产蛋后期农大3号蛋鸡和海兰褐蛋鸡消化生理研究
2021-04-21郭晓瑞王友利周华金袁建敏
郭晓瑞,武 威,陈 静,王友利,周华金,袁建敏
(中国农业大学动物科学技术学院,北京 100193)
延长蛋鸡的饲养周期,提高经济效益是营养学研究的热点。蛋鸡进入产蛋后期,由于体内新陈代谢的变化导致其生殖与代谢机能逐渐衰退,肠道结构与功能受到破坏,消化酶数量和活性下降,从而降低机体对营养物质的吸收利用能力,最终影响生产性能[1]。马宁[2]研究发现,育成期农大3 号蛋鸡和海兰褐蛋鸡各肠段相对重量、肠道形态结构与50 周龄成年阶段的趋势不同。马丹丹[3]研究发现,随着周龄增加,太行鸡各段消化道相对长度逐渐降低,且肠道形态结构发生变化,但消化酶活性基本保持不变。王淑红等[4]研究发现,产蛋后期蛋鸡体重显著影响产蛋量,但对蛋重影响不显著,体重越大则产蛋数越低。本研究前期发现,产蛋高峰期农大3号蛋鸡消化、吸收机能和养分代谢率显著高于海兰褐蛋鸡;如不考虑品种因素,产蛋高峰期体重偏轻组蛋鸡消化、吸收机能显著高于体重正常组[5]。但产蛋后期不同体重农大3 号和海兰褐蛋鸡消化生理功能是否仍然存在差异;产蛋后期蛋鸡脂肪积累,超重现象严重,产蛋后期蛋鸡体重对消化生理指标有何影响等问题仍待探究。
因此,本实验旨在比较产蛋后期不同体重农大3 号蛋鸡和海兰褐蛋鸡消化生理的差异,为制定农大3 号蛋鸡适宜的营养标准和体重标准提供科学依据和理论指导。
1 材料与方法
1.1 实验设计与饲养管理 试验采用2×2 因子设计,采用76 周龄体重偏大(平均体重/标准体重为1.2)、体重正常(平均体重/ 标准体重为1.0)健康的商品代农大3 号蛋鸡与海兰褐蛋鸡共216 只,分为4 个处理,每处理6 个重复,每重复9 只蛋鸡,体重分组见表1。采用玉米-豆粕型基础日粮,其组成及营养成分见表2。代谢日粮在基础饲粮中添加0.5% TiO2,充分混匀,冷压制粒。试验于中国农业大学涿州养殖基地进行,鸡只采用舍内笼养方式饲养,每个鸡笼3 只蛋鸡,自由采食和饮水。每日人工光照加自然补光16 h,黑暗8 h。鸡舍配备湿帘风机,纵向负压通风降温。每日07:00 以及14:00 饲喂,15:00 收集鸡蛋。按正常程序免疫和鸡舍消毒。

表1 产蛋后期蛋鸡分组
1.2 产蛋性能测定与代谢试验 每天以重复为单位记录蛋鸡产蛋数和只数,计算各组蛋鸡产蛋率、平均蛋重;每周测定各组蛋鸡耗料量,计算平均日采食量、日均产蛋量和料蛋比(日采食量/日均产蛋量)。统计周期12 d。
采用指示剂法,预试期12 d,饲喂相同粉状基础料;正试期3 d,饲喂相同颗粒状代谢料,最后一天收集8 h内新鲜粪样。挑拣出粪盘中羽毛、饲料,混匀喷洒10% 盐酸溶液固氮,置于-20℃冰箱保存。通过检测饲料和粪样中能值、粗蛋白质、粗脂肪和指示剂含量计算蛋鸡表观代谢能和粗蛋白质、粗脂肪表观代谢率。计算公式:
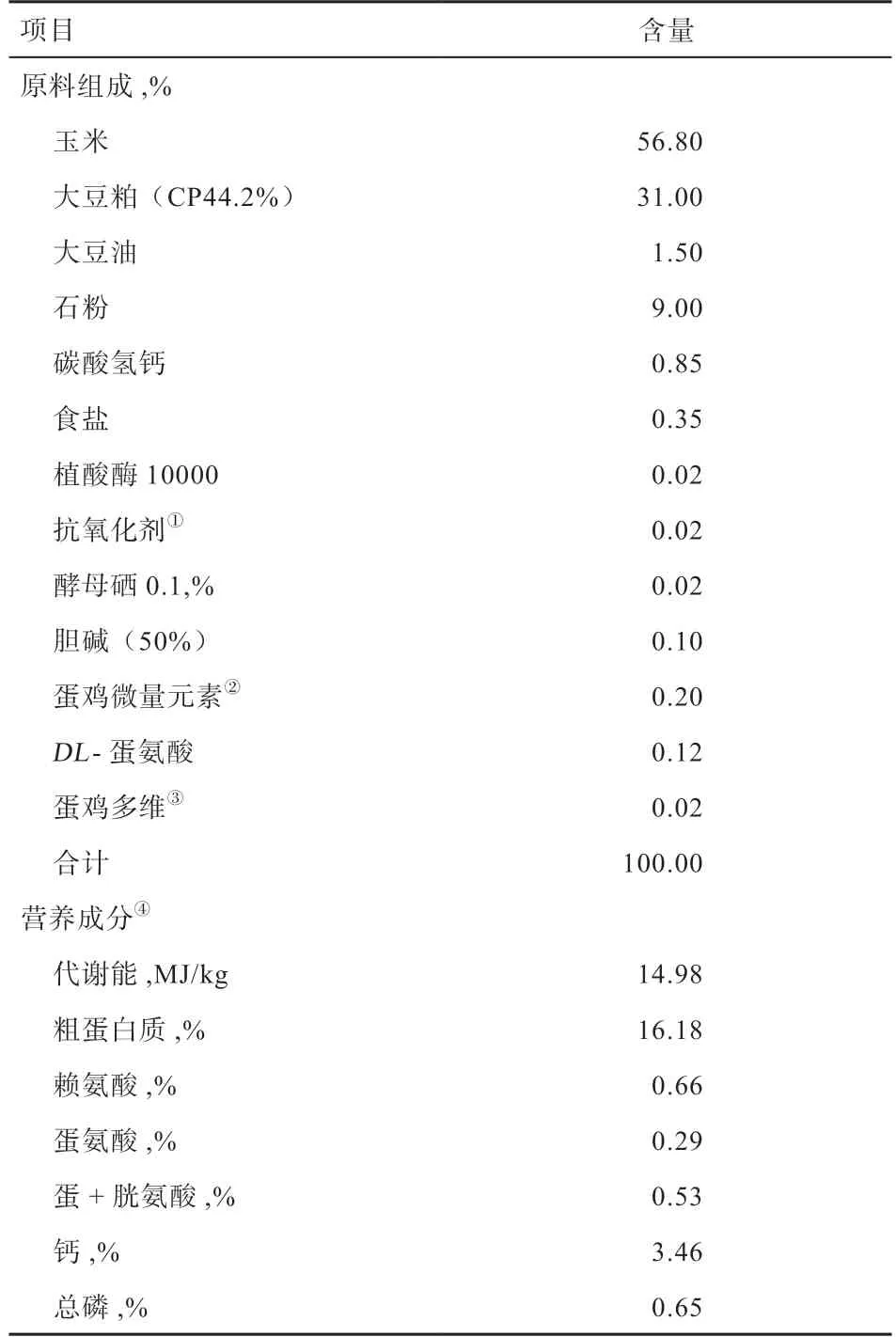
表2 基础日粮组成和营养成分

1.3 血液生化指标测定 粪便收集完毕,在蛋鸡自由采食状态下,每个处理随机选取6 只蛋鸡进行翅下静脉采血。血液经3 000 r/min 离心10 min 制备血清。采用德国Prodia Diagnostics 试剂盒使用日立7600 全自动生化检测仪(日本日立公司)检测血清丙氨酸氨基转移酶、天门冬氨酸氨基转移酶、总胆固醇、甘油三酯、尿酸、葡萄糖含量;采用南京建成生物工程研究所总胆汁酸试剂盒(E003-2)使用Multiskan MK3 酶标仪(Thermo Fisher Scientific 公司)检测血清胆汁酸含量;采用天津九鼎医学生物工程有限公司试剂盒,使用GC—2010型免疫计算器(安徽中科中佳科学仪器有限公司)检测三碘甲状腺原氨酸(T3)和四碘甲状腺原氨酸(T4)含量。
1.4 器官相对重量测定 采血结束后,每个处理每个重复选取1 只蛋鸡麻醉致死,屠宰取样。迅速分离腺胃、肌胃、肝脏、胰腺、输卵管、卵巢(带卵泡)。称量腺胃、肌胃(去除食糜)、肝脏、胰腺、输卵管、卵巢(带卵泡)重量,用于器官相对重量的计算。

1.5 空肠消化酶活性测定 取空肠食糜、黏膜,糜蛋白酶(试剂盒编号:No.A080-3)、淀粉酶(试剂盒编号:No.C016-1)、脂肪酶(试剂盒编号:No.A054-2)以及空肠黏膜麦芽糖酶(试剂盒编号:No.A082-3)、蔗糖酶(试剂盒编号:No.A082 -2)活性均采用南京建成生物工程研究所试剂盒进行检测,具体操作参考南京建成生物工程研究所试剂盒相关说明。胰腺、空肠食糜胰蛋白酶(试剂盒编号:BC2315)活性采用Solarbio 试剂盒(北京索莱宝科技有限公司)检测,检测方法参考试剂盒说明书。
1.6 营养素转运载体测定 取空肠分子样于-80℃冰箱保存,用于测定脂肪酸转运载体表达量。空肠总RNA 的提取、反转录以及荧光定量PCR 操作方法参考Wang 等[6]的方法。目的基因以及内参基因引物序列见表3,基因丰度表达的结果采用2-ΔΔCT进行分析比较。

表3 实时荧光定量PCR 引物序列
1.7 统计分析 所有数据均通过Grubbs 法进行异常值剔除,采用SPSS 21.0 的一般线性模型(GLM)进行双因子方差分析,互作效应差异显著的数据采用Duncan's多重比较检验,以P<0.05 作为差异显著标准。
2 结果
2.1 产蛋后期不同品种、体重蛋鸡生产性能比较 蛋鸡品种、体重对其生产性能均无显著互作效应(P>0.05)。农大3 号蛋鸡产蛋率、总蛋重、平均蛋重、日均产蛋量、日均采食量均显著低于海兰褐蛋鸡。不考虑品种因素,体重正常组蛋鸡产蛋率、总蛋重、日均产蛋量均显著高于体重偏大组,但料蛋比显著低于体重偏大组,其余指标无显著差异(图1)。
2.2 产蛋后期不同品种、体重蛋鸡器官相对重量和黄卵泡数比较 由图2 可知,产蛋后期蛋鸡品种、体重对肝脏相对重量具有显著互作效应,海兰褐-体重正常<海兰褐-体重偏大<农大3 号-体重偏大<农大3 号-体重正常,海兰褐蛋鸡不同体重差异不显著。农大3 号蛋鸡与海兰褐蛋鸡相比,肌胃、肝脏、卵巢相对重量显著提高。其余器官指数差异不显著。蛋鸡品种、体重对大黄卵泡数存在显著互作效应,海兰褐-体重正常<农大3号-体重偏大<农大3 号-体重正常<海兰褐-体重偏大,农大3 号蛋鸡不同体重大黄卵泡数差异不显著。不考虑品种因素,体重偏大组蛋鸡大黄卵泡数显著高于体重正常组。品种、体重以及二者的交互作用对蛋鸡小黄卵泡数均无显著影响。
2.3 产蛋后期不同品种、体重蛋鸡养分代谢率比较 由图3 可知,产蛋后期蛋鸡品种、体重对饲料粗脂肪表观代谢率具有显著互作效应,农大3 号-体重正常<农大3 号-体重偏大<海兰褐-体重正常<海兰褐-体重偏大(P<0.05)。农大3 号蛋鸡与海兰褐蛋鸡相比,饲料粗脂肪表观代谢率极显著降低;不考虑品种因素,体重偏大组蛋鸡粗脂肪表观代谢率极显著高于体重正常组(P<0.001)。蛋鸡品种、体重以及二者的互作效应均对饲料表观代谢能以及粗蛋白质表观代谢率无显著影响。
2.4 产蛋后期不同品种、体重蛋鸡空肠消化酶活性比较由表4 可知,产蛋后期蛋鸡品种、体重以及二者的交互作用对空肠食糜、黏膜消化酶活性均无显著影响。

图1 不同体重农大3 号蛋鸡和海兰褐蛋鸡生产性能比较

图2 不同体重农大3 号蛋鸡和海兰褐蛋鸡器官相对重量比较
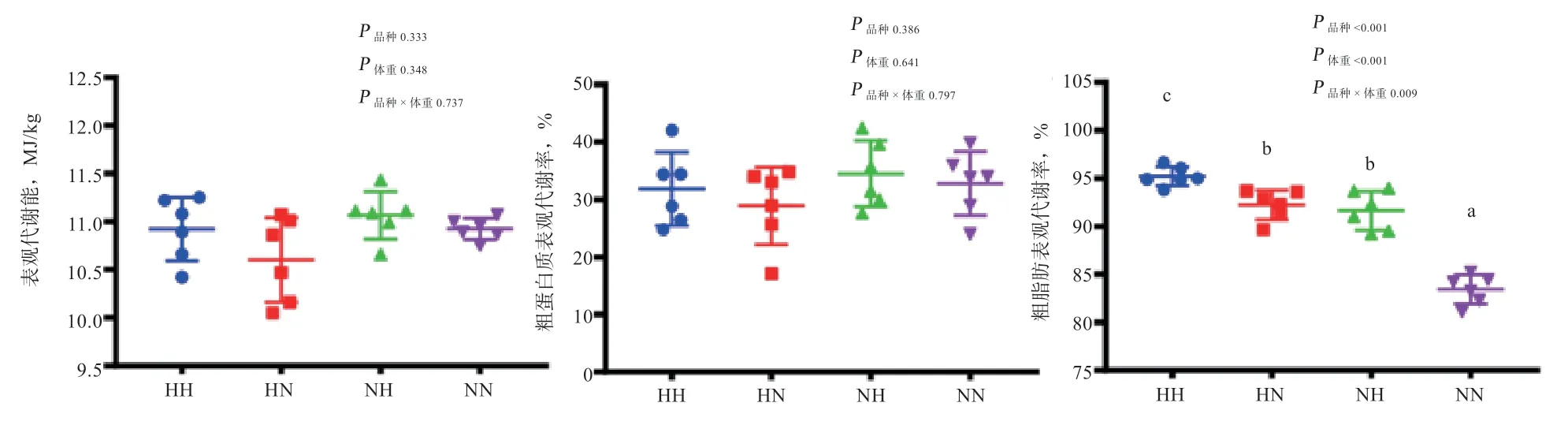
图3 不同体重农大3 号和海兰褐蛋鸡养分代谢率比较

表4 不同体重农大3 号蛋鸡和海兰褐蛋鸡空肠消化酶活性比较
2.5 产蛋后期不同品种、体重蛋鸡空肠脂肪酸转运载体mRNA 相对表达量比较 产蛋后期蛋鸡品种、体重对空肠脂肪酸合成关键酶乙酰辅酶a 羧化酶(ACC)和脂肪酸合酶(FASN)mRNA 相对表达量具有显著互作效应,海兰褐-体重正常<农大3 号-体重偏大<农大3 号-体重正常<海兰褐-体重偏大,FASNmRNA 相对表达量前两者趋势相反,农大3 号蛋鸡不同体重之间2种酶基因mRNA 相对表达量差异不显著。农大3 号蛋鸡脂肪酸转运体FAT/CD36mRNA 相对表达量显著高于海兰褐蛋鸡,而FASNmRNA 相对表达量显著低于海兰褐蛋鸡。不考虑品种因素,体重偏大组蛋鸡FASNmRNA 相对表达量有高于体重正常组的趋势,但差异不显著(表5)。

表5 不同体重农大3 号蛋鸡和海兰褐蛋鸡脂肪酸转运载体相对表达量
2.7 产蛋后期不同品种、体重蛋鸡血液生化指标比较 由图4-a 和图4-b 可知,产蛋后期蛋鸡品种、体重仅对血清丙氨酸氨基转移酶活性具有显著互作效应,农大3 号-体重正常<海兰褐-体重正常<海兰褐-体重偏大<农大3号-体重偏大,海兰褐蛋鸡不同体重血清丙氨酸氨基转移酶活性差异不显著。农大3 号蛋鸡与海兰褐蛋鸡相比,血清丙氨酸氨基转移酶活性、尿酸含量显著升高;不考虑品种因素,体重正常组蛋鸡血清丙氨酸氨基转移酶活性、甘油三酯、胆固醇、胆汁酸、尿酸含量显著低于体重偏大组。其余指标差异不显著。蛋鸡品种、体重以及二者的互作效应对血清T3和T4含量均无显著影响(图4-c)。
3 讨 论
3.1 产蛋后期不同品种、体重蛋鸡生产性能比较 本研究中,产蛋后期蛋鸡体重普遍较大,这可能与产蛋后期蛋鸡脂肪沉积增加有关。研究发现,蛋鸡体重与蛋重呈正相关关系,但与产蛋率呈负相关关系[7]。本研究发现,产蛋后期不同体重蛋鸡蛋重差异不显著,但体重正常组蛋鸡产蛋率显著高于体重偏大组。这一结果与王淑红等[4]研究结果类似,即产蛋后期蛋鸡体重对产蛋量影响显著,但对蛋重影响不显著,体重越大则产蛋数越低。这可能是由于产蛋后期蛋鸡体重偏轻,脂肪沉积少,更有利于产蛋。此外,产蛋后期农大3 号蛋鸡生产性能较产蛋高峰期而言,其劣势更加明显。Merat 等[8]研究发现,矮小型基因DW 对蛋鸡后期产蛋率的不良影响要高于前期。乔红光[9]的研究结果也同样证实,在相同的饲养条件下,矮小型农大3 号蛋鸡72 周龄产蛋数比普通蛋鸡要低。研究认为,当蛋鸡产蛋率下降至75% 左右,同时卵巢内的初级卵泡、次级卵泡和等级卵泡数量显著减少,而卵泡闭锁率显著增加表明家禽卵巢机能快速衰退[10]。本研究中,农大3 号蛋鸡卵巢相对重量较海兰褐蛋鸡有显著提高,但其大黄卵泡数显著低于海兰褐-体重偏大组;同时农大3 号蛋鸡产蛋率显著低于海兰褐蛋鸡,仅在75% 左右,这说明农大3 号蛋鸡生产性能骤降可能与其产蛋后期卵巢衰退速度较快有关。
3.2 产蛋后期不同品种、体重蛋鸡消化器官相对重量比较 本研究中,相较于海兰褐蛋鸡,农大3 号蛋鸡的肌胃、肝脏相对重量显著提高。宁中华等[11]和周大薇[12]的研究也发现,矮小鸡肌胃、腺胃、十二指肠相对重量显著高于正常鸡,消化系统较发达。许多学者研究发现,家禽的消化器官重量,尤其是肝脏、肌胃的相对重量与养分代谢率呈显著正相关关系[13-14]。杨瑛[15]研究发现,矮小型黄羽肉种公鸡对棉粕的总氨基酸消化率显著高于正常型肉种公鸡。这说明矮小型蛋鸡可能是通过促进消化器官的发育进而提高饲料养分代谢率。

图4 不同体重农大3 号蛋鸡和海兰褐蛋鸡血液生化指标比较
3.3 产蛋后期不同品种、体重蛋鸡消化、吸收、代谢功能比较 研究发现,动物衰老的同时伴随着小肠肠道形态、细胞增殖速率等的改变,导致对营养物质的吸收能力降低[16]。本研究中,产蛋后期农大3 号蛋鸡消化酶活性以及饲料表观代谢能、粗蛋白质表观代谢率与海兰褐蛋鸡差异不显著,这可能是由于产蛋后期蛋鸡消化代谢功能衰退。此外,本研究还发现,产蛋后期海兰褐蛋鸡粗脂肪表观代谢率显著高于农大3 号蛋鸡,以体重偏大组最高,显著高于其他3 组。但产蛋后期农大3 号蛋鸡与海兰褐蛋鸡饲料表观代谢能以及粗蛋白质表观代谢率无显著差异。由此推断,产蛋后期农大3 号蛋鸡日粮代谢能水平可以参考海兰褐蛋鸡营养标准,或略低于海兰褐蛋鸡。饲粮中的脂肪主要以三酰甘油的形式存在,被胃、胰腺消化酶水解为甘油一脂和脂肪酸,由脂肪酸转运载体转运吸收进入血液,随血液运输至各个组织器官,参与机体的合成代谢。此外,脂肪酸还可通过ACC和FASN等关键酶从头合成,也可通过β氧化分解供能。本研究中,海兰褐-体重偏大组这2 种酶(ACC、FASN)基因相对表达量显著高于其他3 组,与粗脂肪表观代谢率的结果一致。研究表明,FASN、ACC基因表达量的增加会引起内源脂肪酸含量增加,导致脂肪的沉积[17]。这说明产蛋后期海兰褐体重偏大组蛋鸡通过改变脂肪酸从头合成的关键酶FASN、ACCmRNA 相对表达量,提高脂肪酸的合成效率,造成脂肪沉积量增加,进而改变粗脂肪表观代谢率。
3.4 产蛋后期不同品种、体重蛋鸡血液生化指标比较丙氨酸氨基转移酶和天门冬氨酸氨基转移酶是催化脱氨基作用的关键酶,对肝脏的蛋白质代谢起着枢纽的作用[18]。尿素氮是蛋白质分解代谢的最终产物,血清尿素氮水平可以较准确反映动物体内蛋白质的代谢情况和日粮氨基酸的平衡情况[19-21]。本研究中,农大3 号蛋鸡与海兰褐蛋鸡相比,血清丙氨酸氨基转移酶活性、尿酸含量显著提高。血清丙氨酸氨基转移酶、尿酸的含量与蛋白的分解代谢相关,这说明农大3 号蛋鸡对饲粮蛋白质的分解代谢速率高于海兰褐蛋鸡。体重正常组蛋鸡相较于体重偏大组而言,血清甘油三酯、胆固醇、胆汁酸含量显著降低。血清甘油三酯和胆固醇含量的升高导致脂肪沉积量以及体重增加。这说明体重偏大组蛋鸡的脂肪沉积量高于体重偏轻组。胆汁酸主要是通过促进饲粮中脂肪的消化吸收并减少脂肪酸的分解,进而提高血清甘油三酯和胆固醇的含量。研究发现,血清中的总胆汁酸含量与甘油三酯及胆固醇的含量呈正相关,与前期研究结果一致[22]。
研究发现,矮小鸡由于较低的肝脏脱碘酶活性,抑制T4向T3转化,导致血液中T3循环水平下降,而T4水平维持正常或略有升高[23]。而本研究中,农大3 号蛋鸡T3和T4含量与海兰褐蛋鸡无显著差异。赵东伟等[24]研究发现,矮小型鸡血清T3、T4含量与正常型鸡未存在明显差异,与本研究结果一致。乔红光[9]研究同样发现,6 周龄和10 周龄普通蛋鸡血清T3含量显著高于矮小鸡,其余周龄显著低于矮小鸡。这说明检测周龄以及品种选择会影响血清中T3和T4含量。
4 结 论
本研究结果显示,产蛋后期农大3 号蛋鸡脂肪酸转运能力降低,造成其消化吸收功能以及粗脂肪表观代谢率显著低于海兰褐蛋鸡。因而,在配制产蛋后期农大3号蛋鸡日粮时,其代谢能需要量应略低于海兰褐蛋鸡。产蛋后期体重正常组蛋鸡可能通过降低对脂肪的合成代谢,减少脂肪沉积来提高生产性能。
