不同抗寒性葡萄品种根系形态结构及生理性状差异分析
2021-04-25孙艳韩斌袁军伟刘长江李敏敏尹勇刚贾楠郭紫娟赵胜建
孙艳, 韩斌, 袁军伟, 刘长江, 李敏敏, 尹勇刚,贾楠, 郭紫娟, 赵胜建
(河北省农林科学院昌黎果树研究所, 河北 昌黎 066600)
葡萄(VitisviniferaL.)作为世界性的重要经济水果,因其具有适应性强、产量高、经济效益显著等优点,受到我国北方地区果农的广泛青睐,成为我国农业产业化的重要组成部分。低温冻害是影响北方地区葡萄产业发展的主要限制因素。近年来我国北方地区极端低温频发,使葡萄产业遭受巨大经济损失。因此,葡萄抗寒性研究成为研究热点,特别是探索适宜、快速评价品种抗寒性的方法对于葡萄抗寒性研究具有重要意义。
前人对葡萄抗寒性的鉴定方法及抗寒生理指标进行了研究,由于相对电导率大小与葡萄抗寒能力呈负相关[1-2],因此,相对电导率法是目前葡萄进行抗寒性鉴定较为可靠的方法之一。但电导率法预处理所需时间较长,对处理温度及时间要求严格,且试材需求量大,因此不适用于大量种质的抗寒性筛选[3]。生长试验法和组织褐变法也可用于鉴定葡萄抗寒性。李桂荣等[4]研究发现,随着胁迫温度的降低,6个葡萄品种扦插枝条的生根率均呈下降趋势;郑晓翠等[5]利用组织褐变法对14个葡萄砧木品种的抗寒性进行了分级。这两种鉴定方法均存在检测时间长的问题,且要求操作者有一定经验积累[6]。除以上3种方法外,可溶性糖、脯氨酸、丙二醛和淀粉含量等指标也可用来鉴定葡萄抗寒性[7-9]。然而生理指标的测定过程更加繁琐,且对环境条件要求较高[10]。Viands[11]和刘志鹏等[12]研究了苜蓿根部抗寒的形态机理,指出根系形态与抗寒性密切相关,分枝型苜蓿比直根型苜蓿具有更强的抗寒性,该方法直观且操作简便。然而,截至目前,关于葡萄根系形态和抗寒性关系的研究尚不多,关于葡萄根系质地硬度与抗寒性关系的研究也相对空白[13]。基于此,本研究以8个不同抗寒性葡萄品种为试验材料,探讨了葡萄休眠期的根系角度、解剖结构、含水量及质地硬度等指标与抗寒性的关系,以期筛选出操作简单高效、结果可靠的抗寒指标,进而为葡萄抗寒性的鉴定、高抗寒品种的选育及抗寒栽培提供科学依据和技术支撑。
1 材料与方法
1.1 试验材料
供试葡萄品种包括高抗寒品种:左山一(VitisamurensisRupr.)和贝达(Vitisriparia×Vitislabrusca),中抗寒品种:山河2号(Vitisamurensis×Vitisriparia)、5BB(Vitisberlandieri×Vitisriparia)、SO4(Vitisberlandieri×Vitisriparia)和1103P(Vitisberlandieri×Vitisrupestris),低抗寒品种:玫瑰香(VitisviniferaL.)和红地球(VitisviniferaL.),不同组分间抗寒性差异显著[14-15]。葡萄品种均来自于河北省农林科学院昌黎果树研究所葡萄资源圃,栽培管理条件一致。以一年生扦插苗的成熟根系为试验材料,试验于2017年11月进行。
1.2 试验设备
试验设备包括CT3质构分析仪(美国Brookfield公司)、GDJS-500L高低温交变试验箱(江苏无锡索亚特试验设备有限公司)、雷磁DDS-11A型电导率仪(上海仪电科学股份有限公司)、切片机(江苏常熟科学仪器厂)、KD-T电脑生物组织摊烤片机(浙江省金华市科迪仪器设备有限公司)、奥林巴斯BX51显微镜(日本奥林巴斯有限公司)、海尔BCD-137TMPA低温冰箱(青岛海尔股份有限公司)和2WAJ阿贝折光仪(上海光学仪器五厂)。
1.3 测定项目及方法
1.3.1相对电导率测定预处理:以一年生扦插苗根系为材料,将根剪下后用自来水冲洗以去除表面泥沙,剪去须根,选取粗度为2.0~3.0 mm的根段。用去离子水反复冲洗,晾干,剪成4.0~5.0 mm长的根段,放入10 mL离心管,加盖,置于0 ℃冰箱保存。参照前人的研究结果[16-17],以-9 ℃作为最佳冷冻处理温度,采用高低温交变试验箱进行低温处理。处理过程如下:以4 ℃·h-1的降温速度将温度降至预设温度,在预定温度下保持4 h,以4 ℃·h-1的速度升至4 ℃,然后于4 ℃的冷藏冰箱中备用。
将根段切成2.0~3.0 mm 长的小段。称取1.00 g根段置于30 mL带刻度试管,加入25 mL去离子水,加盖,置于20~23 ℃下保温24 h。保温期间用手震荡1~2次。用电导率仪测定浸出液电导率(C1),然后煮沸20 min杀死组织,静置3~4 h至室温测定电导率(C2),计算相对电导率(relative conductance,REC)。每个品种重复3次。
REC=C1/C2×100%
1.3.2根系角度的测量将葡萄根系挖出,统计骨干根(2.0 mm<直径<5.0 mm)总数。用量角器测量骨干根角度,以根系与主干延伸方向形成的夹角来衡量根系类型[18],夹角小于40°为垂直根, 夹角大于 70°为水平根, 介于两者之间为倾斜根,每个品种测量50棵植株。骨干根总数为垂直根、倾斜根和水平根三者之和。分枝角度(root angle,RA)为所有骨干根角度的平均值。垂直根、水平根和倾斜根的比例计算公式如下。
垂直根占比=垂直根条数/骨干根总数×100%
倾斜根占比=倾斜根条数/骨干根总数×100%
水平根占比=水平根条数/骨干根总数×100%
1.3.3根系解剖结构的观察将各品种葡萄根系洗净去除须根后,选取粗度为2.0~3.0 mm的根段,剪成4.0~5.0 mm长。制作石蜡切片[19],步骤如下:固定-脱水-透明-浸蜡-包埋-修蜡块和粘蜡块-切片-展片与粘片-脱蜡-染色-封片。最后,将晾干后的载玻片置于显微镜下观察、照相,并对射线细胞大小(size of ray cells,SRC)、导管直径(vessel diameter,VDR)和导管密度(vessel density,VDY)等指标进行测量。射线细胞大小为25个细胞的平均值,导管直径为20个导管的平均值,数据均在40 倍镜下观察测得,皮层比率(ratio of cortex thickness,RCT)在10倍镜下观察测得。每个品种随机选择5个根段进行处理。
1.3.4根系含水量的测定根系总含水量(total water content,TWC)、自由水含量(free water content,FWC)参照高俊凤[20]的方法进行测定。束缚水含量(bound water content,BWC)及自束比(自由水含量与束缚水含量之比,F/B)计算公式如下。
BWC=TWC-FWC
F/B=FWC/BWC
1.3.5根系硬度及做功量的测定选取粗度为(2.5±0.2) mm成熟根段,洗净将外表水分吸干,剪成长度(5.0±0.3) mm,放置于质构仪测试平板上,用刀片探头TA-CKA对根段进行压缩模式测试,横切即刀片垂直于根系延伸方向,纵切即根系延伸方向垂直于测试平板。测试参数如下:触发点负载5 g,测试速度0.7 mm·s-1,返回速度0.7 mm·s-1,数据频率10 points·s-1,根系横切目标值2.0 mm,纵切目标值1.0 mm,分别测定横切及纵切过程中的根系硬度及做功量。根系硬度包括横切硬度(cross cutting hardness,CCH)和纵切硬度(longitudinal cutting hardness,LCH),做功量包括横切总功(cross cutting total work,CCTW)和纵切总功(longitudinal cutting total work,LCTW)每个品种横切和纵切分别选取20段根系进行测定。葡萄根系质地特征曲线见图1。以图1中的最高峰F表示硬度(g),峰面积S表示做功量(mJ)。
1.4 数据统计
利用Microsoft Office Excel 2016软件进行数据处理,并制图;采用SPSS 17.0软件对差异显著性分析及Duncan 多重比较。
2 结果与分析
2.1 不同葡萄品种根系相对电导率
图2结果显示,高抗寒品种左山一和贝达的根系相对电导率值较小,分别为41.49%和42.86%,均显著低于其他品种;低抗寒品种玫瑰香和红地球的根系相对电导率值较大,分别为72.9%和75.98%,显著高于其他品种;中抗寒品种山河2号、5BB、SO4和1103P的根系相对电导率在45%~55%,显著高于高抗寒品种,但低于低抗寒品种。
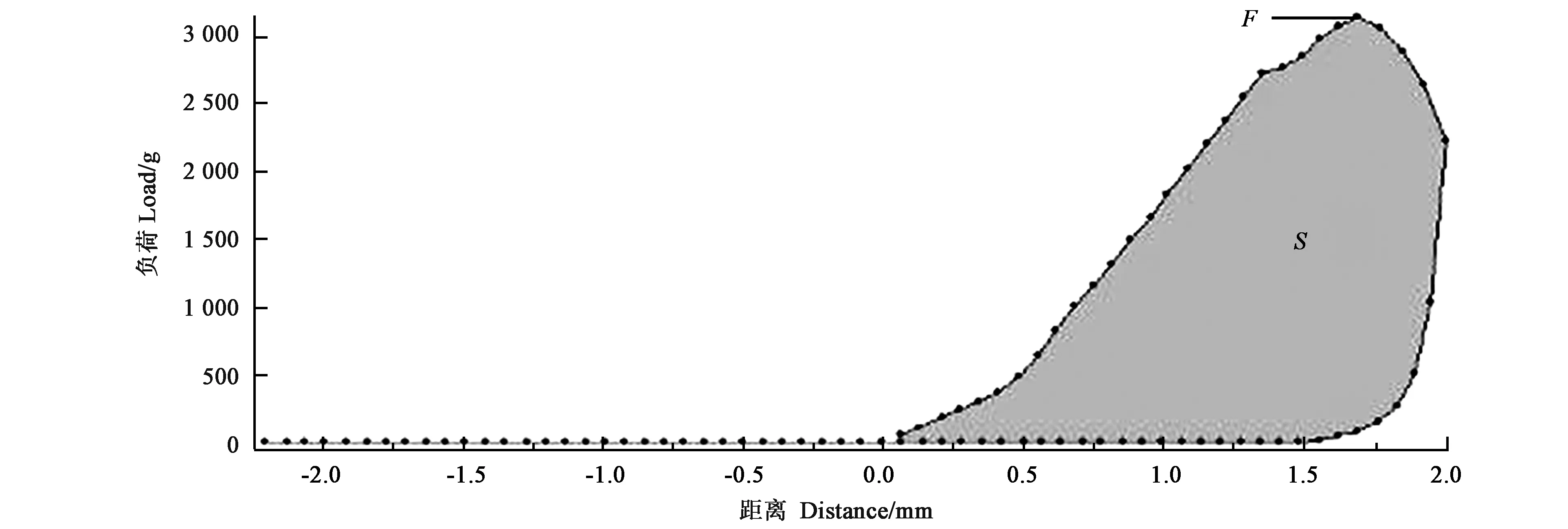
图1 葡萄根系压缩模式下质地特征曲线Fig.1 Texture characteristic curve in grapevine root under compression mode
2.2 不同葡萄品种根系角度
从表1可知,高抗寒品种(左山一和贝达)的分枝角度与低抗寒品种(玫瑰香和红地球)间差异不显著(P>0.05)。其中,低抗寒品种红地球的垂直根占比较高,为50%;倾斜根占比较低,为38.89%。中抗寒品种5BB的垂直根占比较低(7.69%),水平根占比较高(11.54%);品种SO4的倾斜根占比较高(85.00%);品种1103P的根系中未检测到水平根。由此可见,不同葡萄品种的根系角度分布与抗寒性未呈现规律性。
2.3 不同葡萄品种根系解剖结构
葡萄根系解剖结构与其抗寒性有相关关系[21]。从表2结果可知,不同抗寒性葡萄品种的根系组织结构差异较大。高抗寒品种左山一的射线细胞大小、导管密度及导管直径均最小;贝达的皮层比率最小。低抗寒品种(玫瑰香和红地球)的射线细胞大小、皮层比率、导管密度及导管直径均显著高于高抗寒品种(左山一和贝达)。显微观察不同品种的根系解剖结构(图3),高抗寒品种(左山一和贝达)的射线细胞较小、数目较多,组织结构紧密,形成了清晰的髓射线,导管细且数目较少,且在木质部边缘分布较少;而低抗寒品种(玫瑰香和红地球)的射线细胞较大且数目较少,组织结构疏松,导管粗且数目较多,分布可达到木质部边缘。由此表明,根系的内部结构特征与葡萄抗寒性密切相关。
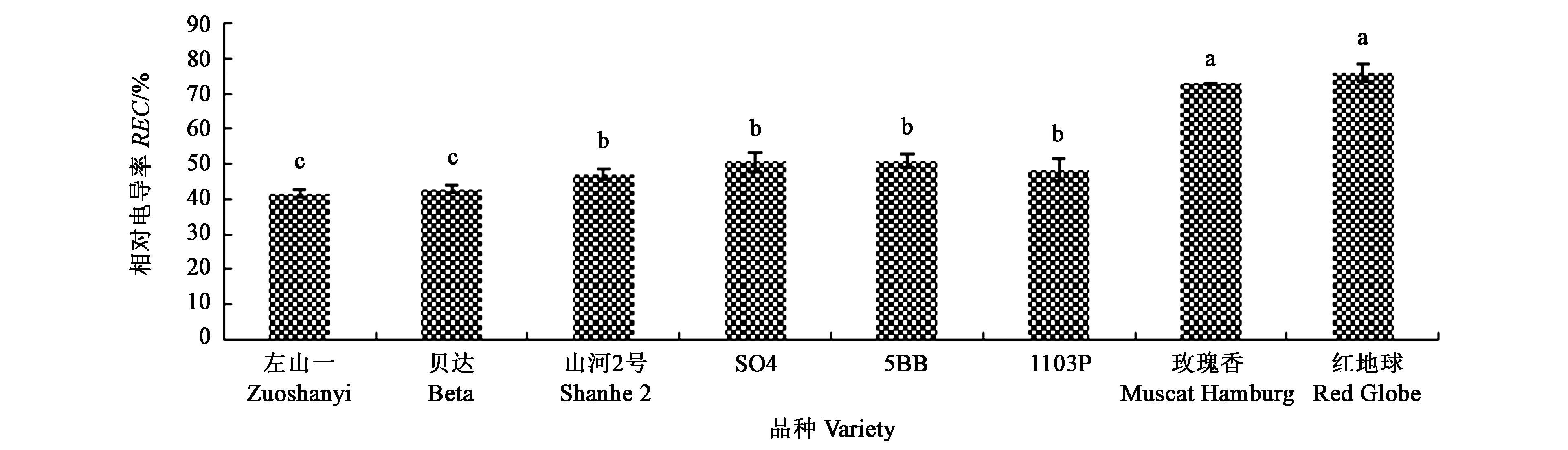
注:不同小写字母表示在P<0.05水平差异显著。Note:Different small letters indicate significant difference at P<0.05 level.图2 葡萄根系的相对电导率Fig.2 Relative conductivity of the grapevines roots
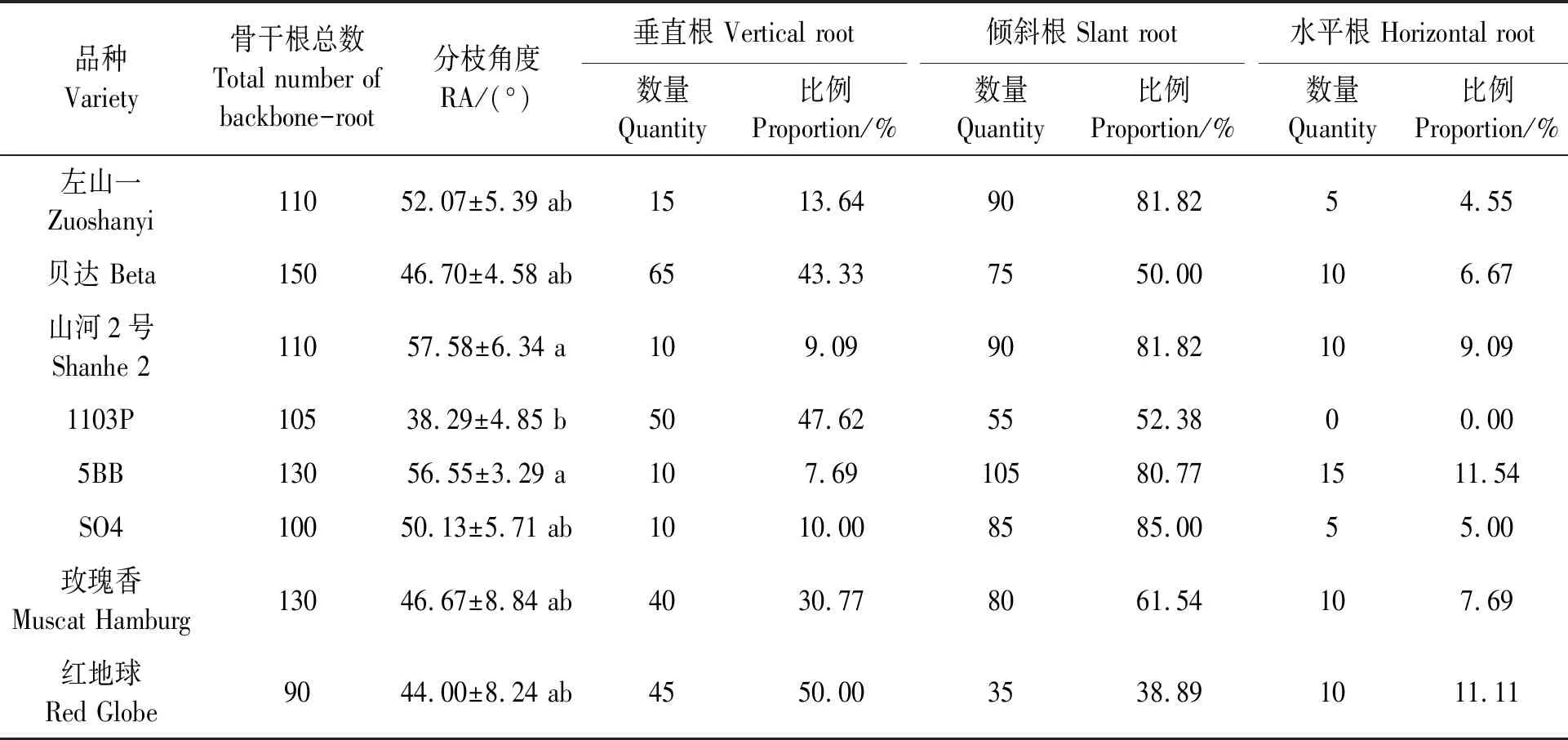
表1 葡萄根系的类型Table 1 Root types of the grapevines

表2 葡萄根系解剖结构特征Table 2 Anatomy characteristics of the grapevines roots

图3 葡萄根系解剖结构Fig.3 Anatomical structure of the grapevines roots
2.4 不同葡萄品种根系含水量
从表3可知,不同抗寒性品种间的总含水量及自由水含量差异显著。高抗寒品种(左山一和贝达)的自由水含量和自束比显著低于其他品种,左山一的总含水量显著低于其他品种。低抗寒品种(玫瑰香和红地球)的总含水量、自由水含量和自束比均显著高于其他品种。
2.5 不同葡萄品种根系硬度及做功量
根系硬度和做功量代表根系在外力的切割作用下抵抗断裂的能力。不同品种根系的硬度和做功量均存在较大差异(表4)。其中,高抗寒品种左山一的根系横切硬度、横切总功、纵切硬度和纵切总功均最高,且后三项指标显著高于其他品种;低抗寒品种红地球的根系横切硬度、横切总功、纵切硬度和纵切总功均最低,且后两项指标显著低于其他品种。
2.6 葡萄根系生理指标相关性分析
对葡萄根系14个抗寒指标进行相关性分析,结果(表5)表明,根系电解质渗出率与除根系角度外的12个性状均呈极显著相关;射线细胞大小与横切总功、横切硬度、纵切总功和纵切硬度间均呈极显著负相关;总含水量与横切总功、横切硬度、纵切总功和纵切硬度间均呈极显著负相关。

表3 葡萄根系含水量Table 3 Water content of the grapevines roots
3 讨论
3.1 葡萄根系相对电导率与抗寒性的关系
植物细胞电解质的外渗程度可用相对电导率来表示,反映植物细胞膜系统的低温伤害程度[22]。相对电导率越大,表明植物组织细胞电解质透过细胞膜的通透性越大,即植物抗寒性越弱[23]。葡萄根系的相对电导率能够真实反映不同品种的抗寒性差异[24]。本研究中,低温胁迫下,左山一和贝达的抗寒性较强,其根系的相对电导率显著低于其他品种;而红地球和玫瑰香的抗寒性较弱,其根系的相对电导率显著高于其他品种,这与高振等[15]研究结果一致。
3.2 葡萄根系角度与抗寒性的关系
植物根系分布影响着植物对逆境的抵抗能力[25]。目前,有关植物根系形态与抗寒性关系的研究主要集中于草本作物。刘志鹏等[12]研究指出,苜蓿根系形态与抗寒性密切相关,不同抗寒性品种的根系类型差异显著,分枝型根系比直根系能更好地忍受冻害。孙波等[26]研究发现,高抗寒甘蔗品种根系的分枝角度小,呈向下生长趋势;而低抗寒品种根系的分枝角度大,呈平周生长趋势。本研究中8个葡萄品种的根系角度与抗寒性间未呈现相关性,低抗寒品种红地球的垂直根占比为50%,高抗寒品种左山一的垂直根占比为13.64%。这与前人研究结果存在差异,可能与材料处理时期有关。葡萄根系的形态特征受到一系列对根原基进行负调节的基因型控制[27],不同品种可能存在不同的抗寒基因,这些抗寒基因只有在特定时期、特定条件下被激活,从而诱发根系形成不同的形态结构以抵御逆境[28],而这种适应性变化存在明显的基因型差异[29]。本研究中葡萄根系由于低温进入休眠,而不同品种根系对温度变化的敏感程度不同,左山一和贝达根系感受到的低温环境可能不足以诱发控制根系形态的抗寒基因表达,从而导致其垂直根比例较低。除此之外,根系形态建成也受地上部生长发育的影响。潘晓华等[30]研究指出,根冠的生长方向受地上部向根系输送碳水化合物的影响,地上部光合强度高,则垂直根和倾斜根的比例增加,反之则水平根的比例增加。由于不同葡萄品种的地上部光合特性存在差异,造成本研究中中抗寒品种1103P和SO4垂直根和倾斜根的比例较大,水平根比例较小;而高抗寒品种左山一和贝达垂直根和倾斜根的比例较小,水平根比例较大。由此可见,不同抗寒性葡萄品种的根系形态建成机理仍需深入研究。
3.3 葡萄根系解剖结构与抗寒性的关系
葡萄根系内部结构的形成是对环境长期适应的结果,与品种抗寒性密切相关。本研究发现,高抗寒品种左山一和贝达的根系具有组织紧密、射线细胞小、皮层比例低、导管小且密度低的结构特点,与低抗寒品种红地球和玫瑰香之间差异显著。因此,根系解剖结构特征可作为葡萄品种抗寒性鉴定的指标之一。这与郭修武等[31]的研究结果相一致。射线细胞体积小、排列紧密,不仅有利于水分经较短距离迁出细胞、减少细胞内结冰[32],还可以贮藏大量营养物质,增强对冻害的防御功能。木质部导管直径小,则树液结冰造成的木质部栓塞化程度低[33],使得木质部能够保持一定的水分疏导功能,从而减轻胞外结冰带来的伤害,有助于葡萄植株在受冻后的解冻过程中快速恢复疏导能力[34],从而恢复正常生理代谢。因此,导管直径小这一结构特性对于植物抗寒尤为重要。
3.4 葡萄根系含水量与抗寒性的关系
植物组织内的水分主要以自由水和束缚水两种状态存在。自由水不与原生质体紧密结合,主要存在于细胞间隙,可以自由移动;束缚水则相反[35],束缚水是细胞内亲水性大分子物质强行结合的水,对维持原生质体胶体稳定具有重要作用。葡萄根系含水量特别是自由水含量低,细胞液浓度高,生理代谢活动弱,有利于增强树体抵御冻害的能力。本研究证实,不同抗寒性葡萄品种在同一休眠时期的根系含水量存在差异,高抗寒品种左山一和贝达表现为根系总含水量和自由水含量较低、自束比较小,与低抗寒品种玫瑰香和红地球间差异显著。Lyons[36]研究指出,冻害发生时,细胞间隙中的水首先结冰形成冰核,随着低温的延续,结冰范围逐渐扩大,原生质内的水向细胞间隙渗透,造成细胞严重脱水,细胞膜皱缩,同时结冰又会对细胞膜产生挤压,造成细胞膜机械损伤。冬季当气温降至0 ℃以下时,细胞间隙的自由水便开始结冰;而束缚水的冰点在-20~-25 ℃以下[37],远低于自由水。因此,当根系中束缚水含量较高、自由水含量较低时,结冰温度相应降低,则抗寒性增强。相反,当自由水含量较高、束缚水含量较低时,葡萄根系生理代谢活动比较旺盛,则抗寒性较弱。陈佰鸿等[38]的研究也证明了根系的含水量与其抗寒性关系密切。
3.5 葡萄根系硬度与抗寒性的关系
质构仪作为一种新型的食品品质测试仪器,被用于测定经碳酸钙真空浸渍后巨峰葡萄的果实硬度[39]。Gwanpua等[40]使用质构仪研究了贮藏期间乔纳金苹果果实硬度的变化。本研究使用质构仪测定葡萄不同抗寒性品种的根系硬度,结果表明,高抗寒品种左山一和贝达的根系硬度及做功量较大,而低抗寒品种玫瑰香和红地球的根系硬度及做功量较小。同时相关分析表明,根系硬度及做功量与射线细胞大小呈极显著负相关,即射线细胞越小,组织排列越紧密,则根系硬度及做功量越大;反之,则根系硬度及做功量越小。根系木质部细胞细胞壁较厚,为根系提供了主要的结构支撑。纤维素、半纤维素、木质素和果胶质作为细胞壁的主要组成成分,关系着植物细胞机械组织的发达程度[41],进而影响着根系硬度。前人研究指出,细胞壁中纤维素、半纤维素及果胶质的含量变化与果实软化密切相关[42]。细胞壁中较多的纤维素和木质素含量能够增加种皮的机械强度和坚硬度[43]。牢固的纤维素结构使细胞壁具有高强度性能,木质素使细胞相连,增加细胞壁的抗压强度,强化植物组织,半纤维素与纤维素微纤丝结合,而果胶质则分布在半纤维素和纤维素微丝中并与之紧密交联,形成复杂的网络结构,对细胞壁的结构起到加固作用[44]。本研究中,根系解剖结果表明,高抗寒品种左山一和贝达的射线细胞较小,排列紧密,纤维素和木质素等细胞壁组成物质相对含量增加[45],因此硬度较低、抗寒性较强。且根系硬度及做功量与总含水量呈极显著负相关,即总含水量越少,根系硬度及做功量越大;反之,则根系硬度及做功量越小。前人研究表明,冬季根系水分含量低,但根系的抗拉强度却较夏季大[46]。郭凯力等[47]和蒋坤云等[48]的研究表明,根系含水量越高,细胞壁纤维韧性越低,遇到外力作用时抵抗断裂的能力也越弱,与本研究结论相似。由此表明,根系的内部结构特征与抗寒性密切相关,是植物对环境长期适应的结果。因此,根系硬度及做功量可作为葡萄抗寒性鉴定的指标之一。
综上所述,高抗寒葡萄品种具有低温胁迫下相对电导率低、根系组织结构紧密、自由水含量低、自束比小、硬度及做功量大等特点,利用这些特点可以有效评价葡萄抗寒性。根系横切总功与相对电导率间相关性较强,且测定方法简单,对环境条件要求低,适宜作为快速、准确鉴定葡萄抗寒性的指标。
