高粱病程相关基因非表达子1(NPR1)基因家族鉴定及胁迫应答分析
2021-04-19杜巧丽方远鹏蒋君梅任明见
杜巧丽 方远鹏 蒋君梅 孙 涛 任明见 谢 鑫
(1贵州大学农学院农业微生物特色重点实验室,贵州 贵阳 550025;2国家小麦改良中心贵州分中心,贵州 贵阳 550025;3重庆海关技术中心,重庆 400020)
病程相关基因非表达子1 (non-expressor of pathogenesis related genes1,NPR1)是在研究系统获得性抗性(systemic acquired resistance,SAR)[1]信号传导途径时从拟南芥突变体中发现的,并以其突变体npr1 的表型命名[2-5]。NPR1 含有典型的多个锚蛋白重复序列Ankyrin 和1 个BTB/POZ 区,其结构域与蛋白-蛋白互作紧密相关[6-7]。NPR1 的表达在植物免疫系统中发挥着重要的作用,它是植物水杨酸(salicylic acid,SA)信号通路的重要组分之一。在拟南芥中,SA 可直接与NPR1 的同源蛋白NPR3 和NPR4 结合,并调节其与NPR1 的相互作用,影响NPR1 蛋白的稳定性[2,6,8];同时SA 也能激活植物的防御基因(如病程相关蛋白基因PR),从而产生系统获得性抗性,以保护植物免受病原菌的侵染[9]。在低温条件下,拟南芥细胞质中的NPR1 会转入细胞核,促使NPR1 与热休克转录因子1(recombinant heat shock transcription factor 1,HSFA1)相互作用,介导植物产生适应低温的调节途径,此时NPR1 可作为调节寒冷和接收病原体信号的中心枢纽,从而保证植物能更好地适应环境变化[10],说明NPR1 能响应逆境胁迫。
NPR1 同源基因广泛存在于植物中,现已作为抗病相关基因而被研究,并在拟南芥(Arabidopsis thaliana)[11]、小麦(Triticum aestivum)[12]、水稻(Oryza sativa)[13-14]、玉米(Zea mays)[15-16]、大麦(Hordeum vulgare)[17]、 杨 树(Populus)[18]和甘蔗(Saccharum officinarum)[19]等物种中均有报道,而在高粱[Sorghum bicolor(L.) Moench]中鲜见相关研究报道。
高粱属禾本科单子叶植物,是一种典型的旱粮经济作物,种植面积广泛。与其他禾谷类作物相比,高粱具有较强的生长力,兼具较强的耐盐碱性、抗旱性、耐高温等抗逆特性[20-21]。高粱为C4光合作用植物,具有光合效率高,生物产量高等特点,享有“沙漠中的骆驼”称号[22-23]。据报道,在水稻[13]、玉米[16]、大麦[17]、甘蔗[19]和油菜[24]等多种植物中,过表达NPR1 基因可有效提高植物对白叶枯病、粗缩病、赤霉病、黑穗病、菌核病等病害的抗性,说明NPR1 基因在植物抗病过程中起着重要作用。因此,本研究从全基因组水平出发,对高粱NPR1 家族成员进行鉴定,通过系统进化树构建,基因结构分析和染色体定位等生物信息学方法研究其基本特征,并通过实时荧光定量PCR(quantitative real-time PCR,RT-qPCR)方法检测SbNPR1 基因在不同组织、不同激素以及病原相关分子模式(pathogenassociated molecular pattern receptors,PAMPs)胁迫条件下的相对表达量,旨在为进一步探究该家族基因在调节高粱响应植物激素以及病原菌胁迫应答反应方面提供基础。
1 材料与方法
1.1 试验材料
1.1.1 高粱品种及处理方法 本研究所用高粱品种BTx623 由贵州大学植物病理教研室保存。高粱种子经表面消毒后,在灭菌水中浸种24 h,之后用纱布保湿培养72 h 进行催芽,将发芽的种子种植于灭菌营养土中,在25℃温室中进行昼夜交替培养,待长到三叶期,用独脚金内酯(strigolactone,GR24,1 μmol·L-1)、 SA(1 mmol·L-1)、氯化钠(NaCl,250 mmol·L-1)、聚乙二醇(polyethylene glycol,PEG6000,20%)、甘露醇(Dmannitol,300 mmol·L-1)、flg22(100 nmol·L-1)和几丁质(Chitin,8 nmol·L-1)处理,分别在0、0.5、1、3、6、9、12 和24 h 取样,设置3 个生物学重复,每个重复处理5 株苗,将所取样品立即用液氮进行速冻,置于-80℃冰箱保存备用。
1.1.2 试剂 本试验中所用的反转录试剂盒购于普洛麦格(北京)生物技术有限公司,RT-qPCR 反转录试剂盒购自北京康润诚业生物科技有限公司。试验所用激素,盐胁迫和干旱胁迫处理剂均购自北京酷来搏科技有限公司。flg22 购自南京金斯瑞生物科技有限公司,Chitin 购自圣克鲁斯生物技术(上海)有限公司。
1.1.3 引物设计 高粱SbNPR1 基因的RT-qPCR 正向引物为SbNPR1-qF:5′-ACCAATATTCTTCCCTTCCCA A-3′,反向引物为SbNPR1-qR:5′-AACAATGTGGGAAG TATCGTCT-3′。高粱内参基因SbEIF4a的RT-qPCR 正向引物为SbEIF4a-qF:5′-AGGATTGGCACCAGAAGGG T-3′,反向引物为SbEIF4a-qR: 5′-CACATCAAGCCCCT TGCAGA-3′。以上引物均由北京擎科新业生物技术有限公司合成。
1.2 高粱SbNPR1 基因家族鉴定、亚细胞定位、染色体定位及基因结构分析
通过Hmmer 3.0 软件以拟南芥、水稻建立的序列徽标检索高粱数据库(Phytozome),基于SMART 和NCBI-CD 网站检测保守结构域;SbNPR1 家族蛋白的基本理化性质用ExPASy-ProtParam tool 预测;蛋白亚细胞定位利用网站http:/ /www.csbio.sjtu.edu.cn/bioinf/Cell-PLoc-2/进行;染色体定位及基因结构分析分别在网站 http:/ /mg2c. iask. in/mg2c _ v2.0/及http:/ /gsds.cbi.pku.edu.cn/进行。
拟南芥NPR1 基因家族的序列均源于拟南芥数据库(https:/ /www.arabidopsis.org),甘蔗、水稻、玉米、高粱NPR1 基因家族的序列分别来源于(https:/ /sugarcane-genome. cirad. fr/)、 MSU ( http:/ /rice.plantbiology.msu.edu/)和Phytozome 数据库。
1.3 高粱SbNPR1 基因家族系统发育树及蛋白保守功能基序分析
利用MEGA 7.0 软件,以邻接法(neighbor-joining,NJ;bootstrap =1 000)构建NPR1 基因家族系统发育进化树,使用Evolview 对所构建的进化树进行美化,蛋白保守功能基序分析通过MEME 软件进行。
1.4 高粱SbNPR1 基因表达分析
以高粱植株cDNA 为模板,通过RT-qPCR 检测SbNPR1 基因的表达量,扩增体系为:cDNA 4.5 μL,SYBR Premix 7.5 μL,上下游引物各0.3 μL,灭菌去离子水补至15 μL。反应程序:95℃预变性10 min,95℃变性15 s,55℃复性15 s,72℃延伸20 s,40 个循环,荧光定量数据以2-ΔΔCt方法进行分析,并利用SPSS 软件进行显著性分析。
2 结果与分析
2.1 高粱SbNPR1 基因家族鉴定
运用Hmmer 程序依据拟南芥和水稻的NPR1 家族全蛋白序列构建Hmmer 模型,检索高粱数据库获得E 值≤1×10-5的序列,利用SMART 及NCBI-CDD 数据库验证其保守结构域,保留含有BTB/POZ 区和Ankyrin 锚蛋白结构域的蛋白序列,去除冗余序列。共鉴定到5 个NPR1 成员,分别命名为SbNPR1~SbNPR5(表1)。其中SbNPR5 编码的氨基酸序列最短(480 aa),SbNPR2 编码的氨基酸序列最长(621 aa);该家族分子量介于50.496 81~67.648 06 kDa;理论等电点介于5.64~6.11 之间,表明这5 个成员基因编码的蛋白呈弱酸性;利用MG2C 网站对高粱的NPR1 基因家族成员进行染色体定位分析,发现3 号染色体上分布3个基因,分别是SbNPR1、SbNPR2 和SbNPR3;5 号染色体上分布1 个基因,为SbNPR4 基因;8 号染色体分布一个基因,为SbNPR5 基因;亚细胞定位预测显示,该家族基因分别位于细胞核和细胞质(表1)。

表1 SbNPR1 基因家族编码蛋白的基本特征Table 1 Basic characteristics of the putative proteins encoded by SbNPR1 gene family
2.2 NPR1 基因家族系统进化分析
利用MEGA 7.0 将高粱NPR1 家族蛋白序列与拟南芥、水稻、玉米及甘蔗的NPR1 家族蛋白序列进行系统发育进化树的构建,如图1 所示。高粱中NPR1 基因家族成员在3 个分支中均有分布,在第Ⅰ分支中高粱SbNPR2 与AtNPR3、AtNPR4 及OsNPR3、ZmNPR1等蛋白聚在一支,推测SbNPR2 可能参与对SAR 的负调控[25]; 在 第Ⅱ分支中包括 SbNPR1、 AtNPR1、AtNPR2、Sh253P03、Sh203L02 及OsNPR1 等成员,其中SbNPR1 与Sh253P03、Sh203L02 亲缘关系最近,其次是Zm00008a010809,且OsNPR1 已证实在提高植株抗病性中起着重要作用[13],因此推测SbNPR1、Sh253P03、Sh203L02 及Zm00008a010809 可能参与对SAR 的正调控;在第Ⅲ分支中,包括高粱3 个成员SbNPR3、SbNPR4 及SbNPR5。从整体来看NPR1 基因在进化过程中相对保守。
2.3 高粱NPR1 基因家族结构分析
通过GSDS 软件绘制高粱NPR1 基因家族的内含子-外显子基因结构图,如图2 所示。该家族基因内含子数目为2 ~4 个,其中SbNPR1、SbNPR2 均被3 个内含子分割为4 个片段;SbNPR3、SbNPR4 和SbNPR5均被1 个内含子分割为2 个片段。此外,该基因家族都具有编码和非编码两个区域,其中SbNPR3、SbNPR4和SbNPR5 这3 个基因均含有2 个编码区,而SbNPR1、SbNPR2 含有4 个编码区。
2.4 高粱NPR1 基因家族蛋白保守motif 分析及多序列比对分析
为了对高粱SbNPR1 家族蛋白保守motif 进行探究,利用MEME 软件预测了10 个保守motif。由图3可知,motif 1、motif 2、motif 3、motif 4、motif 5、motif 6 为所有成员都具有的保守基序,而motif 8、motif 9 仅出现在SbNPR3、SbNPR4 和SbNPR5 中,motif 10 出现在SbNPR4 和SbNPR5 中;另外,motif 8、9 和10 仅存在于SbNPR4 和SbNPR5 中,推测这些基因可能具有某些特殊功能。
将NPR1 与其他物种中的同源氨基酸序列进行比对,并使用SMART 网站分析NPR1 氨基酸序列,发现所有成员都含有NPR1 基因所具有的典型结构域,BTB/POZ 结构域、ANK-repeat 锚蛋白重复序列结构域以及NPR1_like_C-terminal 结构域,且NPR1_like_C-terminal结构域包含核定位信号NLS 位点,该位点与其在核内参与转录调控相关[9]。利用DNAMAN 进行多序列比对,结果表明,多序列同源性比对结果为62.82%;各个结构域之间的同源性分别为:BTB/POZ 结构域62.60%、ANK-repeat 锚蛋白重复序列结构域67.59%、NPR1_like_C-terminal 结构域70.02%(图4)。
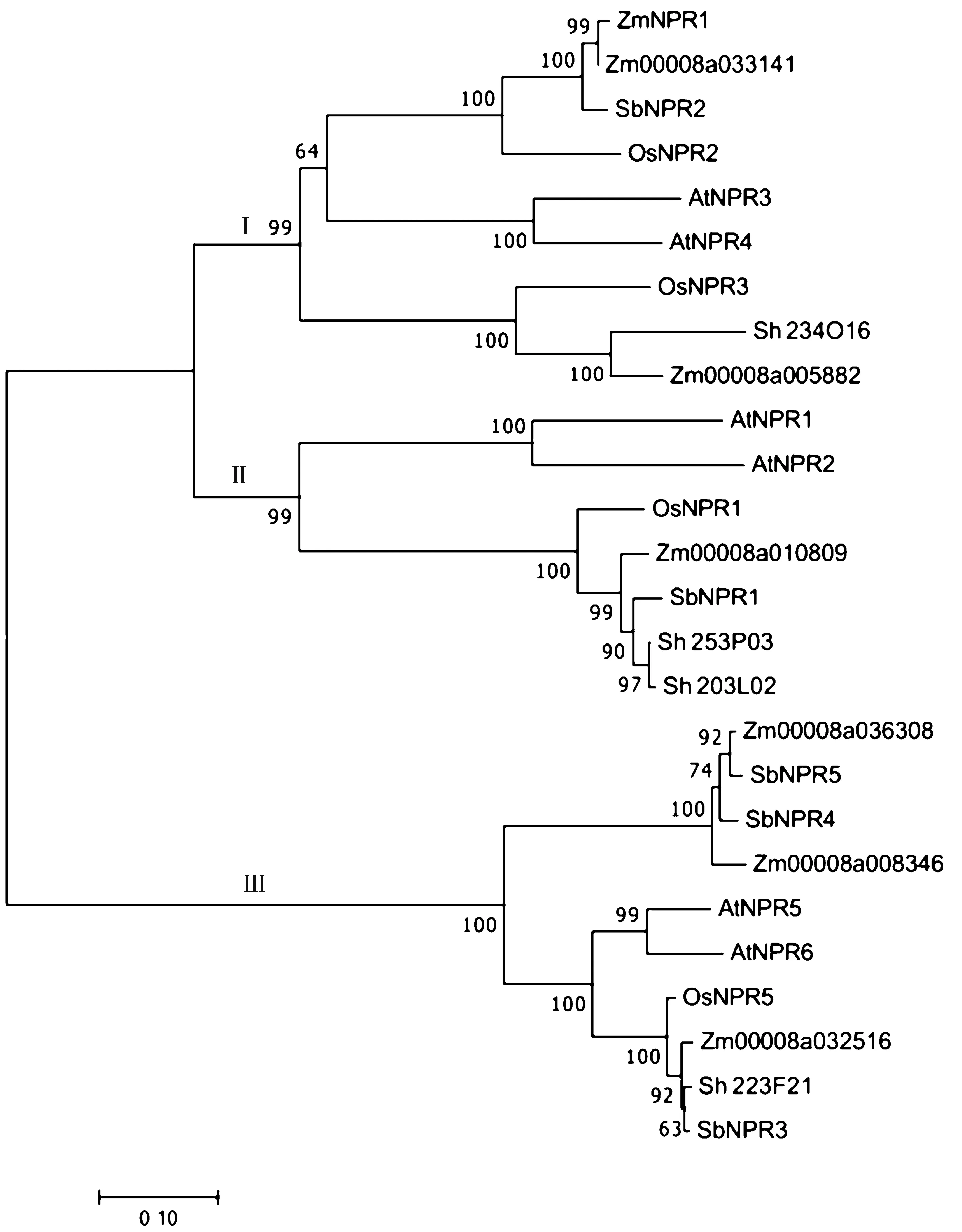
图1 NPR1 基因家族进化分析Fig.1 Evolutionary analysis of NPR1 gene family

图2 SbNPR1 基因家族成员的结构分析Fig.2 Gene structure analysis of SbNPR1 gene family
2.5 SbNPR1 基因表达分析
系统进化分析发现,在5 个高粱NPR1 基因中,由于SbNPR1 基因与水稻OsNPR1 位于同一分支,亲缘关系较近(图1),且OsNPR1 已被证明为SA 的受体基因,因此本研究选取SbNPR1 基因进行进一步的表达分析。

图3 SbNPR1 基因家族保守基序分析Fig.3 Conserved motif analysis of SbNPR1 gene family
2.5.1SbNPR1 基因组织表达特征 对高粱不同组织SbNPR1 基因的表达进行分析,发现其在不同的组织器官中表达不同,具有组织特异性,其在叶中的表达量显著高于根、茎和芽,叶中的相对表达量约是根的10.4 倍(图5)。
2.5.2SbNPR1 基因响应激素表达分析SbNPR1 在GR24 和SA 不同作用时间下的表达模式分析结果表明,SbNPR1 的表达量随着GR24 处理时间的延长而显著降低,SbNPR1 表达量受到了一定程度的抑制(图6-A)。但SA 处理3 h 时SbNPR1 表达量达到最大值,约为对照组的4.4 倍,说明SA 诱导了SbNPR1 的表达(图6-B)。
2.5.3SbNPR1 基因响应干旱及盐胁迫表达分析 用PEG6000、D-mannitol、NaCl 模拟干旱及盐胁迫,分析SbNPR1 基因表达。结果表明,经PEG6000 处理0.5 h时,SbNPR1 的表达显著上调且达到最大值,之后迅速降低(图7-A);经甘露醇处理后,SbNPR1 的表达呈先升后降的模式,其相对表达量在3 h 时达到最大值(图7-B);在NaCl 处理下,SbNPR1 的表达在0.5 h 时显著上调并达到最大值,之后下降(图7-C)。以上结果说明SbNPR1 可能是高粱响应干旱及盐胁迫的一个功能基因。
2.5.4SbNPR1 基因响应病原相关分子模式(PAMPs)表达分析 为检测PAMPs 激发子flg22 和Chitin 是否影响SbNPR1 的表达量,分别用flg22 和Chitin 处理高粱,发现经flg22 处理后,SbNPR1 的表达量在12 h 时表达量显著升高并达到最大值(图8-A)。相反,经Chitin 处理后SbNPR1 的表达量从处理后6 h开始受到抑制(图8-B)。上述结果表明flg22 和Chitin分别激活和抑制SbNPR1 基因的表达,暗示SbNPR1 参与不同的植物先天免疫应答过程。
3 讨论
植物在生长发育过程中会受到一些病原物(如细菌、真菌、病毒等)的侵染,但植物自身也存在能够抵御病原物侵染的免疫应答反应。NPR1 基因在植物的抗病过程中起着核心调控作用,它能提高植物抵御多种病原物侵害的能力,是植物体内多种抗病通路途径之间的一个交叉位点[26]。已有报道表明,AtNPR1 基因的组成性表达能诱导水稻产生抗病性[27],同时在拟南芥、水稻和西红柿中过表达NPR1 基因,能提高对细菌和真菌的抗性[19],表明在植物防御系统中NPR1 起着极其重要的作用;当NPR1 在转基因拟南芥中过量表达时,能提高拟南芥对逆境胁迫的抗性[28];并且在没有病原体侵染的时候,也能对基因的转录进行调节[29]。因此,对高粱NPR1 基因家族进行鉴定和分析对于提高高粱抗性具有重要意义。

图4 SbNPR1 与其他物种的同源氨基酸序列比对Fig.4 Amino acids alignment between SbNPR1 and other species

图5 SbNPR1 基因的组织特异性表达Fig.5 Tissue specific expression of SbNPR1 gene
基因表达分析表明,经过SA 处理后的高粱,其表达量呈现出先升后降的模式,在处理3 h 时,NPR1 表达量显著升高并达到最大值,之后显著降低,说明SA会影响NPR1 基因在植株内的表达,这与玉米[16]、甘蔗[19]、梨[30]、小麦[31]、草地早熟禾[32]、苹果[33]、花生[34]等物种中报道的结果相似。在拟南芥中,NPR1在SA 存在的情况下,能通过RNA 聚合酶Ⅱ聚集类似CDK8、WRKY18 和TGA等转录因子,从而促进靶基因和自身的表达启动植物免疫信号[35];在无SA 的情况下,NPR1 主要是以胞质形式而存在,而当植物受到病原侵染时,SA 的累积会促使NPR1 转移到细胞核,与转录因子TGA相互作用,启动靶基因的表达[36]。研究报道,超表达NPR1 基因可提高植物的抗病性。如在甘 蔗[19]、 芥 菜[37]、 棉 花[38]、 小 麦[39]、 花 生[40]、 苹果[41]、胡萝卜[42]、番茄[43]和水稻[13]中过表达NPR1可分别提高对黑穗病、黑斑病菌、黑腐病、赤霉病、黄曲霉、白粉病、细菌性叶斑病、番茄花叶病毒和稻瘟病的抗性。
本研究显示,外源SA 处理3 h 时,高粱SbNPR1基因的相对表达量显著升高,说明SA 能影响SbNPR1基因在植株内的表达,但是否SbNPR1 基因的表达能提高高粱抗逆性还需进行基因功能学证实。高粱SbNPR1 基因的表达具有组织特异性,在叶中的相对表达量最高,根中最少,这与NPR1 基因在甘蔗组织中表达具有相似性[19],但在花生中,NPR1 主要在根和叶中表达,在茎中表达量较少[44],而拟南芥AtNPR1、水稻OsNPR1 均无组织特异性表达[45],由此推测NPR1在不同物种中其组织表达具有差异性。
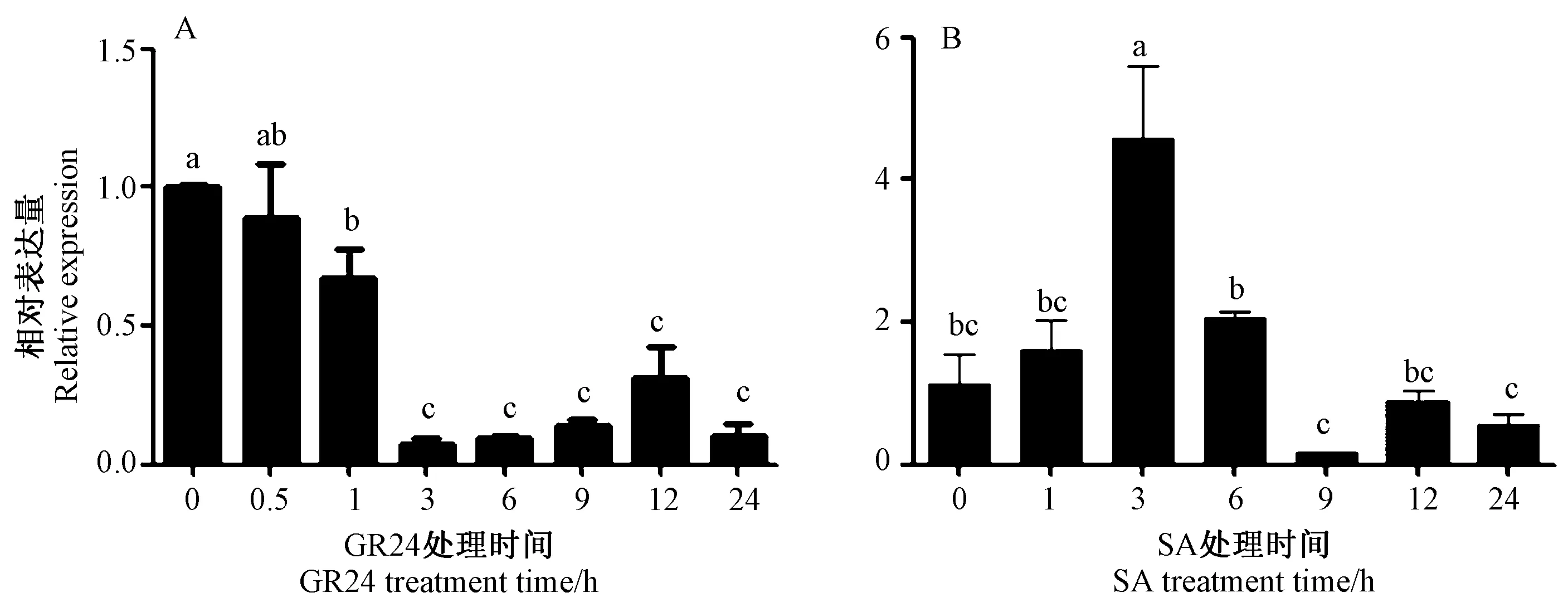
图6 SbNPR1 基因的激素响应表达Fig.6 Expression of SbNPR1 under hormone treatments

图7 SbNPR1 基因的干旱及盐胁迫表达Fig.7 Expression of SbNPR1 gene under drought and salt stress

图8 SbNPR1 基因响应PAMPs 表达分析Fig.8 Expression analysis of SbNPR1 gene under PAMPs treatments
通过PEG6000、甘露醇、NaCl 模拟逆境胁迫处理条件下的基因表达特征,发现SbNPR1 基因在逆境胁迫中,在处理较短时间内表达量显著下降,这与苹果MpNPR1-2 在干旱胁迫下较长时间内处理才会下降的基因有所不同,在持续两周的干旱胁迫结束时,根系中的MpNPR1-2 基因的表达量显著升高[46],因此在SbNPR1 在根系中是否出现此情况还有待进一步研究。研究表明,病原体相关分子模式(pathogenassociated molecular pattern receptors,PAMPs)不仅能激活蛋白激酶( mitogen-activated protein kinase,MAPKs)途径,而且还能诱导植物引发防御信号途径[47],本研究通过检测PAMPs 激发子flg22 和Chitin对SbNPR1 基因表达量的影响,发现flg22 能快速触发SbNPR1 基因的表达,而Chitin 则相反,抑制了SbNPR1基因的表达。拟南芥幼苗在经过flg22 处理后,也能诱导NPR1 快速表达[48],与本研究flg22 试验结果具有一致性,说明PAMPs 激发子flg22 在调控SbNPR1 基因的表达过程中起作用。本研究对高粱SbNPR1 基因的表达分析为今后进一步研究NPR1 基因的功能奠定了基础。
4 结论
本研究利用生物信息学方法从高粱数据库共鉴定到5 个高粱NPR1 家族成员,该家族成员均含有一个BTB/POZ 结构域和一个或多个Ankyrin 锚蛋白结构域,所编码的蛋白序列长度介于480 ~621 aa 之间,所预测的蛋白质分子质量介于50 ~67 kDa 之间,其理论等电点为5.64 ~6.11。RT-qPCR 结果表明,SbNPR1基因的表达具有组织特异性,且受到植物激素、干旱和PAMPs 胁迫的影响。本研究为进一步揭示NPR1 家族基因的生物学功能,及其在调节高粱抗性和响应植物激素等过程中的作用提供了基础。
