基于功能磁共振成像的人脑效应连接网络识别方法综述
2021-04-18冀俊忠邹爱笑刘金铎
冀俊忠 邹爱笑 刘金铎
伴随美国、欧盟、日本等国近年来人脑计划的发布和启动,脑科学的研究得到了越来越多的重视,许 多国家已经将脑科学的研究上升至国家战略的高度.为理解大脑认知的运行机制以及洞悉脑疾病的发病机理,人脑的研究也必须从各个脑区及其相互作用构成的脑网络来分析和把握大脑的整体性[1],因此人脑连接组成为继人类基因组、蛋白质组后生命科学乃至自然科学领域中的一项前沿课题.目前已有的研究发现许多神经精神疾病与脑结构和脑功能网络的异常拓扑变化有关,所以人脑连接组研究不仅能为神经精神疾病病理机制的理解提供新视角,而且能为疾病的早期诊断和治疗评价提供新的脑网络影像学标记[2].
脑功能网络主要包括功能连接网络和效应连接网络.脑功能连接网络是一种由节点和无向边构成的图模型,其中节点表示脑区,无向边表示空间上相互分离的脑区之间神经活动的统计依赖关系[3].虽然功能连接网络描述了脑区间的统计相关性,但它无法揭示脑区间神经活动的因果效应.事实上,识别脑区间的因果效应为理解人脑的神经活动机制提供了更加丰富的信息[3−4],并且在一些脑疾病的诊断和预测上依据脑区间的因果效应比统计相关性获得了更好的效果[5−6].
脑效应连接网络是一种由节点和有向边构成的图模型,其中节点代表脑区,有向边刻画了一个脑区施加于另一个脑区神经活动的因果效应[7],而与边相关的连接参数则表示边的连接强度[8−9].由于脑效应连接网络的识别是评价正常脑功能和多种脑疾病(如阿尔兹海默病(Alzheimer' s disease,AD)、帕金森病(Parkinson' s disease,PD)、精神分裂症(Schizophrenia)、抑郁症(Depression)和自闭症谱系障碍(Autism spectrum disorder,ASD)等)相关损伤的有效手段,故成为人脑连接组研究中的一项极为关键的科学问题.目前,利用计算方法从人脑功能磁共振成像(Functional magnetic resonance imaging,fMRI)数据中进行脑效应连接网络的识别已成为该项研究中的前沿热点.这是因为如果能够通过对fMRI 数据的学习准确地获得脑效应连接网络,将对于理解脑疾病的发病机理,进行脑疾病的早期诊断以及病理的研究具有重要的意义[2,10−11].具体来说,对脑区间因果效应连接的准确识别,有益于了解大脑的工作机制,加深对人脑功能复杂性的理解[10];而生命科学的研究表明人脑效应连接的变化通常先于人体异常行为症状的显现,所以通过脑效应连接模式异常的发现可为一些脑疾病的早期诊断提供新线索[11].由于脑效应连接网络识别和应用的研究具有重要的理论意义和应用价值,所以该研究课题吸引了来自生物、医学、心理、认知和计算机科学等众多领域研究者的广泛兴趣,掀起了一股跨学科的研究热潮.近年来,新的识别方法层出不穷,新的应用不断被拓展.Science[12−16],Nature[1,17],Proceedings of the National Academy of Sciences of theUnited Statesof America (PNAS)[18−21],Brain[22−23],IEEE Transactions on Pattern Analysis and Machine Intelligence[24−26]等不同领域的权威国际杂志以及数据挖掘领域权威国际会议ACM SIGKDD[27]多次报道这方面的研究工作,极大地推动了脑认知和脑科学的快速发展.
在以上研究背景下,本文对基于功能磁共振成像的脑效应连接网络识别方法的研究现状进行了综述,并结合该领域目前所面临的挑战性问题,对未来可能的前沿方向进行了展望.论文试图为脑效应连接网络的识别勾画出一个较为全面和清晰的概貌,以期对该领域的相关研究提供有益的参考.
1 脑效应连接网络的识别过程
利用fMRI 数据进行人脑效应连接网络识别的过程通常包括5个步骤:数据采集、数据预处理、感兴趣区域划分、时间序列数据获取、网络学习和评价,这与获取脑功能连接网络的主要流程大致相同,两者的主要区别在于对网络结构的学习,脑功能连接网络是学习脑区间的统计相关性,而脑效应连接网络旨在学习脑区间的因果效应.图1展示了人脑效应连接网络识别的主要流程,下面将对此过程进行详细地介绍.
1)数据采集
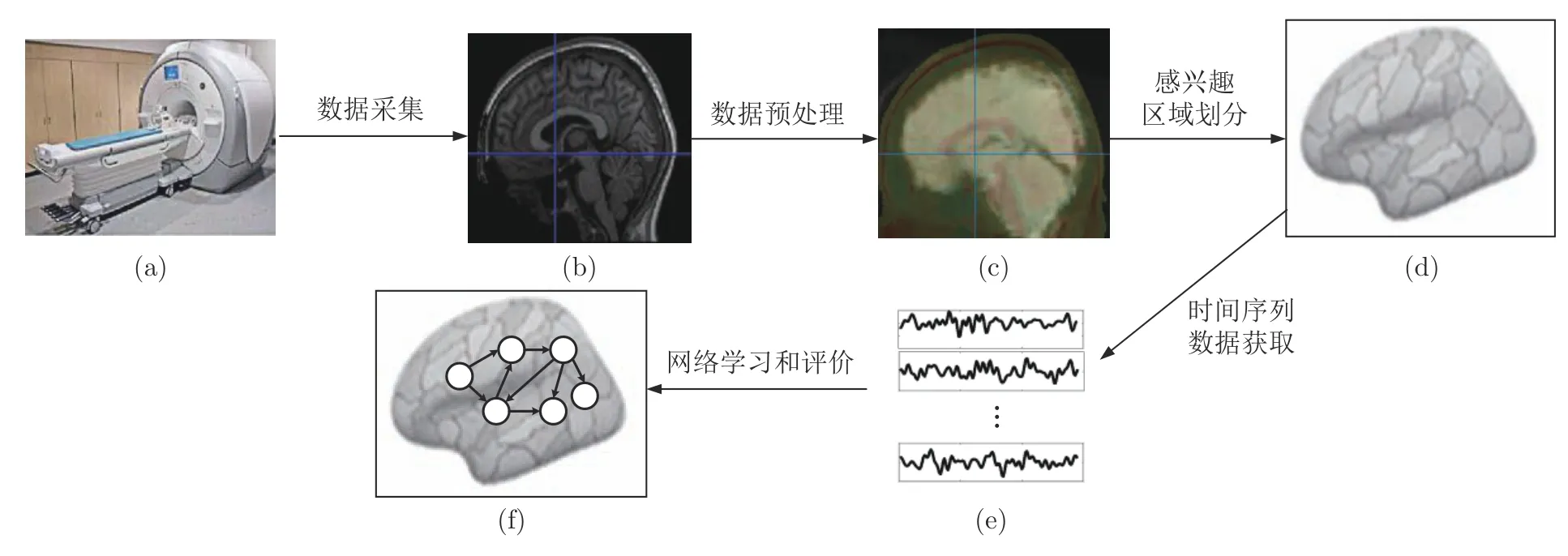
图1 人脑效应连接网络识别的流程Fig.1 The process of human brain effective connectivity networks identification
功能磁共振成像是一种通过检测血氧水平的BOLD信号变化来捕捉大脑神经活动的磁共振成像技术.具体来说,大脑某区域的激活引发局部耗氧量增加,促使该区域的血流量加快以及时补充所需的氧气,造成血液中的脱氧血红蛋白和氧合血红蛋白的浓度发生变化,导致该区域的磁共振信号明显强于其他区域,这一现象被称为血氧水平依赖(Blood oxygen level dependent,BOLD)效应[28].fMRI 数据采集正是利用磁共振扫描仪(如图1(a)[29]所示)以一定的时间分辨率对BOLD信号进行采样,从而获得能够反映人脑神经活动的时间序列数据(如图1(b)所示).所采集的f MRI 数据通常分为两大类:任务态fMRI 数据和静息态fMRI数据.采集任务态fMRI数据需要设计具有特定任务的实验范式,以考察大脑在实验任务刺激下激活脑区的神经元活动;而静息态fMRI数据的采集过程无需试验任务的刺激,它是在被试不做任何刻意的运动或思考,并保持意识清醒情况下的功能磁共振成像.
2)数据预处理
由于采集fMRI数据易受到成像设备以及扫描过程中被试头动、呼吸和心跳等不可控因素的干扰,因此需要对所采集的数据进行预处理,此过程与功能连接网络的数据预处理过程基本一致.目前,常见的f MRI 数据的预处理工具包括:AFNI[30]、ANTs[31]、FSL[32]、SPM[33]、REST[34]、DPABI[35]和最近提出的fMRIPrep等[36],它们的设计机理和功能特点各有不同,可根据实际情况合理地选择和使用.对于静息态和任务态的fMRI数据来说,两者的预处理步骤均包括:时间层校正、头动校正、结构像与功能像配准、空间标准化和平滑处理.此外,静息态f MRI数据的预处理还包括去线性漂移和低频带通滤波;而在任务态fMRI的预处理过程中,通常利用统计分析的方法来检验任务态与对照状态下脑区信号的显著差异性,从而获得在特定实验任务下的激活脑区.预处理后的f MRI数据(如图1(c)所示)被进一步用于脑效应连接网络的识别.
3)感兴趣区域划分
由于fMRI成像的空间分辨率很高,它将人脑划分成数以万计的体素,并针对每个体素采集时间序列数据,因此所采集的fMRI数据具有高维性的特点.为了降低计算复杂度,通常需要对人脑进行感兴趣区域的划分,并将划分后的感兴趣区域作为脑效应连接网络的节点(如图1(d)所示),这是识别脑效应连接网络的关键步骤之一.
一般来说,脑效应连接网络可以从微观尺度、中间尺度和大尺度3个水平上进行识别,但依靠现有的技术从微观尺度和中间尺度来识别脑效应连接网络十分困难.目前,对脑效应连接网络的研究主要集中在大尺度水平上,通过划分感兴趣区域来定义脑效应连接网络中的节点.常见的划分方法主要包括3种:a)对于任务态fMRI 数据,通常依据特定的实验任务,将激活的脑区定义为感兴趣区域[37];b)在分析静息态fMRI 数据时,可以利用独立成分分析方法,从混合信号中分解出多个空间上相互独立的特定的脑网络,并将每一个脑网络定义为一个节点,通过探究各节点间的因果效应连接来揭示静息态下不同脑网络间的信息交互方式[38−39];c)采用脑模板将全脑划分成若干个脑区,进而识别大规模的效应连接网络.目前,常用的脑模板包括:自动解剖标记模板(Automated anatomical labeling,AAL)[40]、神经成像实验室概率模板(Laboratory of neuroimaging probabilistic brain atlas,LPBA)[41]、蒋田仔研究团队提出的包含246个精细脑区亚区的脑网络连接组[42]以及Glasser组绘制的包含360个脑区的模板[43]等.由于合理地划分感兴趣区域对于研究大脑各区域的功能活动,识别高质量的脑效应连接网络非常重要,因此在划分感兴趣区域时需要根据具体的研究内容选择最佳的划分方法.
4)时间序列数据获取
由于每个感兴趣区域内包含大量的体素,而各体素的fMRI数据存在差异,因此通常采用以下两种方式获取每个感兴趣区域的时间序列:a)提取感兴趣区域内所有体素的fMRI数据的平均值.例如,在贝叶斯网络评分搜索中主要运用此方法来提取时间序列[44−45];b)采用主成分分析方法提取感兴趣区域内所有体素fMRI 数据的主成分.该方法已在动态因果模型、格兰杰因果等方法的数据获取中得到了广泛应用[46−47].在此步骤所提取的时间序列数据(如图1(e)所示)将被用于效应连接网络的学习和评价.
5)网络学习和评价
网络学习和评价是整个脑效应连接网络识别过程中最核心的步骤,该步骤基于各脑区的fMRI数据设计合理的识别方法来学习脑区间神经活动的因果效应,并对所得到的效应连接网络(如图1(f)所示)进行评价.一般来说,通常先在具有标准网络的仿真数据集(例如Smith、Sanchez[8,48]等)上识别效应连接网络,并依据精度、召回率和F度量等评价指标来验证方法的有效性,再利用真实数据集探究方法的实际应用效果.
经过近十年的快速发展,已经涌现出了许多基于不同识别机理的脑效应连接网络识别方法,下面将主要围绕识别方法进行讨论,分析和比较现有识别方法的基本原理、特点、不足以及有待解决的问题.
2 脑效应连接网络的识别方法
根据所采用的计算模型和机理,我们可以将脑效应连接网络划分为基于全局评价的识别方法和基于局部度量的识别方法两大类.基于全局评价的识别方法又可进一步分为基于假设模型的识别方法和基于贝叶斯网络评分搜索的识别方法,而基于局部度量的识别方法可细分为基于时间滞后的识别方法和基于概率分布的识别方法.更系统的脑效应连接网络识别方法的分类体系如图2所示,在每个子类方法下又包含多种不同的具体算法.
2.1 基于全局评价的识别方法
基于全局评价的识别方法是同时考虑所有脑区之间的相互作用关系,通过对模型的整体评价来识别脑区间的效应连接.根据识别方法的性质,基于全局评价的识别方法又可分为2类:1)基于假设模型的识别方法,此类方法依据所构建的假设模型来间接估计神经活动信号间的因果效应;2)基于贝叶斯网评分搜索的识别方法,此类方法采用无监督的数据驱动方式直接从脑区fMRI 数据中学习脑效应连接网络.
2.1.1 基于假设模型的识别方法
此类方法首先利用模型来描述脑区间神经活动的因果效应,然后估计模型参数得到多个相互竞争的假设模型,最终通过模型选择,从假设模型中获得一个具有最大化模型证据的模型.目前,基于假设模型识别脑效应连接的方法主要包括3种:基于结构方程模型(Structural equation models,SEM)的识别方法、基于动态因果模型(Dynamic causal models,DCM)的识别方法和基于Ornstein-Uhlenbeck模型(Ornstein-Uhlenbeck models,OUM)的识别方法.
1)基于结构方程模型的识别方法
SEM 是一种从计量经济学领域发展而来的多元线性统计模型,它基于变量间的协方差矩阵来间接地估计变量间的因果关系.1994年,Mclntosh 等首次将SEM 用于人脑功能网络的识别[49],为基于SEM识别脑效应连接网络提供了新思路.下面将对基于SEM识别脑效应连接网络的主要过程进行描述:
首先,令xi表示第i个脑区变量,t表示采样时间,且t∈[1,···,T],n为脑区个数,xt=[x1t,···,xnt]表示所有脑区的神经元活动在t时刻的观测值.利用一个概率生成模型来描述脑区中神经元活动与效应连接的关系,其计算公式如下:


图2 人脑效应连接网络识别方法的分类体系Fig.2 The category system for identification methods of human brain effective connectivity networks
式中,θ表示模型参数;xt服从均值为零、协方差为Σ(θ)的高斯分布.然后通过一个多元回归方程模型表示脑区间的相互作用关系,如式(2)所示:
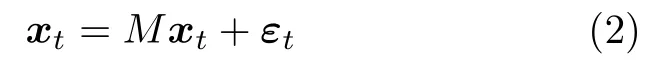
式中,M表示脑效应连接网络的参数矩阵,εt=[ε1t,···,εnt]表示脑区噪声.向式(2)中引入一个单位矩阵In,则xt可表示为如下形式:
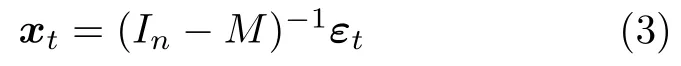
再利用模型的协方差矩阵与脑区观测数据的协方差矩阵构建目标函数,如式(4)所示:
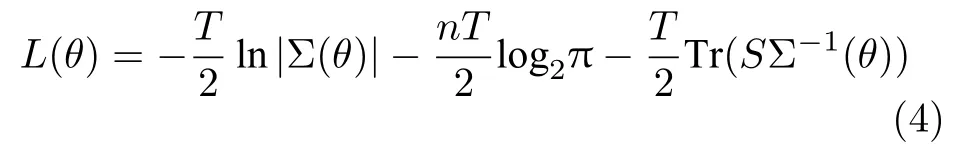
由于SEM具有模型简单、鲁棒性和灵活性强等优势,故已成为识别脑效应连接网络的常用方法之一.但该方法仍存在一定的不足[50−52]:1)在缺乏先验知识时,难以准确地识别脑区间的因果关系;2)仅能用于描述脑区神经活动信号间的同期因果效应,无法有效地利用f MRI 数据的时序信息;3)SEM假设脑区间的因果关系是线性的,无法揭示脑区间非线性的因果关系;4)将外部输入视为不确定性变量,故不适用于建模与事件相关的脑区间的效应连接.近年来,已有许多研究者针对SEM的缺陷,对SEM的基本识别过程进行了改进,设计了多种基于结构方程模型识别脑效应连接网络的新方法.
2006年,Shimizu 等在结构方程模型中引入了三个基本假设:1)观测数据的生成过程是线性的;2)不存在隐变量;3)噪声服从非高斯分布且是相互独立的连续随机变量,故将验证性的结构方程模型改进为一种探索性的数据驱动模型—LiNGAM(Linear non-gaussian acyclic model)[53],该模型不需要预先获悉脑区间因果关系的先验信息,仅利用基于独立成分分析的方法就能从观测数据中学习脑区间的效应连接.但LiNGAM存在对初始值敏感而易陷入局部最优的问题.为了克服这一不足,他们随后又提出了一种非参数的(Direct linear nongaussian acyclic model,Direct LiNGAM)算法[54],该算法通过从给定的数据中依次消除各独立成分的影响来估计脑区间的因果结构,并利用剪枝算法完善整个脑效应连接网络的识别.与LiNGAM相比,新算法不需要进行初始猜测,能够在有限步骤内较快地搜索到最优解.
在文献[55]中,Kim等考虑到fMRI 数据的时序特性,将结构方程模型与向量自回归模型相结合,设计了一种能同时刻画人脑神经活动的同期和滞后因果效应的统一结构方程模型(Unified SEM,uSEM),并将uSEM与广义线性模型融合于统一框架,以探究被试的年龄、性别等协变量对效应连接的影响.由于u SEM将外界刺激视为不确性变量,故无法识别与事件相关的脑区效应连接网络.为了克服此不足,Gates等受DCM的启发,在uSEM中引入一个血液动力学响应函数,使其与外部刺激信号做卷积,进而构建一个离散的双线性模型—eu-SEM(Extended unified SEM)[56]来描述外部刺激对脑区的影响以及脑区间效应连接的变化.与uSEM相比,euSEM不仅能够刻画脑区间同期和滞后的因果效应,而且可以描述人脑在受到外部刺激时的功能整合.此外,euSEM是结构方程模型从验证性方法向探索性方法发展的一个成功范例,它能够根据具体的研究内容,自适应地选择以验证性或数据驱动的方式对脑区间的因果关系建模,从而为基于fMRI数据识别脑效应连接网络提供了一种十分灵活的方法.
目前,在组水平上识别效应连接网络通常是对所有被试的脑结构和功能数据做同质性假设,但事实上不同被试的脑部存在显著差异,其脑结构和功能数据均具有异质性,这导致所获得的脑效应连接网络缺乏可靠性[57−58].为了解决此问题,Gates等在2012年提出了GIMME(Group iterative multiple model estimation)算法[58],这为从异质数据中识别组水平和个体水平的脑效应连接网络提供了一个有效且可靠的方法.该方法主要分为两个阶段:第1个阶段利用所有被试的fMRI数据建立euSEM,并采用极大似然估计确定模型参数,然后依据修正指数选择适合于多数个体的参数来获取组水平的公共因果网络;第2个阶段是在获取公共因果网络基础上识别每个被试的效应连接网络,以半限定的方式建立个体水平的euSEM,然后使用阶段1中的方法估计参数,并选择适合于个体且同时存在于公共因果网络中的参数,从而得到个体水平的脑效应连接网络.2019年,Henry 等对GIMME算法做了进一步扩展,提出了cs-GIMME(Confirmatory subgrouping GIMME)算法[59],该算法通过对子组进行预定义,并检测预定义子组中大多数个体一致存在的连接,有效地提高了识别精度,使算法在识别组间差异上获得了更好的效果.
2)基于动态因果模型的识别方法
DCM是一种根据脑区神经元之间的耦合关系而设计的动力学模型,最早用于识别与事件相关的脑区间的效应连接网络.首先,令xi表示第i个脑区变量,x=[x1,···,xn]表示所有脑区的神经元活动状态.利用一个非线性函数F来描述输入信号对脑区神经元活动的影响,如式(5)所示:

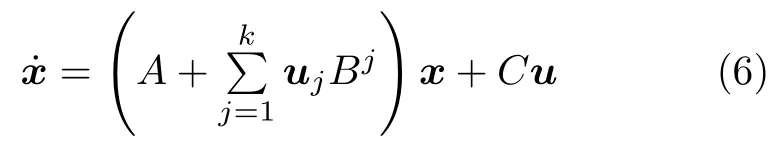
由于脑区中神经元活动将引发附近区域的血液动力学状态发生变化,而这一动态过程可通过血液动力学模型来描述,该模型先利用一组微分方程来描述血液动力学状态的变化趋势,再通过一个非线性函数将血液动力学状态映射为BOLD 响应信号[60].对于任一脑区,其血液动力学状态在某个采样时刻的变化趋势如式(7)所示:
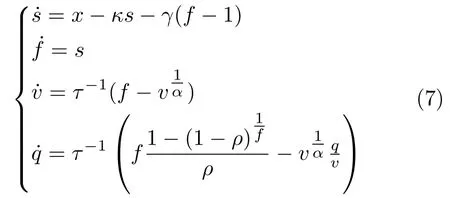

在一段采样时间内,人脑某个脑区的神经生理学响应信号映射为BOLD响应信号的过程如图3所示.由图可知,在刺激信号的作用下,人脑中某个脑区被激活,该脑区的神经元活动会引发血管舒张信号增强,导致局部血液流入量增加,促使血液流出量、血容积和脱氧血红蛋白含量均发生相应的改变,其中,血液流出量对血液流入量具有抑制作用,并且对脱氧血红蛋白含量造成影响;而血容积的大小与血液流出量和脱氧血红蛋白含量密切相关.最终在血容积和脱氧血红蛋白的共同作用下生成BOLD响应信号.
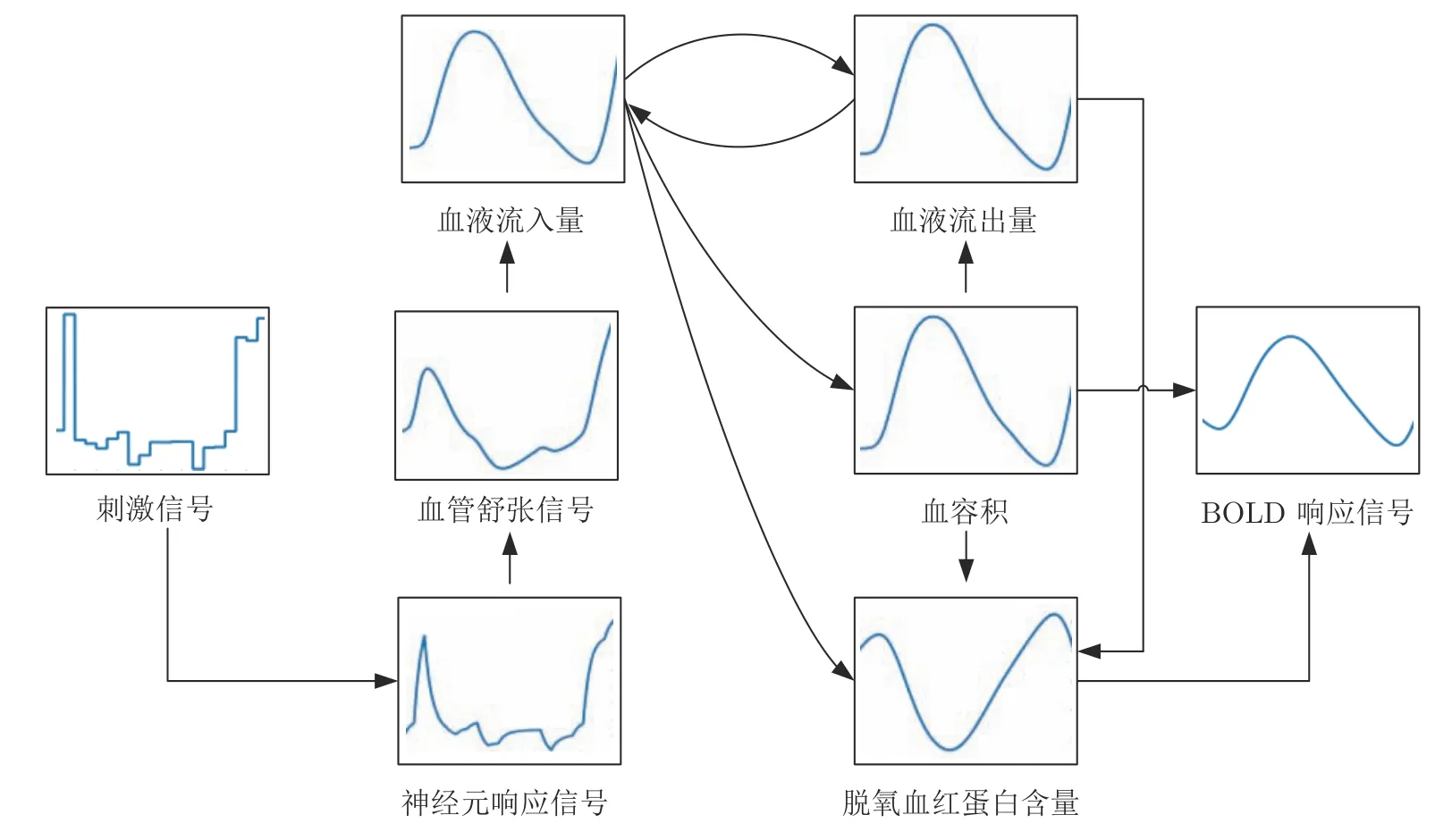
图3 神经生理学响应信号映射为BOLD响应信号的过程Fig.3 The process of mapping a neurophysiological response signal to a BOLD response signal
最后,通常利用动态期望最大化或者广义滤波的方法来估计模型参数,并通过贝叶斯模型选择,获得一个具有最大化模型证据的模型,从而得到脑效应连接网络.DCM的优势在于它通过神经生理学响应所引发的BOLD响应信号来识别脑区间的因果效应,因此具有良好的生理学可解释性.
事实上,DCM不仅可以识别与事件相关的脑效应连接网络,而且能够揭示大脑内部自发性的神经活动规律[61].2014年,Friston等将时域DCM转换到频域,提出一种利用频谱动态因果模型(Spectral dynamic causal models,sp DCM)从静息态fMRI数据中学习脑效应连接网络的方法[62],该方法利用交叉谱函数对脑区神经元的内源性波动和噪声进行编码,通过对交叉谱函数的参数估计来识别脑区神经活动信号间的因果效应,巧妙地解决了估计神经元活动的复杂问题.实验结果表明,该方法具有识别精度好、计算效率高、对组间差异敏感等优点[62−63].但该算法是利用sp DCM对静息态fMRI数据识别脑效应连接网络的初步探索,仍存在2个主要问题尚未解决:1)sp DCM仅识别了较小规模的效应连接网络,如何进一步提高算法的计算效率,使其在识别大规模效应连接网络中发挥作用,是后续研究中亟待解决的问题;2)sp DCM依赖于平稳性假设,忽略了效应连接的动态性.如何利用sp DCM来探究效应连接的动态性是未来需要解决的另一问题.
伴随着脑区个数增多,脑效应连接参数会激增[64],这将导致模型的计算复杂度显著增加,所以动态因果模型常用于识别小规模的脑效应连接网络[65].但实际上这些连接参数中包含了大量的冗余信息,因此设计合理的方法约减冗余连接参数将有利于此类方法识别更大规模的脑效应连接网络.2017年,Razi等提出了一种将功能连接矩阵作为收缩先验来约减冗余连接的算法[66].该算法首先利用奇异值分解从fMRI数据中提取特征向量,并通过贝叶斯估计确定特征维数,然后提取特征向量间的功能连接矩阵来构建效应连接参数的收缩先验,从而剔除了冗余参数,提高了模型反演的效率.该算法是基于sp DCM识别较大规模的脑效应连网络的一个良好的范例,但其不足之处在于算法对特征向量的维度比较敏感.
同年,Frassle等提出了一种回归动态因果模型(Regression dynamic causal models,rDCM)算法[67],该算法将复杂的非线性反演问题转化为一个易求解的贝叶斯线性回归模型来解决.具体来说,首先利用傅里叶变换将时域的神经元状态方程转换到频域,并与一个固定的血液动力学响应函数做卷积,再引入平均场近似假设脑区信号的测量噪声是独立随机向量,然后为效应连接参数和噪声精度构造先验分布,将rDCM形式化为一个易估计的贝叶斯线性回归模型,并利用变分贝叶斯估计模型的参数和超参数.与传统的DCM算法相比,r DCM有效地解决了大规模脑效应连接网络参数估计困难的问题,显著提高了反演效率.文献[68]对r DCM做了进一步的扩展,提出了一种利用稀疏约束自动约减冗余连接的稀疏回归动态因果模型(Sparse regression dynamic causal models,sparse r DCM)算法.该算法首先利用一个全连接网络描述脑区间的相互作用,然后向贝叶斯线性回归模型中引入一个二进制指示变量作为特征选择器,使其在反演过程中将全连接网络自动优化为稀疏网络,进一步提高了算法的效率.实验结果表明,该算法不仅能够快速地识别大规模脑效应连接网络,而且具有良好的准确性和特异性.
由于sp DCM依赖于平稳性假设,因此忽略了效应连接随时间变化的动态性.为了弥补此不足,2018年,Park 等设计了一种利用sp DCM识别脑区间动态效应连接的方法[46],该方法首先利用滑动窗口算法对单个被试在一段采样时间内的f MRI数据进行划分,并利用sp DCM为每个窗口内的基线连接和窗口间效应连接的变化建模,然后采用参数经验贝叶斯(Parametric empirical Bayes,PEB)[69−70]的层次模型对窗口内和窗口间的效应连接进行估计,最终通过叠加基线连接和线性模型的随机效应来识别脑效应连接网络的动态变化.由于该方法具有良好的泛化能力,因此可将其进行扩展到识别组水平上的动态效应连接网络.
3)基于Ornstein-Uhlenbeck模型的识别方法
OUM是一种描述随机过程的模型,它能够表示大脑的神经元活动状态,并通过捕捉脑区间的功能连接来间接地刻画神经元活动的因果效应.基于OUM识别效应连接网络的主要过程为:首先利用大脑的结构数据构建结构连接网络,将其作为效应连接网络的基本骨架;然后利用OUM建立人脑的神经活动与效应连接的关系,并通过估计模型参数(脑区间的协方差矩阵和效应连接的邻接矩阵),获得一个与脑区fMRI 数据的功能连接网络拟合程度最佳的模型.图4给出了基于OUM方法识别脑效应连接网络的主要流程.在图4中,ECij表示任意两个脑区之间的效应连接,Σij表示脑区间的协方差矩阵.
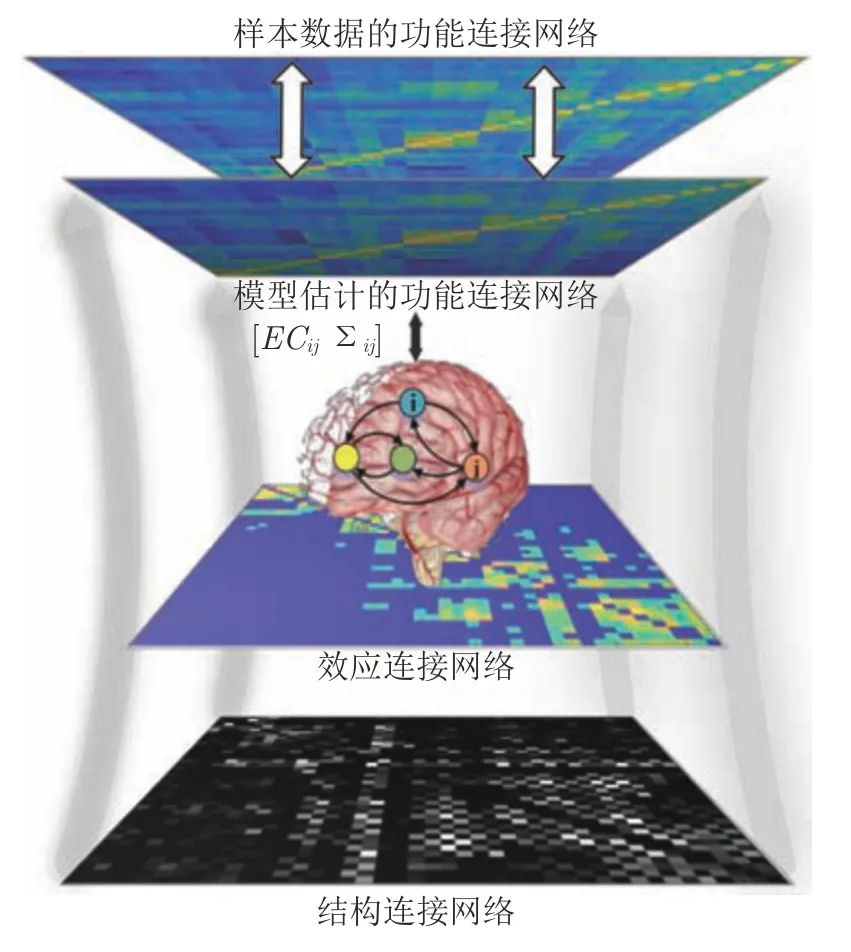
图4 基于OUM方法识别脑效应连接网络的流程图Fig.4 The process of identifying a brain effective connectivity network by the OUM method
早期对脑功能连接的研究主要基于静态的描述,最近一些研究者围绕脑功能连接的动态特性展开研究,研究结果表明动态功能连接不仅包含了大脑丰富的时空信息,而且与大脑内部自发性的神经活动密切相关,这为利用脑功能连接来识别脑效应连接网络提供了重要依据.2016年,Gilson等设计了一种利用OUM从静息态fMRI数据中估计全脑效应连接网络的方法[71],该方法首先依据结构连接网络构建效应连接网络的基本骨架,然后利用OUM捕捉脑区间当前时刻和滞后时刻的功能连接,并采用一个Lyapunov函数迭代地优化效应连接参数和脑区间的协方差矩阵,使由模型计算得到的BOLD信号间的功能连接网络与实际fMRI 数据间的功能连接网络达到最佳的拟合程度,最终通过估计模型参数来识别脑效应连接网络.随后,他们又基于此方法来识别与事件相关的脑效应连接网络[72],通过设计观看电影与黑屏休息的交替任务的实验范式来探究被试处于两种状态时脑效应连接网络的差异.该方法为识别大规模脑区间的效应连接网络提供了一种新思路,但它能准确识别脑效应连接网络的条件是:功能连接的滞后时间需要与动态系统的时间常数相匹配.一旦不能满足此条件,将难以获得高质量的脑效应连接网络.
2018年,Schiefer等提出了一种通过OUM来识别脑区间因果效应的方法[73],该方法基于“对撞”结构[74]的思想,即,若两个脑区x和y之间的因果关系由另一个变量z决定,则x和y具有相关性,此时根据x和y的相关性可以估计脑区间的效应连接.具体来说,该方法首先将外部输入作为驱动变量,通过一个Ornstein-Uhlenbeck 模型来刻画脑区间神经活动的因果效应,并利用观测数据和外部输入信号的零时滞的交叉谱密度来表示脑区间的效应连接,然后对交叉谱密度进行酉变换并构建带有L1惩罚项的代价函数,最终通过梯度下降算法最小化代价函数,从而实现对效应连接的参数估计.该方法基于零时滞的交叉谱密度函数识别效应连接网络,巧妙地避免了使用滞后信息对脑区间因果关系所造成的不良影响,但是该方法易受给定条件的限制,且存在对噪声敏感的缺陷.
虽然基于假设模型的方法在识别脑效应连接网络方面发挥了重要作用,但此类方法仍存在以下几个方面的不足:首先,此类方法在识别过程中通常依赖于先验知识,当先验知识不足时容易对识别结果造成不良影响;然后,基于结构方程模型和基于Ornstein-Uhlenbeck 模型的方法均未充分考虑脑区神经活动所导致的血液动力学状态变化,故缺乏生理学的可解释性;最后,尽管基于假设模型的方法在利用f MRI数据的时序特征方面开展了一些探索,但仍处于初步阶段,如何针对fMRI 数据的时序特征设计更合理的方法将是此类方法未来的一个重要发展方向.
2.1.2 基于贝叶斯网络评分搜索的识别方法
贝叶斯网络(Bayesian networks,BN)是一种由节点和有向弧构成的概率图模型,其节点表示变量,有向弧对应变量间的因果关系,所有变量间的条件概率关系统一用一张有向无环图表现出来.基于BN评分搜索识别脑效应连接网络通常被视为一个模型选择问题,其目的是通过BN结构学习搜索与脑区变量间的条件依赖关系拟合程度最佳的网络结构,并在确定网络结构的前提下计算脑区间的连接强度.由此可见,准确地学习脑区间的因果结构将是基于贝叶斯网络评分搜索方法识别效应连接网络的关键.具体来说,此类方法将学习视为最优化问题,利用评分搜索方法在候选网络结构空间中进行迭代搜索,并将得分最高的网络结构作为学习到的脑效应连接网络结构.基于贝叶斯网络评分搜索方法识别效应连接网络结构的流程图如图5所示.
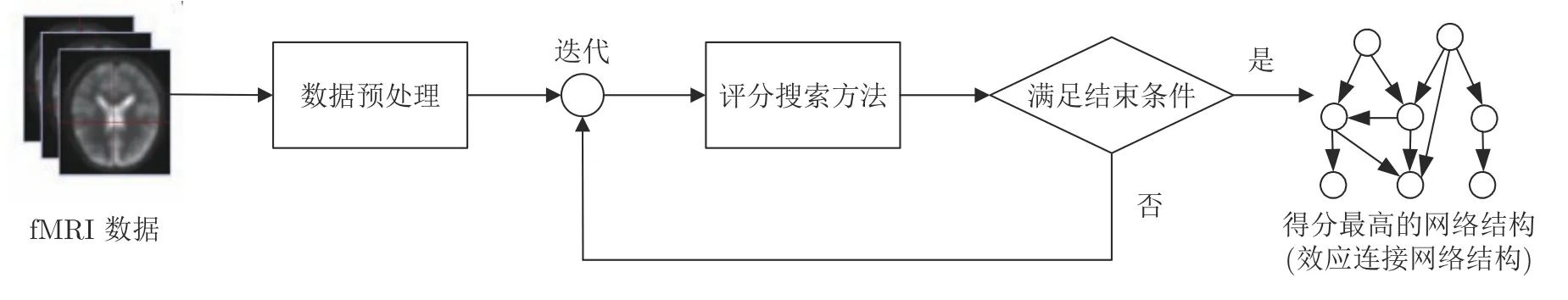
图5 基于贝叶斯网络评分搜索方法识别脑效应连接网络结构的流程图Fig.5 The process of identifying a brain effective connectivity network structure by the Bayesian network scoring search method
根据是否利用fMRI 数据的时序特征识别脑效应连接网络,基于贝叶斯网络评分搜索识别脑效应连接的方法可分为两类:一类是基于静态贝叶斯网络评分搜索的识别方法[75−76],此类方法将所有采样时刻的fMRI数据视为一个整体向量,利用基于评分搜索的贝叶斯网络结构学习方法来识别脑效应连接网络;另一类是基于动态贝叶斯网络(Dynamic Bayesian networks,DBN)评分搜索的识别方法[77−78],DBN是静态贝叶斯网络在时序过程建模方面的扩展,它充分考虑了fMRI数据的时序特性,不仅能识别在同一时间片上脑区间的因果关系,而且也可以分析不同时间片的脑区间时域的相关性,已成为一种揭示脑效应连接网络时序动态性的重要方法.这两类方法在识别脑效应连接网络中均发挥了重要作用.由于贝叶斯网络具有良好的理论基础和丰富的实现方法,故基于贝叶斯网络评分搜索方法识别脑效应连接网络已逐渐成为目前该领域的一个研究热点.
1)基于静态贝叶斯网络评分搜索的识别方法
2016年,Ji 等将人工免疫算法(Artificial immune algorithm,AIA)与BN算法相结合,提出一种识别脑效应连接网络的AIAEC(Artificial immune algorithm effective connectivity)算法[44],该算法首先构建了一个由记忆抗体和随机生成抗体组成的初始解,然后通过克隆选择、交叉和变异算子优化当前种群中的抗体,并利用抑制算子来更新种群,重复这个迭代过程直到搜索到K 2评分最高的抗体,即得到最优的脑效应连接网络.由于AIAEC算法具有快速的局部寻优能力和良好的全局搜索能力,因此在识别精度和效率上均获得了良好的性能.但该算法不能定量地描述效应连接强度.为了弥补此缺陷,他们随后又提出一种根据蚁群寻优机制实现的基于评分搜索的脑效应连接网络学习算法ACOEC(Ant colony optimization effective connectivity)[45],该算法利用K2评分度量来引导蚁群在可行解空间中搜索到全局最优解(脑效应连接网络).其主要过程为:每只蚂蚁将空图作为初始解,通过从候选弧集中选择有向弧增量地构建个体可行解,重复此操作直到评分不再增加时获得该蚂蚁的当前解,当蚁群全部迭代优化完成后所获得的K2评分最高的解为最终的脑效应连接网络.该算法的特点是通过更新信息素浓度来指导蚁群搜索,有效地提高了搜索效率,且其在迭代过程中定期调用增弧、减弧和反向弧的优化策略能避免算法陷入局部最优值.与AIAEC相比,ACOEC不仅在连接方向的识别上具有更高的精度,而且利用信息素浓度定量地描述了脑区之间的连接强度.但该算法的不足在于求解效率较低,尤其在面对大规模脑区时,算法的时间复杂度较高.
为了进一步提高算法的性能,2019年,冀俊忠等又提出一种融合多源信息的蚁群优化学习算法[79],该算法首先利用结构与功能的内在约束关系,将从弥散张量成像(Diffusion tensor imaging,DTI)数据中获取的皮尔森正相关作为结构约束知识来压缩搜索空间,既有效地提高了蚁群的搜索效率,又保证了最优解的所有组件(弧)都能作为蚂蚁选择的候选弧;然后通过在启发函数中融合基于fMRI数据的体素联合激活信息来增强蚂蚁寻优的目的性,以提高算法的求解速度;最终在两种优化策略的共同作用下获取高质量的网络结构.与ACOEC算法相比,新算法在优化效率和鲁棒性上都获得了更好的效果.
2)基于动态贝叶斯网络评分搜索的识别方法
2014 年,Wu 等提出一种采用高斯动态贝叶斯网络(Gaussian dynamic Bayesian network,GDBN)识别脑效应连接网络的方法[80],该方法首先将大脑视为一个一阶线性动态系统,并假设脑区的fMRI数据服从高斯分布,然后采用基于贝叶斯信息准则的评分搜索方法来确定能最佳表征脑区变量在相邻时间片之间转移概率的网络结构,并通过参数学习来获取脑区间的连接强度,最终得到随时间演化的脑效应连接网络.与之前采用离散动态贝叶斯网络识别脑效应连接网络的工作[78]相比,GDBN不需要对数据进行离散化处理,有效地避免了由此而引发的信息丢失.但该方法仅使用了一阶马尔科夫链DBN模型,并未对跨多个时间片的脑区间的交互作用建模.实际上,不同脑区间的反馈以及调节作用并非同步发生,且其时延长短也存在差异,而采用高阶动态贝叶斯网络模型将是解决这些问题的有效方法.
2017 年,Dang 等提出了一种融合动态规划策略的高阶动态贝叶斯网络结构学习方法[81],该方法能够对异步多时延的脑区间的循环和调节作用建模,并采用动态规划策略压缩搜索空间,以提升算法的求解效率,从而准确、高效地刻画脑效应连接网络的动态变化.为了进一步提高识别精度,他们随后又提出了一种融合fMRI 和DTI 数据于动态贝叶斯网络学习框架的脑效应连接网络识别方法[82],该方法首先对仅具有功能信息的网络评分进行改进,重新构建了包含结构信息的基于解剖连接的(Tractography-based,TB)评分函数,并利用一个全局阈值参数确保评分函数中结构信息的有效性,然后基于TB评分函数学习DBN,最终获得随时间变化的脑效应连接网络.该方法利用DBN模拟了人脑的调节和反馈机制,能够揭示不同时间片的脑区间的交互作用.实验结果表明,该方法在识别精度和鲁棒性上均具有良好的性能.
基于贝叶斯网络评分搜索的识别方法本质上是一种探索性的方法,它采用无监督的数据驱动方式从fMRI 数据中识别脑效应连接网络,具有精度高、鲁棒性和泛化能力强等优势,但此类方法仍存在一定的局限性,由于静态贝叶斯网络只能对时不变系统建模,无法考虑前后时刻信息的关联性和互补性,因此不能有效地利用fMRI数据的时序特性来识别脑区间的效应连接.而尽管动态贝叶斯网络能够刻画不同时刻的脑效应连接网络,实现了对时序过程的建模,但随着DBN模型阶数的增多,其计算复杂度将显著增加.
2.2 基于局部度量的识别方法
基于局部度量的识别方法主要包括两个步骤:1)利用度量计算方法识别局部脑区之间的因果网络;2)合并各局部脑区间的因果网络以获得完整的脑效应连接网络.此类方法又可分为基于时间滞后的识别方法和基于概率分布的识别方法.
2.2.1 基于时间滞后的识别方法
基于时间滞后的识别方法是一类利用fMRI 数据的滞后信息来预测当前时刻脑区之间效应连接的方法,此类方法不仅充分考虑了fMRI数据的时序特性,而且能够直接从脑区观测数据中学习因果效应和连接强度,是一类快速、准确地识别脑效应连接的有效手段.根据计算机理的不同,此类方法又可以分为基于格兰杰因果的识别方法、基于预测相关性的识别方法和基于相空间重构的识别方法.
1)基于格兰杰因果的识别方法
基于格兰杰因果的识别方法是一类依据滞后的时序信息预测当前脑区间效应连接的方法,该方法强调预测性是因果关系存在的必要条件,即在对当前脑区信号进行预测时,若加入其他脑区滞后的fMRI时间序列能获得更准确的预测结果,则称后者是前者的原因[83−84].脑区间的格兰杰因果关系由线性自回归模型进行描述,最初是在时域内利用双变量自回归模型识别每对脑区间的格兰杰因果关系,但此模型无法准确地区分直接和间接的因果关系,容易识别到假阳性的连接.为了解决此问题,基于多变量自回归模型的格兰杰因果分析方法被相继提出,包括条件格兰杰因果[85]和偏格兰杰因果[86]等方法.这些多元格兰杰因果方法不仅能够有效地消除脑区间的间接连接,而且可以避免由外部干扰因素对识别结果造成的影响.
事实上,时域格兰杰因果的频域表示形式可用于检测不同频率点或频段的神经活动信号间的因果关系,从而更精确地刻画脑区间的效应连接,于是,偏有向相干、广义偏有向相干、有向传递函数和全频域有向传递函数[87−89]等频域内的格兰杰因果方法被陆续提出,在识别脑效应连接上发挥了重要作用.
随着格兰杰因果检测技术的不断发展,此类方法已逐步摆脱了仅用于描述脑区间线性因果关系的束缚,核格兰杰因果[90]、传递熵[91]等非线性格兰杰因果方法为揭示脑区间的非线性依赖关系提供了新思路.因此,在时域和频域内,基于格兰杰因果的识别方法能够捕捉脑区间线性和非线性的因果效应,全面地揭示大脑的神经活动规律.
2015年,Ting 等提出一种基于子空间向量自回归模型(Subspace vector autoregressive,SVAR)识别大规模脑效应连接网络的方法[92].首先利用因子模型对高维fMRI数据进行降维,然后在低维子空间建立多变量自回归模型,并采用偏有向相干算法识别脑区间的格兰杰因果关系,最终将识别结果映射到高维状态空间以获得脑效应连接网络.该方法利用少量因子表征高维fMRI数据,降低了算法的计算复杂度,提高了识别效率,能够快速地识别大规模脑区间的效应连接网络.为了进一步探究效应连接的动态变化,他们随后提出了将马尔科夫区制转移模型和SVAR 模型相结合的动态效应连接网络识别方法[93−94],该方法将SVAR 模型表示为状态空间形式,通过卡尔曼滤波和平滑变换捕捉脑区间连接状态的变化,并将状态空间划分成不同区制,基于各区制的脑区间的因果效应来获得脑区间的动态效应连接网络.该方法不受固定窗口尺寸的限制,能跟随时间尺度的变化自适应地划分区制,具有较强的灵活性和实用性.但其不足之处在于因子模型的降维策略有时对一些特殊脑区缺乏适用性.
2016年,Wei等提出一种融合时域和频域多元格兰杰因果方法的识别框架[95],将偏格兰杰因果(Partial granger causality,PGC)算法和偏有向相干(Partial directed coherence,PDC)算法相结合来识别特发性癫痫患者核心认知脑区的效应连接.具体分为2个步骤:a)利用fMRI 数据计算每个被试的PGC和PDC测度,并获取两种方法度量结果的平均值;b)采用自举重采样构建替代数据集,假设替代数据集的PGC和PDC测度均满足零假设,通过显著性阈值法判断原始数据和替代数据的检验统计量之间是否具有显著性差异,并根据PGC和PDC的一致性度量结果识别脑区间的效应连接.该方法结合了时、频域格兰杰因果分析的优势,不仅有效地避免了外部输入对识别结果的影响,而且更精细地反映了不同频率的因果响应.不过该方法存在对模型阶数敏感的缺陷.
2017 年,Karanikolas等设计了一种基于多核格兰杰因果模型识别脑区间效应连接的方法[90],其核心思想是在非线性特征空间内通过多核学习识别脑区间的因果关系.首先依据脑区fMRI 的滞后时间序列构建多元线性回归模型,并将其扩展到再生核希尔伯特空间,利用基于核函数的非线性回归模型进行PGC度量;然后采用多核学习算法构建多个基本核的凸组合,通过L2范数从预先选择的核函数字典中学习与样本数据拟合程度最佳的基本核组合,最终运用偏格兰杰因果算法估计脑区间的效应连接.该方法将线性模型转化到非线性空间,准确地捕捉到脑区间的非线性相互作用关系,并利用灵活性较强的多核学习来提高模型的预测精度,获得了良好的识别效果.
同年,Meier等提出一种将传染病模型和传递熵融合于统一框架来识别个体水平的大脑信息流向的方法[96].首先利用DTI 数据构建脑区间的结构连接网络,然后通过传染病模型定义脑区的状态,描述脑区的激活过程,再使用传递熵计算从一个脑区传递到另一个脑区的信息量,并依据熵值大小判断每对脑区间的信息流向,最终通过计算PA(Posterior-anterior)指数来估计整个大脑皮层的信息流动的全局模式.该方法采用基于信息理论的传递熵方法来估计脑区间的信息交换强度和信息流向,是一种准确、高效的非参数非线性识别方法.
2019年,Chockanathan 等提出了一种大规模格兰杰因果(Large-scale Granger causality,lsGC)方法[97],用于识别大规模脑区间的效应连接网络.该方法首先采用主成分分析对高维fMRI数据进行降维,然后在低维空间中建立多元向量自回归模型,并计算每对脑区间的lsGC指数来判断因果关系,最终将低维空间的识别结果映射到高维空间中,从而得到脑效应连接网络.实验结果表明,该方法在由AAL、Harvard-Oxford 和Brainnetome等多种脑模板所划分的大规模脑区上均获得了良好的识别效果,为识别大规模效应连接网络提供了一种可靠的手段.
基于格兰杰因果的识别方法是一类基于时间优先级识别脑效应连接的方法,一般适用于分析时间分辨率较高的MEG(Magnetoencephalography)、EEG(Electroencephalogram)等数据,例如:最近提出的利用高斯回归过程对因果核函数进行非参数学习的GP Cake 算法[98],以及融合自适应神经网络—模糊推理系统的条件格兰杰因果[99]方法.为了从时间分辨率相对较低的fMRI数据中也能获得较好的格兰杰因果识别结果,通常选取较短的滞后时间和较低的模型阶数,尽管这在一定程度上提高了算法识别的精度和效率,但仍存在一定的局限性:首先,该方法依赖于平稳性假设,当fMRI数据不满足此条件时容易产生假阳性的识别结果;其次,由于fMRI数据的时间分辨率较低,因此不同脑区的血液动力学响应的延迟时间存在差异,这有可能对格兰杰因果关系的识别造成混淆.
2)基于预测相关性的识别方法
2017 年,Xu 等提出了一种基于预测相关性识别效应连接的方法[100],该方法首先设计了一个线性时不变因果模型,然后通过该模型利用一个脑区f MRI时间序列的滞后信息获取另一个脑区f MRI时间序列的估计值,并计算脑区fMRI 时间序列的估计值与实际值间的相关系数.由于相关系数反映了每对脑区间的相互预测能力,因此依据非对称的相关系数可以判断脑区间的因果效应,进而得到完整的脑效应连接网络.实验结果表明,该算法具有灵敏度高、鲁棒性强的特点.与基于格兰杰因果的识别方法相比,该方法的优势在于:a)识别机理有所改进,该方法无需测量脑区间依赖关系的统计量,而是通过比较算法对脑区fMRI数据的预测准确性来识别因果关系,容易获得更高的识别精度;b)该方法不依赖于平稳性假设,在脑区信号为非高斯分布的情况下仍能获得良好的识别结果.
3)基于相空间重构的识别方法
基于相空间重构的识别方法是一种无监督的数据驱动方法,该方法利用脑区的时间序列数据重构奇异吸引子相空间,并通过研究其非线性动力学特征分析脑区间的因果效应.相空间重构方法主要包括两类:延时坐标重构和导数重构.由于数值微分容易产生对误差敏感的问题,因此通常采用延时坐标重构来识别效应连接.具体来说,首先假设每对脑区的fMRI时间序列由同一个确定性的非线性动力系统所决定,然后利用选取的嵌入维数和延迟时间重构两个脑区时间序列的相空间矢量,最后依据相空间矢量识别脑区间的效应连接.
早在2002年,Quian等就提出了一种基于相空间重构来评估神经同步性的广义同步(Generalized synchronization,GS)算法[101],该算法利用延迟坐标重构将脑区的时间序列数据映射为相空间矢量,并依据三个非线性独立度量指标(Sd,Hd和Nd)来评估两个脑区神经活动的相互依赖关系.GS算法可用于识别效应连接的方向,但有时三个度量指标所估计的方向存在不一致性.
2018年,Dsouza 等提出一种利用互连接分析(Mutual connectivity analysis,MCA)识别效应连接的算法[102].首先将每对脑区x和y的fMRI时间序列重构为相空间中d维的局部邻域模型,然后使用脑区x的局部模型预测脑区y的当前时刻值,并通过预测值和真实值的相似性度量判断x对y的预测能力,重复此过程,获取y对x的预测值,最终通过对比每对脑区间交叉预测能力的强弱识别脑区间的因果效应.与GS算法相比,MCA不仅能更准确地识别连接方向,而且在数据样本量不足时仍具有良好的性能,是一种精度高、适应性强的识别方法.
基于相空间重构的方法是一种识别脑区间非线性因果关系的可靠手段,但此类方法往往计算复杂度较高,且对数据噪声敏感,因此使用此类方法需要注意以下2个方面的问题:1)由于高维动力系统的运动轨道往往最终收敛到相空间的低维吸引子上,并且时序数据中包含大量的冗余信息,因此合理地选择嵌入维数和延迟时间对于提高识别方法的精度和效率十分重要;2)此类方法存在非线性系统对噪声和初始条件敏感的固有缺陷,而构建鲁棒性更强的平滑相空间将有望缓解此问题.
2.2.2 基于概率分布的识别方法
基于概率分布的识别方法是一类依据统计学的理论框架分析脑区间因果效应的方法,此类方法识别效应连接通常包括两个步骤:1)针对每对脑区fMRI数据的相关信息进行概率统计以获取脑区间的统计相关性;2)从获得的概率分布规律中预测脑区间神经活动的因果效应.近年来,一些基于概率分布的方法已逐渐在识别脑效应连接网络上发挥重要作用.
2006年,Patel等提出一种依据条件依赖关系度量每对脑区间效应连接的方法[103].该方法首先通过定义一个二进制值的列向量来表示每个脑区在一段采样时间内的激活状态,然后利用狄利克雷先验分布的多项式似然函数为每对脑体素的联合激活构建二元变量的伯努利贝叶斯模型,并计算不同联合激活状态下每对脑体素的联合激活概率,最终通过由联合激活概率所构成的度量指标κ和τ来评估每对脑区间的连接强度和连接方向.该方法对脑区连接方向的识别具有很高的准确性.
2013年,Hyvärinen 等提出了一种基于LiNGAM算法的似然比来度量两个非高斯变量间因果关系的方法[104],该方法首先度量每对脑区在两个方向上的因果效应的累积量,然后通过累积量测度的一阶近似似然比来识别脑区间的效应连接,是一种计算效率高且鲁棒性强的识别方法.
2019年,Bielczy k等提出了一种通过脑区BOLD信号分布函数的分数矩识别效应连接的方法[105].该方法首先对每个脑区fMRI 时间序列进行标准化处理,并计算其m阶原点矩,用于表示BOLD信号的多种不同分布情况(期望、方差和偏态等);然后计算这些BOLD分布的分数矩累积量,并根据每对脑区间不对称的分数矩累积量识别脑区间的效应连接.该方法综合考虑了BOLD信号多方面的信息,具有良好的鲁棒性和准确性.
2019年,Lee 等提出一种基于Copula 模型来估计每对脑区间有向依赖关系的方法[106].该方法首先假设每对脑区变量服从[0,1]上的均匀分布,然后利用这两个脑区变量的边缘分布和联合分布构建Copula模型,再分别通过一个脑区变量的边缘分布获取另一个脑区变量的边缘分布的条件期望值,并计算边缘分布的期望值与实际值的方差比,由于该比值刻画了一个脑区对另一个脑区的预测能力,因此依据每对脑区相互预测能力的强弱可以估计脑区间的因果效应.因为Copula模型对边缘分布的选择不受限制,并且能够测量脑区间的单调非线性相依关系,所以该方法是一种灵活、稳健地度量脑区间非线性因果关系的可靠方法.
上述内容对效应连接网络的识别方法进行了较详细的分类和介绍,并对一些代表性方法做了具体说明.为了能够更好地帮助读者理解和使用这些方法,我们从多个方面对各类方法中的部分典型方法进行了对比和总结,如表1所示.
3 脑效应连接网络识别面临的挑战
在脑科学研究中准确地识别脑效应连接网络,不仅有利于研究者探究人脑的功能整合规律,而且能为脑疾病的诊断以及病理研究提供重要的依据.
例如,在脑功能预测方面,如果发现人体的某些行为模式、情感认知与人脑的某些特定脑区的功能活动密切相关,那么可以对这些脑区的功能进行合理地预测;在脑疾病的诊断和治疗方面,如果能够及时、准确地发现人脑效应连接模式的异常,就有可能为脑疾病的早期诊断提供一些新线索,并为脑疾病的临床治疗和药物研发提供有力的帮助.
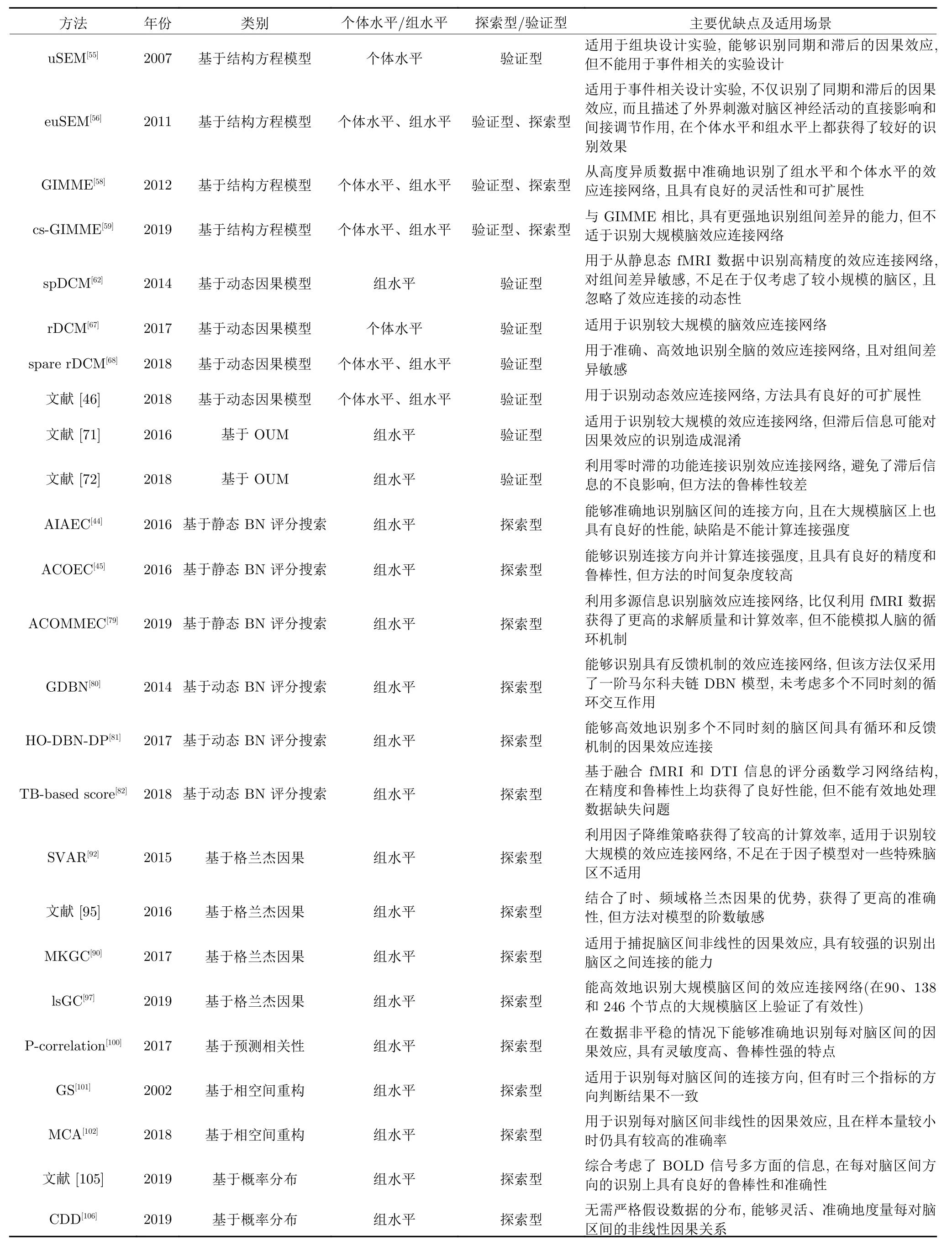
表1 人脑效应连接网络的部分典型识别方法对比Table 1 The comparisons of several typical identification methods on human brain effective connectivity networks
正是由于脑效应连接网络识别在脑科学研究中具有重要的应用价值,所以利用机器学习、数据挖掘中的一些理论和方法识别脑效应连接网络已成为脑信息学中一项重要的研究内容,并取得了许多丰硕的成果.但是随着研究的深入,一些极具挑战性的问题被凸显出来:
1)fMRI 数据的局限性问题:功能磁共振成像以其无创性、易重复、实验范式灵活、时空信息丰富等优势,已逐渐成为探究人脑功能复杂性和脑疾病诊断的一个重要手段.然而,目前基于fMRI数据识别脑效应连接网络仍存在一定的局限性:首先,由于fMRI 数据具有高维特性,因此直接从全脑体素的时间序列中识别效应连接网络十分困难;其次,在采集脑疾病患者的数据时,通常由于被试的配合度低难以保证所采集数据的完整性,导致实际可用于识别脑效应连接网络的fMRI数据样本量缺乏;再次,所采集的fMRI 数据通常包含较多的噪声,尽管预处理能在一定程度上消除噪声影响,但处理后的数据仍包含一些无法避免的干扰因素,造成fMRI数据的信噪比较低,影响了识别结果的准确性;最后,由于fMRI 数据的时间分辨率较低,导致基于时间滞后的方法容易识别到虚假连接,因此如何针对维度高、样本量少、噪声多、时间分辨率较低的fMRI数据,设计更有效的识别方法来克服fMRI数据的局限性是一个极具挑战性的研究问题.
2)脑效应连接网络的识别精度问题:在脑效应连接网络中,三个脑区x,y和z所构成的因果结构通常分为3种:a)链式结构(x→y →z):x与y之间存在效应连接,并且y与z之间也存在效应连接;b)v 型结构(x→z ←y):x和y均与z之间存在效应连接;c)同父结构(x←z →y):z分别与x和y之间存在效应连接.由于这三种因果结构具有马尔科夫等价性,因此已有的大多数识别方法难以对其进行准确的区分,导致所识别的脑效应连接网络极易包含错误的连接.此外,在链式结构和v 型结构中,两个不相邻脑区之间的因果效应通常因受第三个脑区的影响而包含间接的因果效应.因此,如何准确地识别因果结构并消除混淆变量的不良影响是识别高精度的脑效应连接网络的另一挑战性问题.
3)识别方法的实际应用问题:脑效应连接网络识别方法的发展不仅要有丰富而合理的理论作为支撑,还应该在脑认知和疾病诊断的实际应用中发挥作用.目前,大部分的研究思路是首先基于仿真数据集来验证方法的有效性,再利用真实数据集探究方法的实用性.虽然这在一定程度上能够挖掘方法的实用价值,但距离真正的实际应用仍存在较大差距.其面临的主要挑战在于真实的fMRI 数据集往往缺乏统一的衡量标准,难以对所识别的效应连接网络的可靠性进行评价.因此,如何为识别方法构建科学的评价标准或者从脑认知和工作机制的角度对识别结果进行更具解释性的分析是值得研究者们思考的问题.
4 未来的发展方向
面对上述挑战,结合脑科学的发展趋势和实际需求,以下几个方面的研究将是该领域今后重要的发展方向.
1)多模态脑影像信息融合的识别方法将是一个前沿热点:迄今为止,常见的脑影像数据除fMRI以外,还包括DTI、MRI、PET和EEG等.由于每一种脑影像数据仅从一个方面反映了大脑的某种特性,因此多模态脑影像信息融合的识别方法能充分利用大脑的时空信息,发挥各模态数据的优势,有效地缓解fMRI数据样本量不足、时间分辨率较低等问题,从而更加准确、高效地识别脑效应连接网络.目前已有一些研究利用多模态脑影像信息融合的方法来识别效应连接网络,取得了不错的效果.冀俊忠等设计了一种融合fMRI和DTI数据的脑效应连接网络蚁群学习算法[79],该算法利用结构信息来压缩蚁群搜索的空间,并从fMRI数据中提取的体素联合激活信息对蚁群寻优的启发函数进行修正,在识别效应连接的准确性和效率上均获得了良好的性能.Chiang 等提出了一种通过融合f MRI和MRI数据的贝叶斯矢量自回归模型识别效应连接网络的方法[107],该方法能准确地识别个体水平和组水平的脑效应连接网络.Riedl 等给出了一种基于代谢连接映射(Metabolic connectivity mapping,MCM)识别效应连接的方法[20],该方法首先利用fMRI数据构建视觉活动相关脑区的功能连接网络,然后通过PET数据获取局部能量代谢信息来识别功能连接网络中的信息流向.Anwar 等分别利用fNIRS、fMRI 和EEG数据对手指运动过程中皮层感觉运动网络进行格兰杰因果分析[108],并通过分析基于三种模态脑影像数据的识别结果来估计该网络的效应连接.这些多模态脑影像信息融合的识别方法的研究成果已经显现出一定的优势,但尚处于初步探索阶段.随着脑科学研究的进一步深入,人们获取的人脑的数据来源将越来越丰富,利用先进的计算机理论和技术进行多源信息融合,设计新颖、高效且准确的识别方法将是该领域未来追求的研究目标.
2)新型的高精度脑效应连接网络识别方法将层出不穷:识别准确可靠的脑效应连接网络对于探究人脑的功能活动规律、揭示脑疾病的生理病理机制具十分重要意义.为此,基于机器学习的新探索将日益丰富:
a)基于群智能优化的脑效应连接网络识别方法:研究鲁棒性更强的识别方法将是克服不良数据影响,获取高质量脑效应连接网络的关键.群智能算法是一类基于简单个体相互作用时涌现的整体智能行为而提出的元启发搜索方法.其特点是在求解各种组合优化问题时,通过模拟社会型生物(如蜜蜂、蚂蚁、鸟、鱼等)群体间的协作行为来有效获取问题的近似最优解.此类算法具有天然的自组织、自适应的随机搜索机制,能够在候选空间中进行全局搜索,即使在噪声较多等不良环境下仍具有全局收敛的鲁棒性.同时,群智能算法固有的并行机制使其具有高效搜索的能力,能够较快地获得全局最优解.因此基于群智能优化算法识别脑效应连接网络,有可能在识别精度和效率上均获得良好的性能.2019年,纪子龙等提出一种带有繁殖机制的脑效应连接网络萤火虫学习方法(Firefly algorithm with reproductive mechanism for learning brain effective connectivity network,FAR-EC)[109],该方法使用K2评分作为目标函数来衡量萤火虫个体的绝对亮度,通过萤火虫种群的迭代寻优和繁殖机制搜索最佳的脑效应连接网络结构,为识别高质量的脑效应连接网络提供了一个很好的范例,但该方法存在参数多、时间复杂度高等缺陷.为了克服这些不足,他们随后又提出一种基于双萤火虫种群并行搜索的脑效应连接网络学习方法(Parallel searching of double firefly populations for learning brain effective connectivity network,DFA-EC)[110],该方法首先将萤火虫种群划分为精英种群和普通种群,然后通过精英种群的定向移动和普通种群的随机移动分别逐步识别脑效应连接网络,再利用迁移操作动态地调整精英种群和普通种群的规模,以实现两个种群之间的信息交流.在经过一定代数的寻优后,使用基于多样性度量的种群自适应更新机制对两个种群进行更新,最终学习到最佳的脑效应连接网络.相较于FAR-EC,DFA-EC在识别精度和效率上均得到了显著提高.这些基于群智能优化算法识别脑效应连接网络的成功范例为识别方法的发展提供了新思路.但此类方法的研究和应用尚未形成更大的规模,在今后的研究中,如何利用群智能优化增强大规模脑效应连接网络的求解精度和效率将是值得尝试的研究方向.
b)基于深度学习的脑效应连接网络识别方法:随着深度学习的蓬勃发展及其在自然语言处理、图像和语音识别等诸多领域获得的巨大成功,该项技术已成为近年来最受瞩目的技术热点之一.目前,深度学习的主要模型包括卷积神经网络、递归神经网络、长短时记忆网络和生成式对抗网络等,还有一些蓄势待发的基于深度学习与其他方法相结合的混合学习方法正在引领前沿,如深度强化学习、深度森林和深度贝叶斯网络学习等.最近一些新的研究尝试利用基于深度学习的方法识别数据间的因果关系,并获得了不错的效果.2018年,Goudet等提出一种通过因果生成式神经网络(Causal generative neural networks,CGNNs)从观测数据中学习功能因果模型的方法[111],该方法利用深度生成模型的表征学习能力,同时考虑了数据的条件独立性和分布的不对称性,能够准确地识别数据间的因果结构.他们随后又设计了一种基于对抗生成式网络识别数据间因果关系的SAM(Structural agnostic model)[112]算法,该算法不仅能有效地消除混淆变量的影响,而且在噪声较多等不良条件下仍具有很强的鲁棒性.2019年,Njah等利用深度贝叶斯网络学习算法来预测大规模数据集中的因果关系[113],在可解释性和鲁棒性上均获得了良好的效果.同年,Dasgupta 等通过元强化学习训练了一个递归网络模型,将端到端学习与归纳推理相结合来识别数据间的因果结构,是一种准确、高效的因果推理方法[114].这些深度学习技术在因果网络识别中的良好表现为脑效应连接网络识别方法的发展开辟了新思路,我们相信基于深度学习方法识别效应连接网络将成为未来发展的必然趋势.
c)针对fMRI数据时序特性的识别方法将引起人们的广泛关注:获悉脑效应连接网络的动态变化以及滞后信息对当前脑效应连接网络的影响,对于理解人脑的功能整合和脑疾病的发病机制具有十分重要的实际意义.然而,目前大部分的识别方法是将每个脑区在一段采样时间内的fMRI 数据作为向量来分析脑区间的因果关系,并未针对fMRI数据的时序特性进行深入的研究.尽管基于格兰杰因果和基于动态贝叶斯网络评分搜索的识别方法考虑了fMRI 数据的时序特性,但它们在脑效应连接网络的识别上均存在一定的不足.所以,针对fMRI 数据的时序特性,结合新型的机器学习技术,设计新颖、准确且高效的识别方法将是今后该领域的一个重要研究方向.
d)脑效应连接网络的可解释性研究将引起人们的足够重视:在目前识别脑效应连接网络的方法中,仅有基于动态因果模型的方法在识别机理中融合了生理学模型,它利用一个血液动力学模型,将脑区神经生理学响应信号映射为BOLD响应信号,再通过模型反演来估计脑区间的效应连接.然而此类方法受到先验知识的制约,故其灵活性较差;而其他的识别方法虽然在识别精度和效率等方面各具优势,但它们均未将生理学的相关信息融合到识别机理中,故导致所识别的脑效应连接网络缺乏生理学的可解释性.目前,尽管已有一些研究在识别方法中引入了血液动力学模型[56,115],但这些方法的研究和应用仍处于初步阶段.因此,为了增强识别结果的可解释性,使所识别的脑效应连接网络能更真实地反映人脑的神经活动规律,更方便地应用于临床的脑疾病诊断,融合生理学模型的识别方法将有望成为该领域中新的研究热点.
5 总结
脑效应连接网络是人脑连接组研究中的一项重要的研究课题,识别脑效应连接网络已成为评价脑功能及其与神经退化疾病相关损伤的一种有效手段.本文首先系统地阐述了脑效应连接网络的识别过程,然后全面地分析和总结了脑效应连接网络的识别方法,最后深入地剖析了脑效应连接网络识别的挑战性问题,并对该领域未来的研究方向做了展望.总之,脑效应连接网络识别方法的创新与进步具有十分重要的理论意义和应用前景,一方面将推动脑认知和临床脑疾病诊断的发展;另一方面将为脑科学与人工智能的协同发展带来光明前景.
