小麦盐胁迫响应相关ERF基因的分离和初步验证
2021-04-19李世姣张晓军乔麟轶贾举庆常利芳张树伟畅志坚
李世姣 张晓军 乔麟轶 贾举庆 常利芳 张树伟 畅志坚 李 欣
(1山西大学生物工程学院,山西 太原 030006;2山西农业大学农学院,山西 太谷 030800)
土壤盐渍化是影响农作物产量的主要限制因素之一。目前全球遭受盐害的耕地超过8 亿hm2,其中我国盐渍土面积达1 亿hm2[1-2]。随着粮食需求的不断加大,增强作物对盐渍土壤的耐受性进而提高其产量,对于保障粮食安全意义重大。小麦(Triticum aestirumL.)是世界上种植面积最广的粮食作物,广泛发掘小麦耐盐基因、培育耐盐新品种是充分利用盐渍化土壤的有效途径。
隶属AP2/ERF 超家族的乙烯响应因子(ethylene response factor,ERF)在植物应对盐胁迫过程中发挥着重要作用[3]。模式作物拟南芥中,除ERF 以外,AP2/ERF 超家族还包括干旱应答元件结合蛋白(dehydration responsive element binding,DREB)、 PETALA2 蛋 白(PETALA2,AP2)、与ABI3 和VP1 相关蛋白[(related to ABA-insentive3(ABI3)/viviparous1,RAV]和单独亚族(Soloist)[3]等4 个家族。其中,ERF 和DREB 家族序列相似性较高,均包含1 个AP2/ERF 结构域,但DREB 第14 和第19 位氨基酸分别是缬氨酸(V)和谷氨酸(E),而ERF 则是丙氨酸(A)和天冬氨酸(D)[3]。目前拟南芥ERF 家族成员ERF1[4]和ERF3[5]被证实可以提高植株耐盐性。此外,水稻(Oryza sativaL.)OsERF922[6]、大豆(Glycine maxLinn. Merr.)GmERF3[7]和GmERF7[8]、番茄(Solanum lycopersicum)TERF1[9]、JERF3[10]和SlERF5[11]、甘蔗(Saccharum officinarum)SodERF3[12]、烟草(Nicotiana tabacumL.)NtTOE3[13]等10 余个盐胁迫响应相关ERF基因已被鉴定。
目前小麦中共有8 个ERF基因被鉴定[14-17],其中有3 个基因与盐胁迫相关:TaERF1 在小麦受盐胁迫后表达水平上调,过表达TaERF1 的拟南芥植株耐盐性增强[14];TaERF3 过表达的小麦植株耐盐性增强,病毒诱导基因沉默(virus induced gene silencing,VIGS)干扰植株则表现为盐敏感[15];而TaERF4 过表达的拟南芥植株对盐胁迫的敏感性增强[16]。其他盐胁迫相关ERF基因的报道还鲜见。本研究从普通小麦全基因组中分离了ERF 家族,通过系统发育分析和转录组数据分析推测可能与盐胁迫相关的ERF 成员,并进行NaCl 处理下的实时荧光定量 PCR ( real time quantitative PCR,RT-qPCR)验证,以期发掘新的小麦耐盐基因。
1 材料与方法
1.1 植物材料
耐盐小麦材料CH7034(由山西省农业科学院畅志坚研究员选育)和盐敏感品种SY95-71[18]用于盐胁迫处理及RT-qPCR 试验。CH7034 种子萌发后的胚芽鞘以及幼苗的根、茎、叶用于基因组织表达水平分析。
1.2 盐胁迫处理
将小麦种子用1%的双氧水消毒后放置于培养皿中萌发,待胚根伸长至2~3 cm 时,选择长势一致的种子转移到培养箱中的1/2 霍格兰氏营养液中生长,参数设置为16 h 光照/8 h 黑暗,24℃/16℃;待幼苗生长至两叶一心期时更换含250 mmol·L-1NaCl 的营养液进行盐胁迫,在0、6 和12 h 时剪取幼苗根,于-80℃保存,用于RT-qPCR 分析。按上述方法萌发小麦CH7034,在芽期和苗期分别取胚芽鞘和根、茎、叶,于-80℃保存,用于基因组织表达水平分析。
1.3 序列分离与系统发育树构建
利用AP2/ERF 超家族的隐马尔可夫模型文件(注册号:PF00847.20)在nhmmer 软件[19]中检索普通小麦品种中国春IWGSCv1.0 注释蛋白序列(下载自URGI 数据库,http:/ /wheat-urgi.versailles.inra.fr/),设置E≤1e-5,获得小麦AP2/ERF 序列,将其与109 个拟南芥ERF 家族成员在Clustal X 软件[20]中进行序列多重比对,并用MEGA 6.0 软件[21]构建系统发育树,根据聚类结果以及特征序列分离出小麦ERF 家族。根据NCBI 注册号下载19 个已克隆盐胁迫相关ERF基因的编码蛋白序列,将其与小麦ERF 序列在MEGA 6.0 中构建系统发育树。
1.4 基因表达谱与启动子元件分析
小麦耐盐品种Arg 和盐敏感品种Moghan3 受NaCl 胁迫12 h 后的根部转录组数据[22](注册号:SRP158842)下载自NCBI 数据库(http:/ /www.ncbi.nlm.nih.gov/sra/),利用tophat2 将reads 比对到参考基因组IWGSCv1.0 上,并利用cufflinks 进行转录本组装[23],之后利用edger[24]获得基因序列的FPKM(expression fragment per kilobase of exon model per million mapped reads)值。从中检索并筛选表达量差异显著的TaERF 序列,筛选标准为:多重假设检验(FDR)值小于0.01、且|log2(FPKM[t-t0]/FPKMCK[t-t0])|≥1。所得结果用MeV tool (http:/ /www.tm4.org/mev.html)输出。
利用PlantCARE 数据库(http:/ /bioinformatics.psb.ugent.be/webtools/plantcare/html/)对盐胁迫响应相关ERF基因起始密码子前2 000 bp 的基因组序列进行启动子元件分析。
1.5 RT-qPCR
用RNA 提取试剂盒(北京天根生物技术有限公司)提取样品的总RNA,利用反转录试剂盒(英杰生命科技有限公司)将总RNA 反转录为cDNA。RTqPCR 在罗氏LightCycler © 96-PCR 仪上进行,使用的酶为SYBR Premix Ex Taq Ⅱ(北京宝日医生物技术有限公司),内参基因为小麦Tubulin,所用引物列于表1。每个反应重复3 次,所得结果采用2-ΔΔCT法[25]进行分析。
2 结果与分析
2.1 TaERF 家族的分离
利用隐马尔可夫模型文件检索小麦品种中国春数据库得到559 条AP2/ERF 序列,将其与拟南芥AtERF家族进行聚类,共分为7 个组(图1-A),其中有5 个组(组1~3,组6,组7)包含AtERF成员,从上述5 个组中进一步筛选结构域第14 和第19 位分别为丙氨酸(A)和天冬氨酸(D)的序列,获得229 条小麦ERFs(图1-B),在小麦A、B、D 基因组中的数目为70、78 和81,在第1 至第7 同源群分布数目依次为36、45、20、30、34、39 和25 (图1-C)。通过分析A/B/D 同源关系将其归为96 个TaERF成员。
2.2 TaERF 与已知耐盐相关ERF 的聚类分析
对96 个TaERF成员与19 个已报道耐盐相关ERF编码的蛋白序列进行系统发育分析。结果显示,有5 个TaERF成员与耐盐相关ERF聚在同一支中(图2)。其中,TaERF88 和TaERF27 分别是已报道的TaERF1[14]和TaERF3[15];而TaERF76、TaERF35 和TaERF85 则分别是拟南芥AtERF3[5]、 水 稻OsERF922[6]和簇毛麦(Haynaldia villosa)DvERF[26]的同源基因。

表1 RT-qPCR 引物Table 1 Primers for RT-qPCR
2.3 数据库中TaERFs 对盐胁迫的响应
基于转录组数据库的分析结果显示,TaERF家族中有9 个成员(共18 个ERF 序列)在小麦耐盐品种受盐胁迫后的表达水平变化显著,而在盐敏感品种中变化不显著(图3)。其中,TaERF38 在受NaCl 胁迫12 h后表达量下调,其余8 个成员TaERF3、TaERF27、TaERF28、TaERF55、TaERF56、TaERF64、TaERF67 和TaERF68 在受胁迫后表达量均上调。
2.4 TaERFs 受盐胁迫诱导
基于上述分析结果,与已克隆耐盐ERF基因序列相似性高、或受NaCl 诱导的TaERF成员共有13 个。利用250 mmol·L-1NaCl 对小麦耐盐材料CH7034 和盐敏感材料SY95-71 的两叶一心期植株进行盐胁迫处理,验证这13 个成员在根部的表达量变化。RTqPCR 结果显示(图4),TaERF27、TaERF35、TaERF55和TaERF64 共4 个基因在耐盐材料中受NaCl 胁迫后显著上调,而在盐敏感材料中无明显变化,可能为盐胁迫响应基因。
2.5 盐胁迫响应相关TaERF 的组织表达水平和启动子调控元件分析
上述4 个盐胁迫响应相关成员的序列信息列于表2。其中,TaERF27、TaERF35 和TaERF64 无内含子,只含有1 个外显子,而TaERF55 含有2 个外显子。
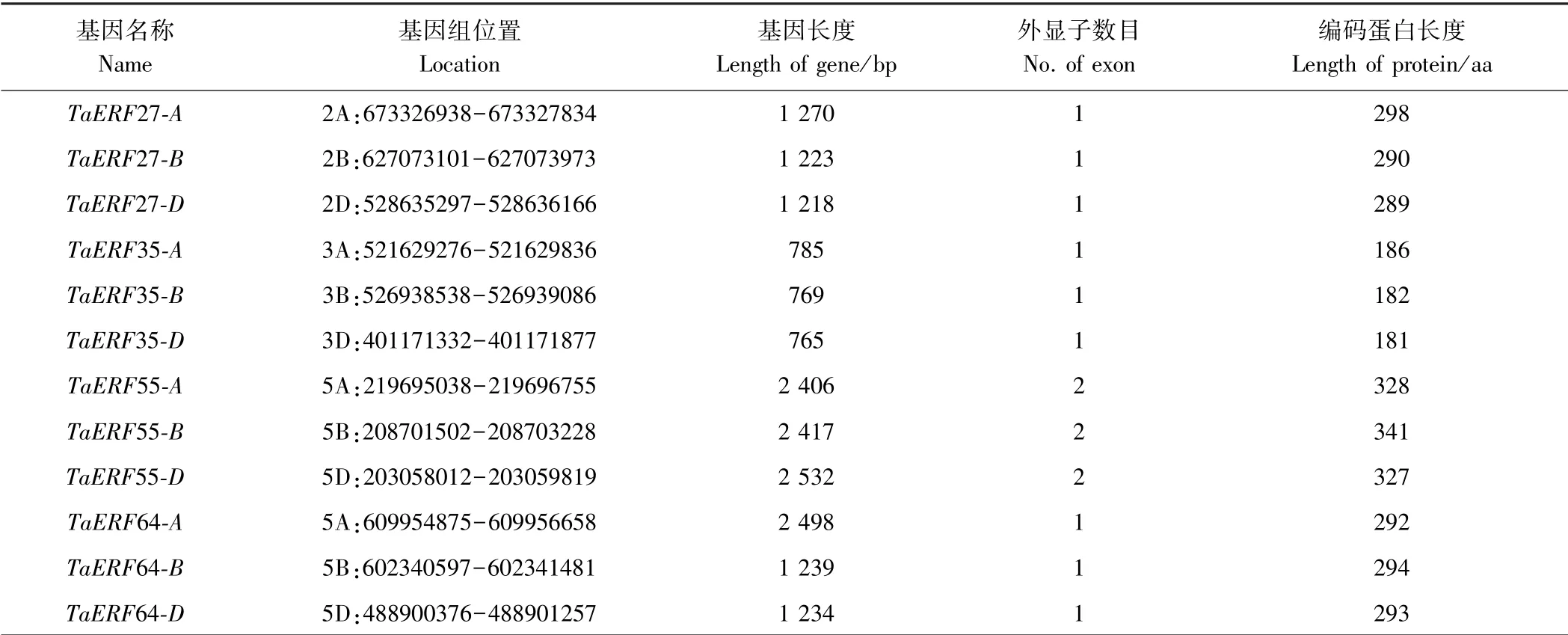
表2 盐胁迫响应相关TaERF 成员信息Table 2 Information of TaERF members related salinity response
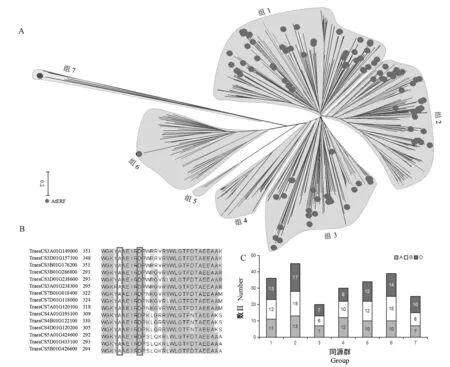
图1 基于聚类结果和结构特征从小麦AP2/ERF 超家族中分离ERF 家族Fig.1 Isolation of ERF family from wheat AP2/ERF superfamily based on clustering results and sequence characteristics
对盐胁迫响应相关成员TaERF27、TaERF35、TaERF55、TaERF64 在耐盐材料CH7034 萌发和幼苗阶段不同组织中的表达水平进行分析(图5),结果显示,上述4 个成员在苗期根和叶中具有较高的表达水平,而在胚芽鞘和茎中的表达量相对较低。其中TaERF35 和TaERF64 分别在苗期根和叶中表达量最高。
随后对4 个成员起始密码子前2 000 bp 基因组序列内包含的调控元件进行分析(图6),结果显示,上述成员除启动子元件TATA-box 和增强子区域调控元件CAAT-box,还含有参与茉莉酸响应的顺式元件CGTCA-motif、参与脱落酸响应的顺式元件ABRE、生长素响应元件TGA-element、参与水杨酸响应的顺式元件TCA-element 以及赤霉素响应元件P-box。此外,TaERF27 还含有GARE-motif、AuxRR-core、TATC-box元件,TaERF55 还含有GARE-motif、TATC-box 元件,TaERF64 还含有AuxRR-core 元件。各个调节元件功能见表3。
3 讨论
耐盐基因的发掘利用是改良小麦品种耐受性,降低土壤盐害的有效措施。目前小麦中除TaERF1[14]和TaERF3[15]外,还 有HTK[27]、SRO[28]、OPR1[29]和AOC1[30]等几个重要耐盐基因被鉴定。其中,HTK在育种中的应用使小麦在盐碱地上的籽粒产量提高了25%[27]。因此,持续发掘耐盐基因对于保障小麦高产稳产具有重要意义。
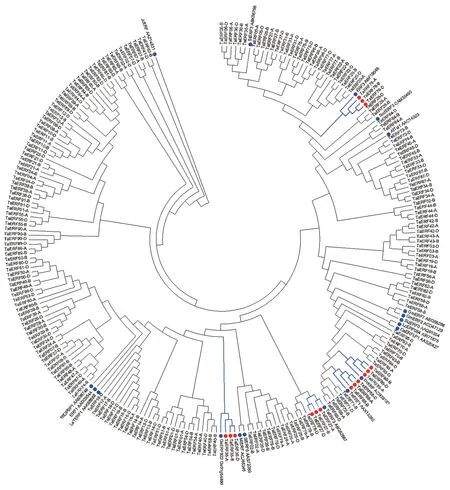
图2 TaERF 家族与19 个已克隆耐盐ERF 基因的系统发育树Fig.2 Phylogenetic tree of TaERF family and nineteen cloned salinity-tolerance ERFs

图3 耐盐品种Arg 和盐敏感品种Moghan3 受NaCl 处理12 h 后表达水平差异显著的TaERFsFig.3 The TaERFs expressed significantly differently between salt-tolerant Arg and salt-sensitive Moghan3 after NaCl treatment for 12 hours
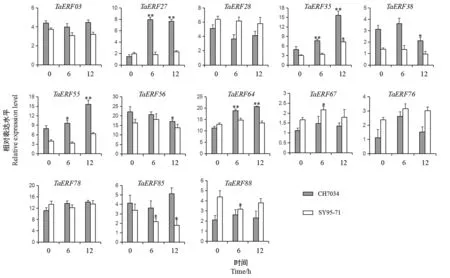
图4 13 个TaERF 成员在小麦耐盐材料CH7034 和盐敏感品种SY95-71 受NaCl 胁迫处理的表达水平Fig.4 Expression levels of 13 chosen TaERF members in wheat tolerant material CH7034 and sensitive variety SY95-71 after NaCl treatment
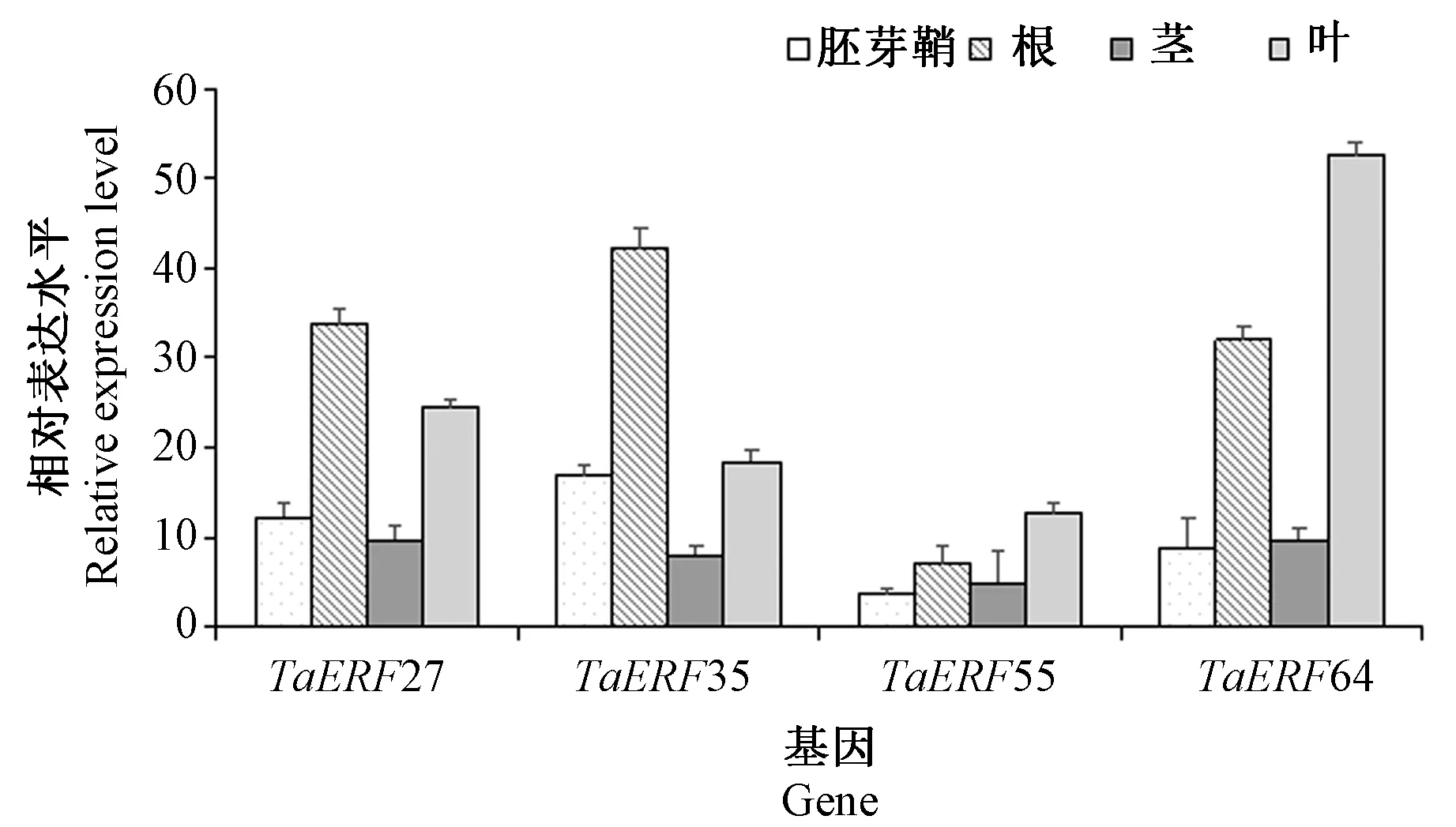
图5 4 个盐胁迫响应相关TaERF 成员在小麦耐盐材料CH7034 萌发和幼苗阶段不同组织中的表达水平Fig.5 Expression levels of four TaERF members related salinity response in wheat tolerant material CH7034 at germination and seedling stage
本研究从小麦全基因组中系统地分离了96 个TaERF家族成员,其中有两个是已报道的TaERF基因。TaERF88 与已报道的耐盐基因TaERF1 为同一基因(图2),但盐胁迫处理后TaERF88 在耐盐品种CH7034 中的表达水平无明显变化,推测CH7034 中可能有TaERF88 的一个新的等位变异,或者受材料遗传背景的影响,导致表达水平的变化;TaERF27 是已报道的耐盐基因TaERF3,研究表明TaERF3 蛋白可通过GCC-box 顺式元件来调控下游应激相关基因,进而提高小麦对盐和干旱胁迫的适应性。本研究通过利用耐盐材料CH7034 和盐敏感材料SY95-71 还鉴定出3 个响应NaCl 胁迫的TaERF,分别是TaERF35、TaERF55和TaERF64,为新的耐盐相关基因。TaERF55 和TaERF64 在CH7034 中受盐处理后表达量上调,在SY95-71 中则无显著变化,这与转录组分析结果一致。TaERF35 是水稻OsERF922 的同源基因,其受盐胁迫诱导后上调,而OsERF922 过表达的植株对盐的耐受性降低[6],因此初步猜测TaERF35 在植株抵御盐胁迫过程中可能发挥负调控作用,但需要进一步试验证实。
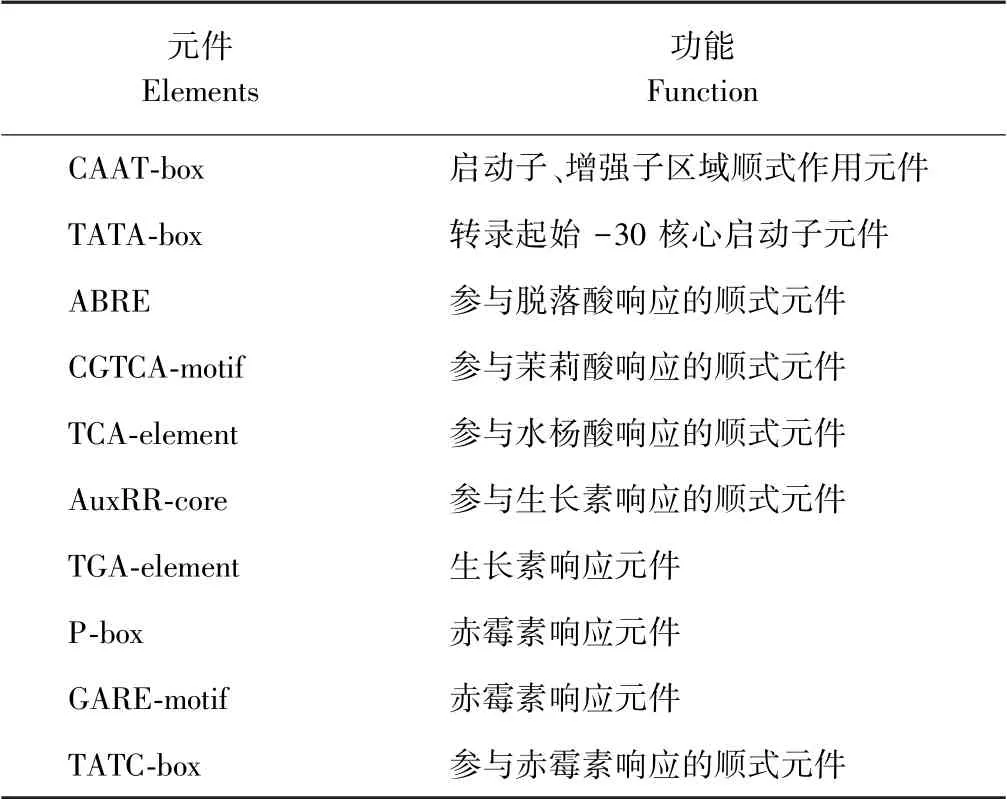
表3 启动子元件及其功能Table 3 The function of the promoter element
本研究初步鉴定的4 个盐胁迫响应相关成员TaERF27、TaERF35、TaERF55、TaERF64 在耐盐材料CH7034 苗期根和叶中均具有较高的表达水平,此外4个成员的启动子区域(-2 000 bp),除包含TATA-box、CAAT-box 元件外,还含有脱落酸、水杨酸、茉莉酸、生长素和赤霉素等多种植物激素响应元件,这几种植物激素在植物抗旱[31]、抗寒、耐盐[32]以及应答逆境胁迫信号转导[33-34]等过程中发挥着重要作用,推测这些成员可能还参与植物多种非生物胁迫信号转导通路。在本研究基础上,下一步可对TaERF35、TaERF55 和TaERF64 进行功能验证,并评估其在生产中的利用价值。
4 结论
本研究从小麦全基因组中分离了96 个TaERF成员,在A、B、D 基因组中共有229 个拷贝序列;利用RT-qPCR 证 实TaERF27、TaERF35、TaERF55 和TaERF64 在小麦耐盐材料CH7034 中受NaCl 胁迫后显著上调,而在盐敏感品种SY95-71 中无明显变化,推测它们可能为盐胁迫响应基因,其中TaERF27 是已报道的小麦耐盐基因TaERF3。这4 个盐胁迫响应相关基因在CH7034 苗期根和叶中均具有较高的表达水平,此外其启动子区域(-2 000 bp)含有脱落酸、水杨酸、茉莉酸、生长素和赤霉素等多种植物激素响应元件。本研究为培育小麦耐盐品种和探索耐盐分子机制提供了参考信息。
