Elastin 基因在小鼠早期妊娠过程中的表达
2021-04-21王一妹王胜轩郭加璇李世杰马兴红
王 悦,王一妹,王胜轩,郭加璇,李世杰,马兴红
(东北农业大学生命科学学院,黑龙江哈尔滨 150030)
小鼠早期妊娠通常是指妊娠第1~8 天,交配次日可见的阴道栓代表着雌雄小鼠成功交配,小鼠胚胎着床发生在妊娠第5 天,成功着床需要具有着床能力的胚胎与接受态子宫建立密切的相互作用联系[1-3],胚胎着床对于胚胎进一步的发育和成功妊娠至关重要。着床后,囊胚周围的梭形子宫内膜基质细胞迅速增殖并分化为上皮样分泌蜕膜细胞,这一过程称为蜕膜化。蜕膜基质细胞在保护胚胎免受局部母体免疫排斥、为生长胚胎提供营养、调节滋养细胞侵袭和胎盘形成等方面发挥着重要作用[4-6]。
弹性蛋白(Elastin)是细胞外基质蛋白的重要成员之一,由Eln基因编码,可与微纤维结合形成弹性纤维,进而赋予了组织一定的弹性回缩功能和可逆的变形能力[7-9]。有研究报道,弹性蛋白是肺部发育所必需的,Eln-/-基因型的小鼠出生时肺的远端气道发育受损而形成异常大的空腔[10-11]。弹性蛋白和其衍生肽也可调节平滑肌细胞的行为,促进健康血管的维持[12]。研究发现,弹性蛋白在维持正常的雌性生殖器官等生理功能上具有重要作用,其功能受损将导致盆底功能障碍性疾病的发生[13]。但有关Elastin在小鼠早期妊娠子宫中的表达和功能未见报道,本研究利用原位杂交和荧光定量PCR 方法检测了Elastin在早期妊娠、假孕及人工诱导蜕膜化模型小鼠子宫中的表达,进而探究Elastin的作用,为研究Elastin在大动物妊娠过程中的作用机制提供理论依据,对于提高畜牧动物繁殖率具有一定意义。
1 材料与方法
1.1 实验材料 本实验中所采用的模式动物为性成熟、6~10 周龄的ICR 品系的小白鼠,由实验室统一购买于黑龙江中医药大学。小鼠在室温22~24℃、湿度适合的人工饲养条件下成长,光照与黑暗循环交替(其中光照时间为07:00—19:00),在人工控制下饲料及饮水自由获取。
1.2 实验动物模型
1.2.1 早期妊娠模型 挑选若干只已满6 周龄性成熟的且正处于发情期、体征健康的雌性小鼠,按2:1 的比例于16:00 左右,与健康雄鼠合笼交配,在次日08:00 左右检栓,若阴道口出现白色栓块堵塞,则说明两者已成功交配,当天即记作该鼠早期妊娠的第1 天。按照此合笼检栓的方式,分别取材妊娠第1~8 天的小鼠子宫组织(每天取3 只),取妊娠第1~4 天子宫时,生理盐水冲输卵管或子宫以确定妊娠的发生;取妊娠第5 天子宫时,静脉注射0.1 mL 1%芝加哥天蓝染料,以确定小鼠胚胎着床位点;取妊娠第6~8 天子宫时,子宫有若干明显凸起,便可确定妊娠的发生,将组织经液氮速冻后放入-80℃冰箱保存。
1.2.2 假孕模型 挑选若干只6~8 周体征健康的雄性小鼠,进行结扎手术切除其双侧输精管,术后恢复2~3 周后,然后让其按1:2 比例与性成熟的发情期雌性小鼠合笼交配,在次日08:00 左右检栓以确认是否成功交配,若阴道口出现白色栓块堵塞,则把当天记作该鼠假孕第1 天,按此方式分别取假孕第1~5 天的小鼠子宫组织,每组取材3 只,于-80℃冰箱冻存。
1.2.3 人工诱导蜕膜化模型 取3 只假孕第4 天的雌鼠,用犬眠宝将其麻醉后,一侧子宫角注射25 μL 芝麻油以诱导蜕膜化,而另一侧子宫角不做任何处理,作为对照组。在假孕第8 天时,将小鼠处死,取子宫材料,于-80℃冰箱冻存。
1.3 实验方法
1.3.1 小鼠Elastin基因片段克隆及探针的制备 从NCBI 数据库中获取小鼠Elastin基因的全mRNA 序列(NM-007925.4)。然后,根据基因序列,使用Primer 5 软件和NCBI 设计引物(上游引物5'-GAAAACCCACA GGACAAGGA-3';下游引物5'-TAATGGGAGTCAGGG ACAGG-3'),设计的PCR 目的产物长度为401 bp。以小鼠子宫cDNA 为模板,PCR 扩增目的片段,采用Promega 的Wizard SV Gel and PCR Clean-Up System试剂盒胶回收PCR 产物,然后连接到pGEM-T 载体上,转化重组质粒于DH5α感受态细胞中,在超净工作台中随机挑取若干菌落,进行菌落PCR,对连接方向正确的菌落进行摇菌,用Axygene 质粒小提试剂盒提取质粒DNA,送哈尔滨擎科生物技术有限公司进行测序,以测序正确的质粒为模板进行PCR 扩增并纯化PCR 产物,利用Roche 公司的DIG RNA labeling kit 试剂盒对cRNA 片段进行地高辛标记。
1.3.2 原位杂交 将组织厚度为8 μm 的冰冻切片置于55℃的展片台上烘干,在4%多聚甲醛(PFA)中固定1 h,洗涤后于Triton X-100 溶液中透膜处理20 min,洗涤,室温预杂交15 min,弃预杂交液,配含有ElastincRNA 探针的杂交液,滴加于切片上,55℃杂交过夜;次日,在55℃水浴下,分别用含有50% 甲酰胺的2×柠檬酸钠缓冲液(SSC)洗涤切片15 min、2×SSC 洗涤30 min、0.2×SSC 洗涤30 min 2 次,在室温下,用不含甲酰胺的0.2×SSC 溶液洗涤5 min,Buffer 1 溶液清洗5 min 后滴加封闭液(HB)于切片上,封闭60 min 后滴加地高辛标记的碱性磷酸酶抗体,4℃过夜;第3 天,在室温下用Buffer 1 溶液清洗切片5 min 3 次,Buffer 2 溶液清洗5 min,滴加碱性磷酸酶显色液于切片上,出现信号后用甲基绿对染、封片及照相。
1.3.3 实时荧光定量PCR 本步骤采用TRIZOL(Invitrogen)提取小鼠子宫组织的总RNA,Nano Drop 2000(Thermo)测RNA 浓度,应用Promega 公司的GoScript Reverse Transcription Mix 反转录试剂盒,参照其说明书将RNA 反转录成cDNA,使用TaKaRa 公司的荧光定量PCR(TB GreenTMPremix Ex TaqTM) 试剂盒,以cDNA 为模板进行PCR,Elastin(NM-007925.4)引物序列:上游引物5'-GATGGTGCACACCTTTGTTG-3',下游引物5'-CCGGTATTTGGGTACCAATG-3',内参基因RpL7(M29016)上游引物:5'-GCAGATGTACCGCA CTGAGATTC-3',下游引物:5 '-ACCTTTGGGCTTACT CCATTGATA-3',反应体系10 μL:SYBR Premix Ex Taq(2×)5 μL,Rox Reference Dye II 0.2 μL,10 μmol/L 上、下游引物各0.2 μL,模板cDNA 1 μL,dH2O 3.4 μL。该反应采用Amplied Biosystems®7500 Real-Time PCR System(Amplied Biosystems)系统,反应条件:95℃30 s;95℃ 5 s、60℃ 30 s,40 个循环后收集荧光信号。利用2-ΔΔCt法来分析目的基因的相对表达。
1.3.4 统计分析 采用 GraphPad Prism 6.0 软件绘图并分析数据。差异性比较采用单因素方差分析或t 检验,统计学意义为P<0.05。
2 结果
2.1ElastinmRNA 在小鼠早期妊娠子宫中的表达 原位杂交结果显示,ElastinmRNA 在小鼠早期妊娠第1~4天子宫中没有表达,随着妊娠的进行,ElastinmRNA在第5~8 天的子宫壁肌层表达,并逐渐增强,其中ElastinmRNA 在第7、8 天的系膜侧蜕膜区也有较强的表达(图1-A)。荧光定量PCR 结果表明,与早期妊娠1~4 d 相比,ElastinmRNA 在5~8 d 小鼠子宫中显著表达,且相对表达量逐渐增加(图1-B)。
2.2ElastinmRNA 在小鼠假孕模型子宫中的表达 原位杂交结果表明,在假孕第1~4 天子宫中,未检测到ElastinmRNA 的表达(图2-A~E),这一表达模式与正常妊娠时类似,但在假孕第5 天子宫中未检测到ElastinmRNA 的表达(图2-F)。
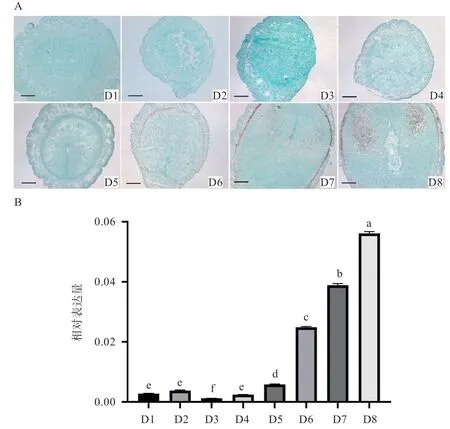
图1 ElastinmRNA 在小鼠早期妊娠子宫中的表达

图2 ElastinmRNA 在小鼠假孕模型子宫中的表达
2.3ElastinmRNA 在小鼠人工诱导蜕膜化子宫中的表达原位杂交结果显示,ElastinmRNA 在人工蜕膜子宫中的肌层及系膜侧的蜕膜区域有很强的表达(图3-A),而在对照子宫中,仅在肌层处检测到微弱的ElastinmRNA 表达(图3-B)。

图3 ElastinmRNA 在小鼠人工诱导蜕膜化子宫中的表达
3 讨 论
弹性蛋白是弹性纤维的主要成分,它的存在使得动物机体具有组织扩张以及缓冲等能力,弹性蛋白所维持的弹性纤维在不同组织器官中具有不同的形态结构[8]。有研究表明,弹性蛋白广泛分布于人子宫肌层和血管平滑肌中,其中在血管周围组织中也有表达[14]。子宫肌层在小鼠妊娠过程中起重要作用,从胚胎定位及胚胎着床一直到分娩启动,子宫肌层都扮演着重要的角色。在小鼠早期妊娠第1~4 天,子宫收缩一方面促进精子通过子宫到达输卵管受精,另一方面受精后的胚胎在妊娠第4 天到达子宫后,子宫收缩有利于胚胎在子宫中的分布。胚胎着床发生在妊娠第5 天,肌层收缩在胚胎着床后被抑制对妊娠的维持非常重要,这一过程通过多方面的激素调节。在哺乳动物妊娠中,孕酮受体(PR)和孕酮(P4)通过抑制收缩相关基因催产素受体(OXTR)、环氧合酶2(COX-2)等在肌层中的表达来维持子宫静止状态[15]。本实验发现,Elastin在小鼠早期妊娠第1~4 天子宫中检测不到表达,在胚胎着床后的妊娠第5 天开始表达,第6~8 天子宫肌层有强烈的表达,且表达量逐渐上升,这一结果说明,Elastin参与小鼠早期妊娠子宫肌层弹性的调控,Elastin的表达可能有助于着床后植入胚胎的稳定。与正常妊娠结果不同的是,Elastin在小鼠假孕第5 天子宫中不表达,在假孕过程中的激素水平与正常妊娠时基本相同,不同的是假孕第5 天没有发生胚胎着床,这提示Elastin可能受着床的胚胎或胚胎着床后诱导的蜕膜分泌的因子调控,通过调控子宫肌层的收缩活动参与妊娠后期维持子宫相对平静的内环境,维持妊娠进行。
在胚胎着床启动后,子宫基质细胞开始发生蜕膜化[16-17]。蜕膜化过程十分复杂,这一过程涉及到许多生物学事件,包括子宫基质细胞生长、基质成纤维细胞再分化、细胞外基质重建、细胞凋亡等[17-18]。分化后的子宫基质细胞对于母体子宫的重塑、血管的发育起到了有力的支持作用,也为后期胚胎的正常发育提供了保障[18-19]。本研究发现,Elastin不但在小鼠妊娠第7~8 天的子宫肌层中表达,还高表达于系膜侧的蜕膜区中。在假孕子宫腔内注射少量芝麻油可诱导啮齿动物发生蜕膜化[20]。这种人工诱导的蜕膜组织称为蜕膜瘤,它与妊娠期间形成的正常蜕膜组织相似。同样,Elastin也高表达于人工蜕膜子宫的肌层及系膜侧蜕膜区域。子宫系膜侧是一个不太致密的区域,对血管形成和胎盘的发育很重要[21]。以上结果说明,Elastin参与小鼠子宫蜕膜化过程,与胎盘的发育相关。
有研究表明,雌激素会影响弹性蛋白基因表达使其发生相关变化,人阴道壁成纤维细胞受到应力牵拉下,ElastinmRNA 表达量增多,对受应力后的细胞加入雌二醇,进一步促进了ElastinmRNA 的表达[13]。Lin 等[22]证明TGF-β1 可上调大鼠尿道细胞中ElastinmRNA 的表达,而雌激素干扰TGF-β1 信号通路可抑制弹性蛋白的表达,导致弹性纤维在尿道中的不当组装。也有科学家利用小鸡血管平滑肌细胞证明,肝素对增殖细胞中Elastin表达的上调是通过抑制蛋白激酶C 活性来介导的[23]。Kuang 等[24]证明,在大鼠新生肺成纤维细胞中,白细胞介素1β(IL-1β)可激活NF-κB,从而下调ElastinmRNA 的表达。NF-κB 的激活是IL-1β诱导弹性蛋白启动子转录活性下调所必需的。在人肺成纤维细胞中,TGF-β依赖于磷脂酰肌醇3-激酶(PI3K)/AKT信号激活Elastin的转录[25]。综上所述,在不同类型的细胞中,多种调节因子均可通过不同方式调控Elastin的表达,然而,关于Elastin在小鼠早期妊娠子宫肌层及蜕膜细胞中的表达受何种分子的调控,值得进一步研究。
一直以来,畜牧动物繁殖性能的高低是养殖行业面临的重要问题,较低的着床率、产仔率以及较高流产率都是导致其繁殖力低下的主要原因,这给畜牧业带来巨大的经济损失。本研究首次揭示了Elastin在小鼠早期妊娠中的表达规律,证明Elastin在小鼠妊娠子宫壁肌层弹性的调控与蜕膜化过程中发挥重要作用,Elastin可能通过调控子宫肌层的收缩活动参与妊娠后期维持子宫相对平静的内环境,进而维持妊娠的进行,防止了流产现象的发生,这一研究为后续在家畜中的研究奠定了实验基础,对于畜牧动物繁殖率的提高及治疗等具有重要的理论意义和潜在的经济价值。
4 结 论
本实验结果表明,ElastinmRNA 在小鼠早期妊娠第6 天的子宫壁肌层及第7、8 天的肌层和蜕膜区强表达,在人工诱导蜕膜子宫中肌层和系膜侧蜕膜区强表达。Elastin很可能参与调控小鼠早期妊娠子宫壁肌层的弹性及蜕膜化过程,进而在小鼠早期妊娠过程中发挥作用。
