抗病种质资源逐渐丧失抗锈性[15],因此在育种实践中需要不断发掘新的抗条锈病资源,研究其中的抗病基因以便加以利用。本研究组前期研究发现,春小麦种质资源MY004730和ZM018243具有良好的成株期抗条锈性,本研究对以上2个春小麦种质资源的抗病基因数目及遗传规律进行分析,拟为其在抗病育种中的合理利用提供理论依据。
1 材料与方法
1.1 试验材料
供试抗条锈病春小麦种质资源MY004730、ZM018243由青海国家种质资源复份库提供,青海省农林科学院植物保护研究所保存;感病春小麦品种Taichung29由中国农业科学院植物保护研究所提供,青海省农林科学院植物保护研究所保存;以Taichung29与MY004730、ZM018243与MY004730杂交获得的F2∶3群体,由青海省农林科学院植物保护研究所保存。
1.2 试验方法
分别于2017年和2019年在青海省西宁市青海省农林科学院植物保护研究所小麦条锈病抗性鉴定自然发病圃进行抗性鉴定。
每年播种时间为3月下旬,亲本MY004730、ZM018243和 Taichung 29 先各播种1行,行长1 m,行距0.3 m,2017年将F2群体单粒播种,每行播10粒,播种30行,每个试验点1个处理,其中每20行中间加设1行 Taichung 29 作为感病对照,鉴定圃的四周各种3行 Taichung 29作诱发行。2017 年F2群体单株收获所得F3家系群体于2019年继续于同一地点测试,每个F3家系种 1 行,每个试验点1个处理,每20 行中间加设1行 Taichung 29作为感病对照,鉴定圃的四周各种3行Taichung 29作诱发行。
从7月上旬当Taichung 29充分发病的时候开始调查,对亲本MY004730、ZM018234和Taichung 29实行单行整体评估,对F2群体实行单株调查(2017 年),对F3家系群体每个家系实行单行整体评估调查(2019年),调查2次,间隔7d,记载反应型和严重度,反应型(IT)按Line等[16]0~9级的方法记录,其中0~5级为抗病,6~9级为感病,田间病害严重度(DS,%)按0、2、5、10、15、20、25、30、35、40、45、50、55、60、65、70、75、80、85、90、95、100分级记录[17],反应型以2次调查数据的平均值作为最终统计,严重度以第二次调查的数据作为最终统计。
1.3 统计分析
2 结果与分析
2.1 反应型和病害严重度正态性检验结果分析
对Taichung 29/MY004730与ZM018243/MY004730 F2群体的反应型和病害严重度的正态性检验结果如图1所示,2个杂交组合F2群体的反应型和病害严重度的正态性分布检验结果值均小于0.005,表明均不符合正态分布,初步推测种质资源MY004730和ZM018243对小麦条锈病成株期抗性均具有质量性状抗性特征。
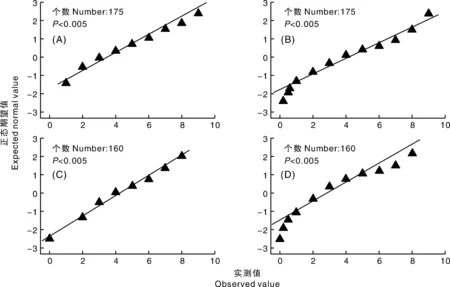
图1 Taichung 29/MY004730(A、B)与ZM018243/MY004730(C、D) F2群体反应型(A、C)和病害严重度(B、D)的正态概率图Fig.1 Normal probability of Taichung 29/MY004730 (A) and ZM018243/MY004730 (B) F2 population response type and disease severity
2.2 供试亲本的抗条锈性鉴定结果
由图2(箭头所示)可知,感病亲本Taichung 29(P1)的病害反应型均为8,病害严重度为80%,即高感。亲本MY004730反应型为2或3,即为免疫或高抗。而亲本ZM018243病害反应型为3,为高抗。
2.3 MY004730的抗条锈性遗传分析
Taichung 29与MY004730 F2:3群体的反应型和病害严重度频率分布如图所示,F2:3群体反应型和病害严重度中间型较多,反应型和病害严重度整体上均未呈现连续性分布,但不同区段内又出现连续性分布的现象。对Taichung 29与 MY004730 F2群体单株具体调查结果(表1)显示,抗病植株为130株,感病植株为45株,即Taichung 29/MY004730杂交后代F2群体卡方检验符合3R∶1S的抗感分离比(χ2=0.02,P=0.83)。对F3群体家系调查中,抗病家系131个,感病家系有44个,也符合3R∶1S的抗感分离比(χ2<0.01,P=0.97),由卡方检验结果分析得出种质资源MY004730的成株期抗条锈性由1对显性基因控制。
2.4 ZM018243与MY004730杂交 F2∶3群体的抗条锈性遗传分析
ZM018243与MY004730 F2∶3群体的反应型和病害严重度频率分布图如图2,F2∶3群体反应型和病害严重度整体均未呈连续分布,但是不同的区段内却又出现了连续性分布的现象,两年测试的结果变化趋势一致。对ZM018243与MY004730 F2∶3群体的抗条锈遗传结果见表1:在F2群体调查中,抗病植株为89株,感病植株为71株,ZM018243与MY004730 杂交后代F2群体卡方检验符合9R∶7S的抗感分离比(χ2<0.01,P=0.87)。对F3家系群体调查中,抗病家系有79个,感病家系68个,ZM018243与MY004730 F3群体卡方检验符合9R∶7S的抗感分离比(χ2=0.28,P=0.54)。即ZM018243与MY004730杂交F2∶3群体的成株期抗条锈性由2对显性基因共同作用。由MY004730成株期抗条锈性基因遗传分析结果,MY004730中有1对显性成株期抗条锈病基因,可推断出ZM018243中可能有1对显性成株期抗条锈病基因。
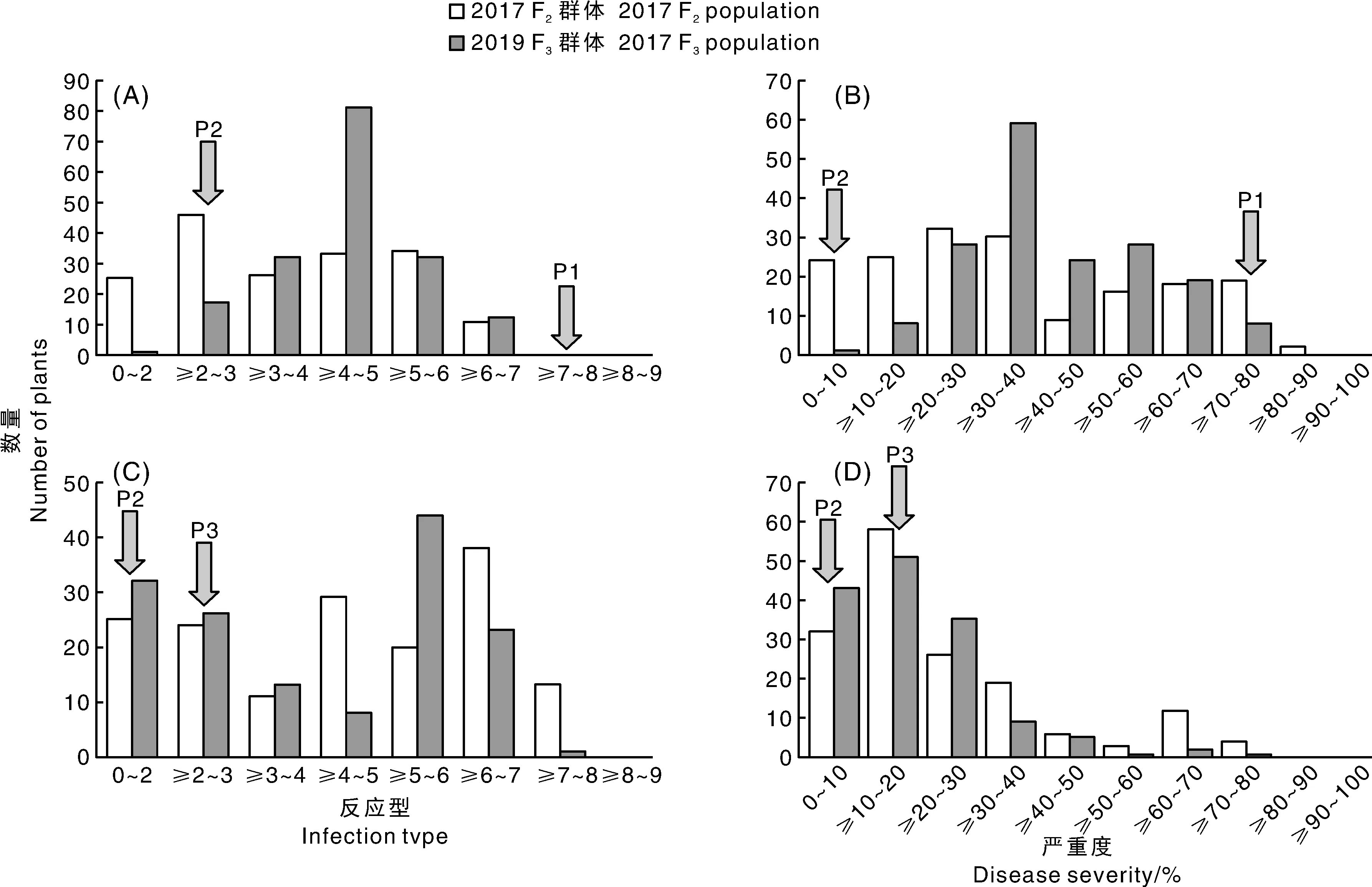
P1,Taichung29;P2,MY004730;P3,ZM018243。图2 Taichung 29/MY004730(A、B)和ZM018243/MY004730(C、D) F2∶3群体在西宁试验地所测反应型(A、C)和病害严重度(B、D)的频率分布图Fig.2 Frequency distribution of infection type and disease severity of F2∶3, populations from Taichung 29/MY004730 and ZM018243/MY004730tested in Xining
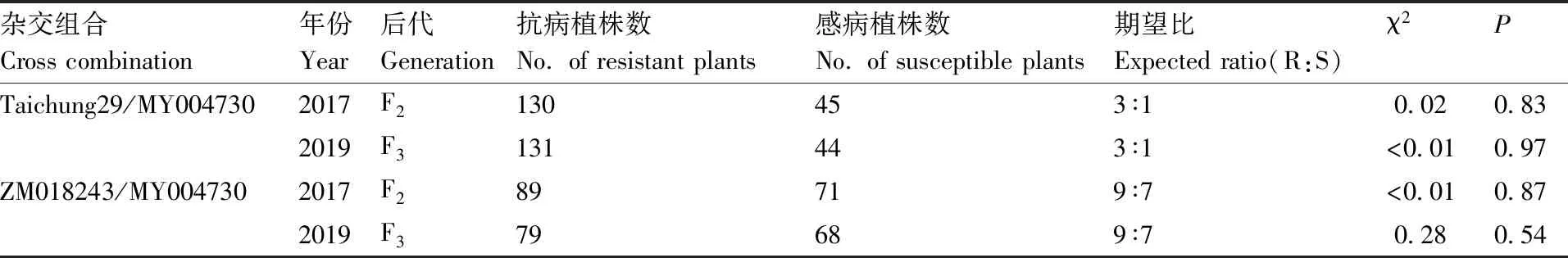
表1 Taichung 29/MY004730和ZM018243/MY004730 F2∶3群体抗病性遗传分析结果
3 结论与讨论
本研究供试的2份春小麦种质资源:ZM018243来自绵阳市农业科学研究院[19],品系名为绵阳013466-3-3-1,由绵阳11/墨西哥材料第18区//77-D301杂交选育而成;MY004730引自加拿大[19]。前期研究发现,它们均具有良好的成株期抗条锈性。本研究利用Taichung29/MY004730抗-感杂交和2个抗性亲本(抗-抗杂交)ZM018243/MY004730杂交的春小麦分别创建F2:3群体进行2个年份的大田成株期抗条锈性测试,遗传结果分析发现,MY004730中有1对显性成株期抗条锈病基因,结合2个杂交组合的遗传分析结果,推导ZM018243中也可能有1对显性成株期抗条锈病基因。本研究中,F2:3群体2个年份的试验研究结果一致,均符合孟德尔遗传学定律,且未发现群体后代的抗病性状偏向于母本的情况,存在细胞质遗传控制因素的可能性很小。因此,未再用反交群体进行测试分析。
小麦材料的抗条锈性是小麦与条锈菌在长期的协同进化中形成的一系列防御机制,是寄主植物和病原物之间相互作用所表现出来的抗病或感病现象,是二者相互作用的结果[20];对条锈病具有抗性的小麦资源中抗性基因的遗传方式有多种,单基因、多基因、主效基因与微效基因共同控制等[21],国内对小麦成株期抗条锈病基因遗传分析的报道主要是根据孟德尔遗传规律进行分析,获得1对、2对或3对抗条锈病基因的作用规律[7,10,12,22-24],本研究也是根据孟德尔遗传规律进行分析。
在对春小麦成株期抗条锈病基因遗传分析研究方面:张调喜等[25-26]应用植物数量性状主基因+多基因混合遗传模型单个分离世代分析方法对春小麦品种青春38和墨波进行了成株期抗条锈性遗传解析,推测青春38和墨波的成株期抗条锈性均为2对主基因作用;春小麦品种高原363的成株期抗条锈性也是由2对主基因作用[27]。应用植物数量性状主基因+多基因混合遗传模型的分析方法主要从主效基因和微效基因的角度来分析和推断,在数量性状的基因遗传分析中应用较多。孟德尔经典遗传规律在质量性状的基因遗传分析中应用广泛,不仅可以分析出抗病基因的数目,还能分析出抗病基因的显、隐性特性,有利于本研究中涉及到的抗病基因的推导分析,基因推导的方法省去了单独再做抗病春小麦资源ZM018243中抗病基因的遗传分析的工作量,提高了工作效率。此外,可从抗-抗杂交的春小麦群体中筛选抗病基因聚合材料有利于下一步工作的进行。
小麦抗条锈病基因的作用方式复杂多样,本研究发现,春小麦种质资源MY004730和ZM018243中可能均含有单显性成株期抗条锈病基因,更容易直接进行下一步对抗病基因的研究,不再进行基因分离和单基因系筛选的工作。根据本研究遗传分析结果,下一步可以进行抗病基因的分子标记定位,找到与抗病基因紧密连锁的标记,为分子标记辅助选择育种服务,同时还能在抗-抗杂交的春小麦群体中直接筛选抗病基因聚合材料,为育种提供优良抗病材料。
