草地贪夜蛾在不同小麦品种上的取食选择性和适应性及其与叶片生化物质含量的关系
2021-04-13陈巨莲
刘 欢, 张 勇, 陈巨莲
(中国农业科学院植物保护研究所, 植物病虫害生物学国家重点实验室, 北京 100193)
草地贪夜蛾Spodopterafrugiperda,俗称秋粘虫(fall armyworm),属鳞翅目(Lepidoptera)夜蛾科(Noctuidae)灰翅夜蛾属Spodoptera,是一种原产于美洲热带和亚热带地区的害虫(Sparks, 1979),具有寄主范围广(Montezanoetal., 2018; 梁沛等, 2020)、繁殖能力强(Johnson, 1987; 梁沛等, 2020)、生态型多样(Acevedoetal., 2018)、适生范围广(谢殿杰等, 2021)、迁飞扩散快(Sunetal., 2019; 梁沛等, 2020)和为害严重(Dayetal., 2017; 梁沛等, 2020)等特点。自2019年入侵我国后除严重威胁最适寄主玉米外,对小麦Triticumaestivum生产也产生潜在影响(徐丽娜等, 2019; 李艳朋等, 2020; 任学祥等, 2020; 张智等, 2020)。小麦是我国第三大粮食作物,2019年种植面积占全国农作物总播种面积的14.3%(国家统计局, 2019)。在草地贪夜蛾周年繁殖区、迁飞过渡区和重点防范区,小麦均有较大播种面积(张智等, 2020)。据全国农业技术推广服务中心发布的病虫情报显示,自2019年9月以来,分别在云南、安徽、江苏、河南、山东、四川6省15个县的小麦田查见草地贪夜蛾幼虫为害,且部分地区为害较重,如云南寻甸县严重田块虫株率高达 95%以上。田间调查也发现,草地贪夜蛾主要在冬前为害秋苗期小麦,田间分布型显示低龄幼虫呈聚集分布,高龄幼虫呈均匀分布(杨现明等, 2020);室内研究表明,取食小麦的草地贪夜蛾也能正常完成生活史,且在小麦上也表现出较高的适合度(吕亮等, 2020; 孙悦等, 2020)。由此可见,草地贪夜蛾对我国的小麦生产存在潜在威胁。
冬小麦-夏玉米轮作和春玉米-春小麦间作是我国许多粮食产区的主要种植制度。以黄淮海麦区为例:2019年在冬小麦-夏玉米种植区,河南省夏玉米种植面积近333.33万 hm2,其中草地贪夜蛾发生面积达1 758 hm2(孙悦等, 2020)。由于在草地贪夜蛾南北迁飞的春秋季节,黄淮海麦区玉米种植面积较少,因此其是否会因食料资源缺乏而“被迫”取食该区冬小麦,从而对小麦生产构成威胁尚不明确。昆虫的生物学指标和行为指标在一定程度上能反映昆虫对寄主的适应程度,如幼虫发育历期、存活率和化蛹率以及成虫羽化率(郭婷婷等, 2018),寄主适合度指数(host suitability index, HSI)(Maw, 1976)等。寄主植物营养物质(Dhillon and Kumar, 2017)和次生物质(王琛柱等, 1993)含量影响植食性昆虫的取食偏好和行为,进而影响其发育进程。亚洲玉米螟Ostriniafurnacalis在取食含有较高可溶性糖、可溶性蛋白、多酚氧化酶和超氧化物歧化酶含量的京科 968玉米花丝后其发育历期和平均世代周期明显延长,说明该品种具有一定的抗螟性(刘燊等, 2019)。单宁和总酚是植物体内与抗虫性相关的重要次生代谢物质,抗虫能力越强的木麻黄品系,其可溶性糖含量越低,总酚、单宁、黄酮含量越高,且可溶性糖含量与抗虫性关联度最大,其次是总酚、单宁和黄酮含量(林秀琴等, 2014)。然而,目前鲜见关于草地贪夜蛾对不同小麦品种适应性的相关研究报道,同时针对小麦叶片中生化物质含量对草地贪夜蛾取食影响的研究也较少。
因此,本研究采用室内恒温饲养观察的方法比较了草地贪夜蛾幼虫对不同小麦品种的取食选择;采用初步筛选的4个具感草地贪夜蛾特性和2个具抗草地贪夜蛾特性的小麦品种,测定了叶片生化物质含量,并利用Pearson相关性分析阐明了幼虫取食选择和叶片生化成分含量的关系;最后采用离体叶片饲喂的方法观察了草地贪夜蛾在这6个品种小麦上的适应性,进一步验证了品种的抗虫特性。研究结果为小麦抗虫品种受草地贪夜蛾为害后生理生化物质的变化及抗虫机制的解析奠定了基础,同时为小麦抗虫品种的合理利用提供了理论依据。
1 材料与方法
1.1 供试小麦品种及栽培
供试小麦品种为黄淮海麦区主栽品种鲁原502(LY502)、烟农21(YN21)、百农207(BN207)、山农17(SN17)、淮麦33(HM33)、济麦22(JM22)、周麦27(ZM27)、存麦16(CM16)、铜麦6号(TM6)、烟农19(YN19)、西农805(XN805)、存麦11(CM11)、淮麦46(HM46)及室内鉴定的抗蚜品种“KOK1679”和感蚜品种“矮抗58(AK58)”,共计15个品种。小麦播种于塑料育秧盒(直径×高=14 cm×19 cm)中,按照花卉营养土(鸿华庆伍园艺花土场)∶壤土=3∶1(m/m)的比例配置播种土壤,后放入人工气候室中(温度26±2℃,相对湿度70%±5%,光周期16L∶8D)中,苗龄25-30 d时供试验用。期间不施用任何化肥和杀虫剂。
1.2 供试虫源
供试昆虫于2019年9月采自云南德宏玉米田,在室内温度26±2℃,相对湿度70%±5%,光周期16L∶8D条件下饲养。低龄幼虫(1-3龄)采用新鲜玉米叶片(苗龄15-20 d)群体饲养,高龄幼虫(4-6龄)采用人工饲料单头饲养。饲料配方参考王世英等(2019)。成虫羽化后为其提供10%的蜂蜜水,在金属网笼(直径×高=26 cm×30 cm)的顶部和周边覆盖湿纱布供其产卵。
1.3 1和3龄幼虫对不同小麦品种叶片的取食选择性测定
参考叶碟法(Hanetal., 2015)进行。每小麦品种随机选取8段剪成5 cm长的叶片,用脱脂棉在一端包扎成一束供试,每品种为1组,共计15组。将供试叶片等距随机摆放在垫有湿润滤纸的白色塑料圆盘(直径×高=30 cm×3.5 cm)边缘,圆心处接入60头1龄幼虫,用扎有一定数量小孔的黑色保鲜膜封口,让其在黑暗环境下(Luginbill, 1928; 赵胜园等, 2019)自由取食,5 h后统计各组中叶片上的幼虫数量,试验重复15次。3 龄幼虫取食选择测定方法与1龄幼虫类似,但为避免3龄幼虫自相残食(Chapmanetal., 1999),每次只接入20头饥饿2 h 的幼虫,5 h后统计各组中叶片上的幼虫数量,试验重复15次。
1.4 不同小麦品种叶片生化物质含量测定
选取苗期小麦叶片为测试样品。每品种随机剪取1~2株小麦的新鲜叶片称重,后迅速放于盛有液氮的研钵中研磨成粉,研磨过程中持续加入液氮,确保操作环境低温。将研磨好的叶片粉末迅速转入离心管保存。每处理3个重复。采用植物可溶性糖检测试剂盒、蛋白定量(TP)测定试剂盒、植物总酚检测试剂盒(南京建成海浩生物科技有限公司)分别测定小麦叶片可溶性糖、总蛋白和总酚含量;采用单宁含量试剂盒(苏州科铭生物技术有限公司)测定小麦叶片单宁含量。可溶性糖含量的测定采用蒽酮比色法,蛋白含量测定采用考马斯亮蓝法,植物总酚含量测定采用分光光度法,单宁含量的测定方法为微量法。各物质含量测定的具体操作按照说明书进行。叶片可溶性糖、总蛋白和总酚含量的测定在分光光度计(屹谱仪器 U-T6A紫外可见分光光度计)上进行;单宁含量的测定在酶标仪[FlexStation 3 Molecular Devices,美谷分子仪器(上海) 有限公司]上进行。
1.5 草地贪夜蛾在不同小麦品种上的适应性观察
采用离体叶片饲喂法比较草地贪夜蛾在不同小麦品种上的适应性。各龄幼虫、雌雄蛹及成虫的识别采用赵胜园等(2019)的方法。首先将初羽化雌、雄成虫放入塑料养虫盒(4 000 mL)中配对,并提供 10%蜂蜜水,盒顶部覆盖纱布供其产卵。产卵高峰收集同日产卵块,放至底部铺有湿润滤纸的塑料养虫盒(长×宽×高=30 cm×21 cm×4 cm),盒盖开细密小孔保证通气,置于人工气候箱中(温度26±2℃,相对湿度70%±5%,光周期16L∶8D)。选取同日初孵幼虫供试,每品种重复6次,每重复观察12头幼虫。初孵至2 龄幼虫群体饲养,3 龄后在塑料指形管中(45 mL)单头饲养。每24 h更换一次叶片,直至化蛹。每日记录幼虫存活情况、化蛹日期,分辨雌、雄蛹并称重。收集的蛹依据对应编号按雌∶雄=1∶1配对转移至装有浸湿蜂蜜水棉球的指形管内,每日观察。记录成虫羽化日期和存活状态,直至死亡。计算草地贪夜蛾在不同小麦品种上的寄主适合度指数(HSI)。HSI=[(化蛹率×雌蛹重)/幼虫发育历期](Maw, 1976; Uyietal., 2018)。
1.6 数据分析
数据初步整理使用 Excel 2016,采用SPSS 23.0进行统计分析。各处理间幼虫取食选择率、叶片生化物质含量、草地贪夜蛾生物学指标及寄主适合度指数间的差异显著性采用单因素方差分析(one-way ANOVA)和Duncan氏检验进行分析;不同小麦品种叶片生化物质含量与1和3龄幼虫取食选择率的关系采用Pearson相关性分析进行分析。百分率数据在方差分析前进行反正弦平方根转换,以满足方差分析的要求。
2 结果
2.1 1和3龄幼虫对不同小麦品种叶片的取食选择性
1龄幼虫(图1: A)和3龄幼虫(图1: B)对不同小麦品种叶片的取食选择率存在显著差异(1龄:F=8.052,df=14, 224,P<0.001; 3龄:F=8.905,df=14, 224,P<0.001)。1龄幼虫依次对YN19, HM33, TM6, KOK1679, CM16和HM46 6个品种的取食选择性较高,取食选择率在7.34%~9.32%之间,而对其他9个品种的取食选择率均低于7%,且由高到低依次为:BN207, XN805, AK58, SN17, JM22, CM11, LY502, YN21和ZM27; 3龄幼虫依次对KOK1679, HM46, TM6, HM33和CM11这5个品种的取食选择性较高,取食选择率在8.35%~11.35%之间,而对其他10个品种的取食选择率均低于8%,且由高到低依次为:SN17, AK58, BN207, XN805, JM22, YN19, CM16, YN21, LY502和ZM27。

图1 草地贪夜蛾对不同小麦品种的取食选择性
2.2 不同小麦品种叶片中生化物质含量
根据 2.1节中幼虫取食选择性的结果,初步筛选1和3龄幼虫取食选择率均较高的 4个品种(HM46, HM33, KOK1679和TM6)和较低的2个品种(LY502和YN21)进行叶片生化物质含量测定及草地贪夜蛾适应性观测。
不同小麦品种叶片间营养物质和次生物质含量均存在显著差异(总蛋白含量:F=3.156,df=5, 17,P=0.048; 可溶性糖含量:F=3.779,df=5, 17,P=0.027; 总酚含量:F=7.379,df=5, 17,P=0.002; 单宁含量:F=3.938,df=5, 17,P=0.024)(图2)。由图2(A和B)可知,YN21叶片的总蛋白含量和可溶性糖含量最低。KOK1679, HM33和TM6叶片的总蛋白含量均显著高于YN21(P<0.05),而这4个品种叶片的总蛋白含量与其他两个品种LY502和HM46间无显著差异(P>0.05) (图2: A)。叶片的可溶性糖含量以TM6最高,其次为HM46,而这两个品种的可溶性糖含量均显著高于YN21(P<0.05),但是与其他3个品种HM33, LY502和KOK1679间均无显著差异(P>0.05) (图2: B)。YN21叶片的总酚和单宁含量最高,与LY502总酚含量间无显著差异(P>0.05),但显著高于KOK1679(P<0.05)(图2: C)。YN21, LY502和KOK1679叶片间单宁含量无显著差异(P>0.05),但这3个品种均显著高于TM6(P<0.05)(图2: D)。
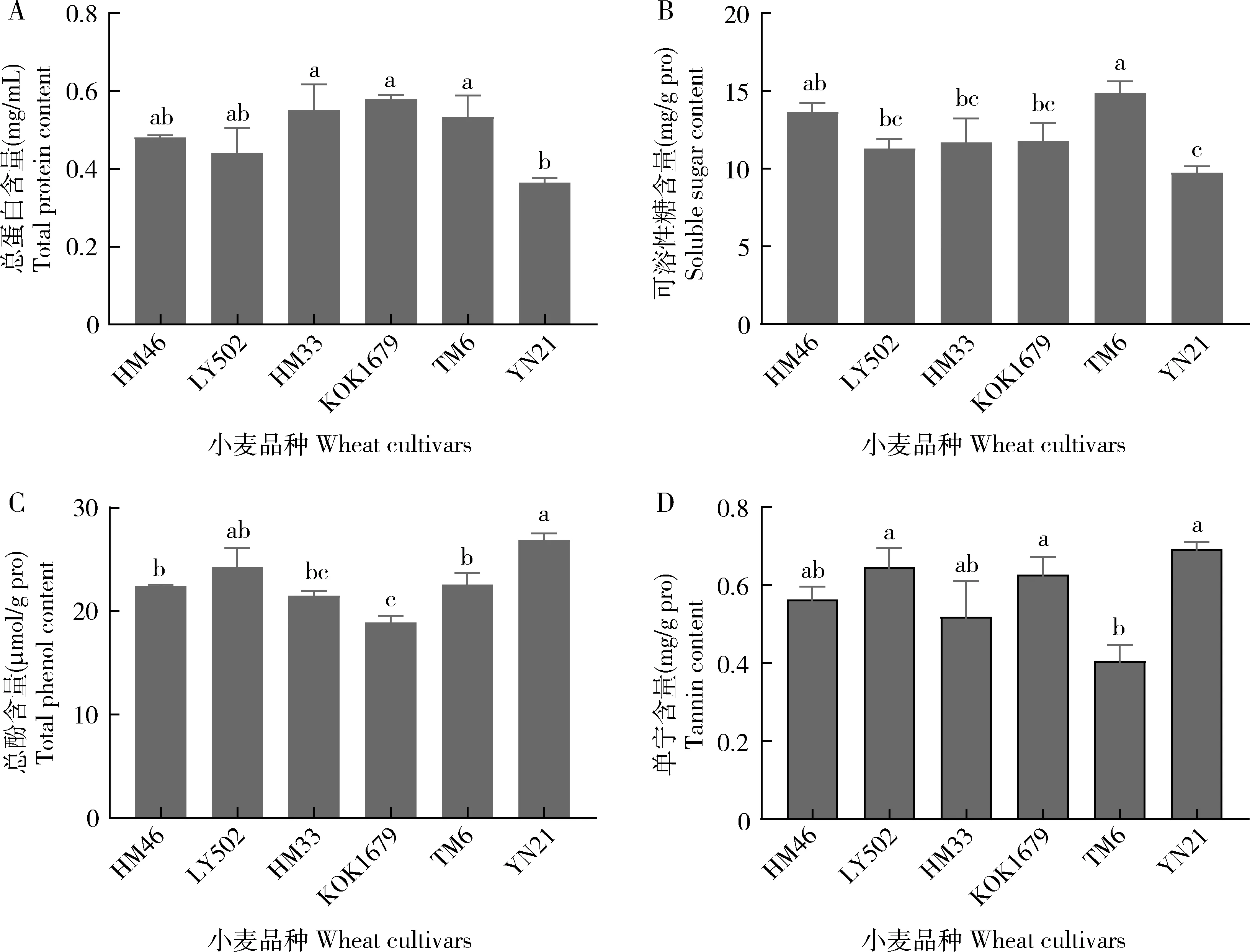
图2 不同小麦品种叶片中的生化成分含量
2.3 幼虫取食选择性与叶片生化成分含量间的关系
对草地贪夜蛾1和3龄幼虫对6个小麦品种叶片的取食选择率与其叶片生化成分含量进行Pearson相关性分析,结果如表1。 1龄幼虫的取食选择率与叶片总蛋白含量(r=0.674,P=0.002)、可溶性糖含量(r=0.472,P=0.048)呈显著正相关;与叶片总酚含量(r=-0.592,P=0.010)、单宁含量(r=-0.486,P=0.041)呈显著负相关;而3龄幼虫的取食选择率与叶片总蛋白含量(r=0.666,P=0.003)呈显著正相关,与可溶性糖含量(r=0.452,P=0.059)呈不显著正相关,与叶片总酚含量(r=-0.750,P<0.001)呈显著负相关,与单宁含量(r=-0.445,P=0.064)呈不显著负相关。

表1 草地贪夜蛾幼虫对不同小麦品种叶片的取食选择率与小麦叶片生化成分含量的Pearson相关性分析
2.4 草地贪夜蛾在不同小麦品种上的适应性
2.4.1幼期发育:草地贪夜蛾幼虫取食不同小麦品种叶片后,幼虫历期(F=140.901,df=5, 412,P<0.001)、蛹历期(F=5.088,df=5, 369,P<0.001)及蛹重(F=12.532,df=5, 407,P<0.001)均存在显著性差异(P<0.05)(表2)。取食LY502, HM33和HM46的幼虫,发育历期间无显著差异(P>0.05),但是与取食TM6和YN21的相比,幼虫历期显著较长,尤以取食LY502的最长,分别比取食YN21和TM6的长20.5%和26.2%。幼虫期取食TM6, YN21, KOK1679和HM46的蛹历期间无显著差异(P>0.05),但与取食LY502和HM33的相比,蛹历期显著较长;幼虫期取食HM33和HM46的幼虫,其蛹重间无显著差异(P>0.05),但与幼虫取食KOK1679, TM6, LY502和YN21的相比,其蛹重显著较重(P<0.05),取食HM46叶片的幼虫,其蛹重比取食YN21的高17.3%。
2.4.2幼虫存活率、化蛹率和寄主适合度指数(HSI):取食不同小麦品种的草地贪夜蛾,其幼虫存活率(F=2.182,df=5, 35,P=0.083)和化蛹率(F=0.958,df=5, 35,P=0.459)均无显著性差异(P>0.05),而寄主适合度指数(F=12.034,df=5, 35,P<0.001)存在显著差异(P<0.05) (表3)。对幼虫而言,TM6的寄主适合度指数最高,达15.6,显著高于其他5个品种(P<0.05);HM46和HM33的寄主适合度指数次之,二者间虽没有显著差异(P>0.05),但是均高于寄主适合度指数最低的YN21和LY502。
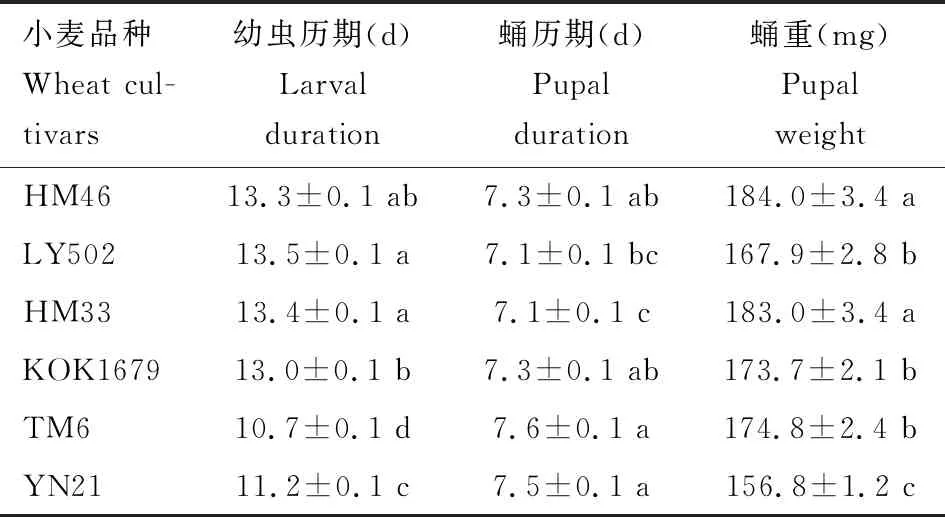
表2 取食不同小麦品种的草地贪夜蛾的幼虫和蛹历期及蛹重

表3 取食不同小麦品种的草地贪夜蛾的幼虫存活率、化蛹率和寄主适合度指数
2.4.3成虫羽化率、性比及寿命:取食不同小麦品种的草地贪夜蛾幼虫,其成虫羽化率(F=0.229,df=5, 35,P=0.947)和性比(F=0.987,df= 5, 35,P=0.442)均无显著差异(P>0.05) (表4),但成虫寿命间存在显著差异(F=15.324,df=5, 368,P<0.001),其中以幼虫取食TM6叶片后成虫寿命最长,达6.1 d,与幼虫取食LY502的无显著差异(P>0.05),但是均显著高于其他4个品种(P<0.05);以幼虫取食HM33的成虫寿命最短,仅为4.0 d。
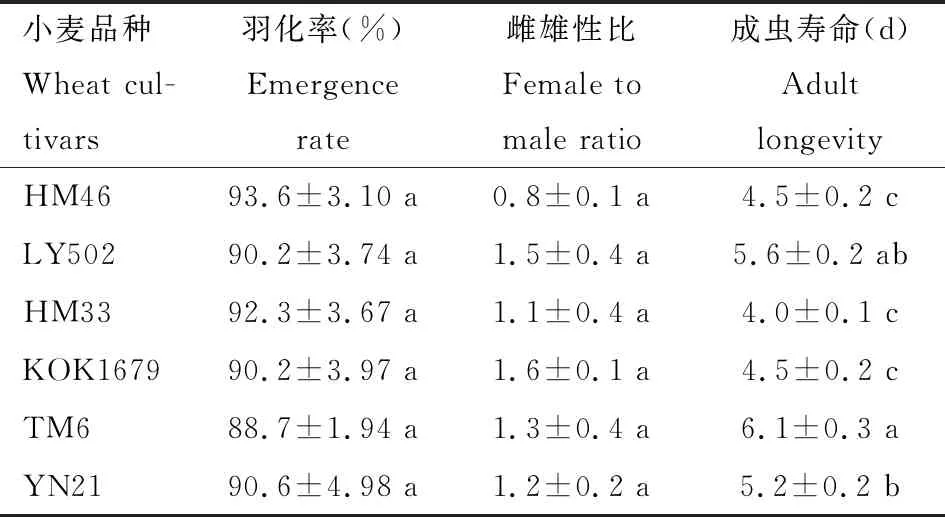
表4 取食不同小麦品种的草地贪夜蛾的羽化率、性比及成虫寿命
3 讨论
在植食性昆虫与寄主植物的互作中,昆虫进化出对复杂生态环境中寄主植物的取食选择及适应性。影响植食性昆虫对寄主选择性的因素很多,除了昆虫触觉、视觉、嗅觉、味觉等自身因素外(钦俊德, 2003; Britoetal., 2016),环境变化(陈瑜和马春森, 2009)、寄主形态(王美芳等, 2009)、寄主营养状态(葛超美等, 2018)、寄主挥发物(张娜等, 2009)、寄主发育阶段(Piyasaengthongetal., 2016; Feietal., 2017)以及自然天敌(郭林芳和李保平, 2008)等也影响昆虫对寄主植物的选择。从昆虫角度来讲,其嗅觉和视觉在寄主选择中发挥重要作用(钦俊德, 2003);从寄主植物方面来讲,植物挥发物是决定昆虫产卵的关键因素(Knolhoff and Heckel, 2014),而寄主发育阶段、营养状态及挥发物的综合作用则影响幼虫的取食选择(Piyasaengthongetal., 2016)。迫于植食性昆虫长期的选择压力,寄主植物也表现出相应的防御策略,即表现出一定的抗虫性。植物抗虫性主要表现为抗生性、不选择性和耐害性(Painter, 1951)。其中,不选择性是指植物不具备引诱产卵或刺激取食的特殊化学或物理性状,导致害虫不趋于产卵或取食。以嗜食寄主玉米为对照,仅以3种杂草为选择对象时,草地贪夜蛾成虫在婆婆纳Veronicadidyma上的产卵量显著高于其他两种杂草;而以玉米和婆婆纳为选择对象时,成虫在玉米上的产卵量显著高于婆婆纳;此外,1-4龄幼虫显著偏好取食玉米,而5和6龄幼虫对不同植物的取食选择性无显著差异(姚领等, 2020)。因此发现,当存在草地贪夜蛾的嗜食寄主玉米时,其低龄幼虫和成虫均优先选择在玉米上取食并产卵。本研究也发现,草地贪夜蛾幼虫对不同小麦品种的取食选择性存在显著差异,1和3龄幼虫均对HM33, TM6, KOK1679和HM46这4个品种的取食选择性较高,而对LY502和YN21的取食选择性较低(图1)。
植食性昆虫对寄主植物的选择和适应还与寄主体内营养物质和次生物质含量密切相关(秦秋菊和高希武, 2005)。植物营养物质含量的高低影响昆虫的生长发育,而次生物质含量的高低与其防御能力密切相关。Vanitha等(2015)测定了抗/感水稻叶片中部分化合物含量,发现抗虫水稻品种叶片内总酚含量高,而感虫品种叶片内糖含量、粗蛋白和总游离氨基酸的含量较抗性品种高。本研究也发现,小麦品种间在可溶性糖、总蛋白、总酚、单宁含量上存在显著差异。YN21叶片的总蛋白和可溶性糖含量最低,总酚和单宁的含量最高;TM6的叶片可溶性糖含量最高,而单宁含量最低;KOK1679叶片的总蛋白含量最高,总酚含量最低(图2)。这分别从营养物质和次生物质含量角度说明了YN21具有一定的抗虫性特征;而KOK1679和TM6具有一定的感虫特征。在本试验中,LY502叶片在总蛋白、可溶性糖、总酚和单宁含量上与YN21无显著差异(图2),因此与YN21表现出相似的抗虫特征,这也从生理生化角度映证了取食选择性试验中草地贪夜蛾对LY502和YN21取食选择性均较低的结果。
我们又对草地贪夜蛾1和3龄幼虫对6个小麦品种叶片的取食选择率与小麦叶片生化成分含量进行Pearson相关性分析,发现1和3龄幼虫的取食选择性与小麦叶片总蛋白含量和可溶性糖含量呈正相关;但与叶片总酚含量和单宁含量呈负相关(表1)。说明草地贪夜蛾幼虫偏好取食总蛋白和可溶性糖含量高的小麦叶片,不喜食总酚和单宁含量高的叶片,即总酚和单宁含量越高的小麦品种,其抗虫性越强,而总蛋白含量和可溶性糖含量越高,则小麦品种的抗虫性越低。这与林秀琴等(2014)对星天牛Anoplophorachinensis抗性与木麻黄Casuarinaspp.内含物含量关系的研究结果一致。表明小麦苗期对草地贪夜蛾的抗性主要是由较高水平的抗虫次生物质和较低水平的营养物质共同决定的,而不是单纯的通过降低小麦营养水平来抑制草地贪夜蛾的取食选择。而关于生化物质对草地贪夜蛾幼虫取食影响的机理有待于进一步研究。
抗生性作为植物抗虫性的重要表现特征,也已成为评价植食性害虫对寄主植物适应性的关键指标。抗生性是指因植物含有毒或不利于植食性昆虫生长发育的物质或缺乏足够营养物质而引起的植食性昆虫生长发育异常,如死亡率增加,化蛹率、羽化率降低,成虫寿命缩短,繁殖力下降等(Painter, 1951)。因此,昆虫的生物学指标能够较好地反映昆虫和寄主植物互作关系中植物对昆虫适应性的影响程度(钦俊德和王琛柱, 2001)。对于不同的寄主植物,昆虫对其选择性和适应性不同。如取食玉米、烟草和马铃薯的草地贪夜蛾,其幼虫发育历期、幼虫存活率、成虫寿命和繁殖力均存在显著差异,以取食偏好寄主玉米时,其种群发展最快(Guoetal., 2020);即使是同一寄主植物的不同品种,昆虫也表现出不同的适应性。在4个水稻品种中早39、中浙优8号、秀水134和甬优12中,取食秀水134的稻纵卷叶螟Cnaphalocrocismedinalis幼虫,其蛹重最轻,羽化后雌成虫的产卵量最低(王亭等, 2018),而蛹重作为反映昆虫体内能源物质储备及耐寒力的重要指标,能够较好衡量昆虫对环境的适应性(程鲁雪等, 2014; 刘燊等, 2019),因此综合评价,秀水134最不利于稻纵卷叶螟种群发展。本研究中,取食LY502的幼虫历期最长、蛹历期最短,幼虫历期显著长于取食TM6的,而蛹历期显著短于取食TM6的;幼虫期取食HM46的蛹重最大,显著高于蛹重最轻的YN21(表2);对幼虫而言,TM6的寄主适合度指数最大,达15.6,显著高于寄主适合度指数最低的YN21和LY502;同时,取食TM6叶片的幼虫,其成虫寿命也最长(表3和4)。就草地贪夜蛾对不同小麦品种适应性的整体表现而言,取食偏好品种TM6后,其发育速率最快,幼虫化蛹率较高,蛹重较大,寄主适合度指数最高,成虫寿命最长;而取食具有抗性特征的品种YN21后,幼虫发育速率变缓,化蛹率较低,蛹重最轻,寄主适合度指数最低(表3和4)。
抗虫品种利用和生物防治是草地贪夜蛾长期防控策略中提倡的较有效、安全的方法(杨普云等, 2019)。本研究从植物抗虫的不选择性和抗生性两方面建立了室内小麦品种对草地贪夜蛾抗性评价的方法,明确了小麦品种 YN21为草地贪夜蛾抗虫品种。对室内评价的抗虫品种,需要在草地贪夜蛾发生高风险的麦田生态区域进行田间抗性筛选,才能为田间抗性品种布局提供参考。本研究为深入研究小麦品种对该虫的抗虫机制奠定了基础。因此,基于植物抗虫机制的复杂性和草地贪夜蛾较强的适应性,小麦品种对草地贪夜蛾抗虫机制的研究是一项亟待开展的工作。
