网粒体脱落有助于茶小绿叶蝉成虫逃离蜘蛛网
2021-04-13林美珍王珍燕尤民生
林美珍, 杨 广, 王珍燕, 尤民生,*
(1. 福建农林大学应用生态研究所, 福州 350002;2. 福建农林大学闽台作物有害生物生态防控国家重点实验室, 福州 350002)
几十年来我国茶园防治茶小绿叶蝉Empoascaonukii主要依赖化学防治,导致该叶蝉已对多种杀虫剂产生抗药性(陈宗懋, 2009)。在有机生态茶园,我们经常在茶树冠层发现许多微型蜘蛛网,这种蜘蛛网被称为不规则网,其优势种群主要是草间小黑蛛(曾明森等, 2008; 毕守东等, 2019)。草间小黑蛛Hylyphantesgraminicola,又称为草间钻头蛛,属皿蛛科(Linyphiidae)钻头蛛属Hylyphantes,广泛分布于我国南北各省,食性广,是我国茶园、菜地、水稻田等农田作物的重要捕食性天敌(赵敬钊和刘凤想, 1982)。草间小黑蛛体小,所结不规则网既可以结网捕食,也可以游猎性捕食。王沅江等(2008)通过时空二维生态位的调查,认为茶小绿叶蝉若虫与草间小黑蛛的生态位重叠值较大,而成虫的天敌则主要以游猎型蜘蛛为主。谢振伦(1996)报道草间小黑蛛对茶小绿叶蝉成虫和若虫都有很高的日捕食量。目前多数学者都是通过室内试验测定草间小黑蛛在封闭容器内对多种害虫的捕食能力,未见到其结网捕食行为的相关报道(谢振伦, 1995; 刘凤想等, 2007),在野外草间小黑蛛能否通过不规则网对茶小绿叶蝉成虫具有较强的捕食功能尚未明确。另一方面,在前期研究中,我们观察到茶小绿叶蝉与叶蝉科其他昆虫一样,体表覆盖着一层被称为网粒体的特殊分泌物(Rakitov, 2000, 2009)。网粒体为微纳米级蜂窝状中空球体颗粒,依靠肉眼无法观察。Rakitov和Gorb(2013a, 2013b)报道网粒体具有超强疏水性,可以保护叶蝉排放的蜜露不会粘黏到翅面上。
Wittmaack (2005)报道叶蝉网粒体可被风吹散到空气中,成为气溶胶的一种微粒,对大气中的气溶胶微粒进行分析,发现网粒体占有很大比例,且一般成簇存在。但目前关于网粒体从叶蝉体表的脱落尚未见有直接证据。因此,本试验提出假设,茶小绿叶蝉是否可以通过体表网粒体脱落而快速逃离蜘蛛网。在前期研究中我们观察到茶小绿叶蝉成虫腹部腹面分布大量的网粒体,网粒体的超疏水特性有助于成虫逃离叶片表面残留的水滴(Linetal., 2016)。本试验分析茶小绿叶蝉成虫陷入蛛网后,在蜘蛛胁迫条件下以及无蜘蛛胁迫时的脱网策略,通过视频逐帧分析其翻身行为、挣扎行为和脱网行为,在这几个关键行为动作中判定叶蝉虫体各部位与蛛丝的粘黏情况,进一步明确茶小绿叶蝉体表网粒体是否具有脱落避险的生物学功能,同时也为保护和利用有机生态茶园草间小黑蛛防治茶小绿叶蝉的意义增加更多了解。
1 材料与方法
1.1 供试虫源
茶小绿叶蝉成虫采自福建农林大学南山茶园,试验所用叶蝉均为当天采集,保证试虫具有较强活力。试虫采用拍网法采集,带着手套拍打茶蓬表面,将试虫赶入捕虫网(100目纱网)内,用自制人工吸虫器将试虫收集到透明试管内。选择雄健、体色较深的试虫开展本试验。为明确试虫翅面网粒体覆盖的完好情况,利用相同采虫方式从相同采集地点采集100头茶小绿叶蝉成虫,用CO2麻醉后再随机抽选10头,在体式显微镜下使用显微剪将其前翅和后翅剪下,用显微镊夹住前翅或后翅的基部或端部,小心粘贴到铜台上,利用扫描电镜(SEM)观察试虫翅面网粒体的分布完整性。经验证,抽取的样本中叶蝉翅面网粒体均匀分布,完好性佳,表明拍网采集法不会导致试虫翅面网粒体脱落,样本中也未发现触碰过蛛网导致网粒体脱落的痕迹。
1.2 蛛网的收集
本试验所用蛛网采集自福建农林大学南区茶园。该茶园为有机生态茶园,蜘蛛种类较多。本试验选择草间小黑蛛的不规则网作为研究对象。每天上午9∶00时之前到茶园采集,采集时选择单枝茶梢上有完整的蛛网,如上下叶片间连接的不规则网或茶梢茎杆与叶片间连接的不规则网,将剪下的带蛛网茶梢插到泡沫板上,放入加有少许清水的亚克力箱内带回实验室。试验所用蛛网均为当天采集,选择干净、完整的新鲜蛛网,避免选择有较多尘土、虫体残留物或残缺破旧的老旧蛛网。
1.3 不同处理条件下茶小绿叶蝉逃离蛛网的主要行为参数统计及行为解析
草间小黑蛛一般躲在茶梢蛛网旁边的叶背等隐蔽处,将采集回来带有蛛网的茶梢先进行认真检查,确认茶梢上是否带有草间小黑蛛。试验将蛛网分为带蜘蛛处理组(S组)和不带蜘蛛处理组(NS组)。将带蛛网的茶梢通过泡沫板固定在加清水的烧杯上,试验利用4K高清摄像机(索尼FDR-AX100E)对茶小绿叶蝉成虫逃离蜘蛛网的行为进行拍摄记录,运用Vegas Pro 15.0软件对记录的行为片段进行逐帧分析,计算各种行为的时长。带蜘蛛处理组(S组),先将蜘蛛转移到指形管内进行饥饿处理2 h,将投网试虫经CO2麻醉后用毛笔轻轻转移到蜘蛛网上,约3 min茶小绿叶蝉苏醒后,将蜘蛛用毛笔转移到蛛网旁5 cm处的茶梢上,并开始拍摄记录捕食及逃逸行为,观察60 min内试虫的逃脱情况,统计脱网率(r1)和逃离时长(Te),逃离时长为从在蛛网挣扎开始计时至离开蛛网的时间;不带蜘蛛处理组(NS组),考虑田间茶小绿叶蝉成虫基本飞行状态下触碰草间小黑蛛的不规则蛛网,其背面碰网概率比正面高,故试虫麻醉后转移到蜘蛛网时统一背面贴网,待试虫3 min苏醒后,开始统计60 min内试虫的脱网率(r1)和挣扎逃离蛛网时长(Te)。此外,为明确茶小绿叶蝉成虫翅面覆盖的网粒体层是否对其逃离蛛网起到关键作用,通过Vegas Pro 15.0软件逐帧分析试虫的翻身率(r2)和翻身时长(Tt)。每个蛛网仅用于单次试验,S组的试验虫数为20头,NS组试验虫数为30头。
除了统计行为参数外,试验利用Vegas Pro 15.0软件分别对茶小绿叶蝉成虫触碰蜘蛛网后的挣扎及逃离的行为片段进行放大并逐帧观察分析,判定茶小绿叶蝉逃离蛛网的行为策略;重点分析成虫脱离蛛网的主要策略与哪些虫体部位有关,以便对这些部位进行显微观察,分析是否与其表面网粒体的分布有关。
1.4 茶小绿叶蝉体表网粒体分布特征的SEM观察
从野外采集的试虫中选择100头体色较深、虫体雄健茶小绿叶蝉成虫,用CO2麻醉后再随机抽选10头,剪下双翅进行步骤1.1节中翅面网粒体分布完整性的检验。将剪除双翅后的成虫用CO2再次麻醉,然后放入自制纱布小滤袋中,用针线绑紧滤袋口,放入2.5%戊二醛溶液中,4℃下固定12 h,取出后用PBS缓冲液反复漂洗3次,每次10 min,接着依次用30%, 50%, 70%, 80%, 90%和100%叔丁醇进行逐级脱水(郭素枝和季清娥, 2001),每次5 min,结束后用100%叔丁醇再次浸没5 min后取出,放入玻璃干燥器内自然干燥。将获得的样品在显微镜下用导电胶固定于样品台上,镀金(KEOLLV 1680,EIKO,离子溅射仪),扫描电镜(JSM-6380LV,日本电子株式会社JEOL)观察,分析试虫头部、胸部、腹部、翅面和3对足的网粒体分布密度。采用虫体每个部位扫描电镜图放大5 000倍进行分析,固定刻度尺为5 m,随机统计5个正方形方框(5 μm×5 μm)内的网粒体分布数量后取平均值。比较分析试虫与蛛网粘黏时容易脱离和难以脱离蛛网的虫体部位覆盖的网粒体分布情况。
1.5 茶小绿叶蝉网粒体触网后的脱落性显微观察
茶小绿叶蝉成虫双翅的面积较大且平整,能均匀覆盖网粒体,因此选择翅面对网粒体脱落性进行分析。使用0#毛笔将叶蝉成虫翅面触碰草间小黑蛛的不规则蛛网,随后将试虫与所粘连的蛛丝一起移到体式显微镜下,用显微剪将叶蝉翅面剪下,粘到贴有导电胶的铜台上,直接进行喷金及扫描电镜观察,查看翅面网粒体覆盖层的脱落情况以及蛛丝上是否有残留的网粒体颗粒。
1.6 数据分析
观测数据用SPSS21.0软件进行分析,不同处理组行为参数差异显著性分析采用独立样本t检验, 脱网率和翻身率的组间差异分析采用卡方检验。
2 结果
2.1 茶小绿叶蝉逃离蛛网的主要行为参数
当利用草间小黑蛛进行胁迫时,茶小绿叶蝉成虫具有45.0%的脱网率(表1),被捕率为55.0%,说明茶小绿叶蝉成虫具有较强的脱网能力避开结网性蜘蛛的捕食;当无蜘蛛胁迫时,茶小绿叶蝉成虫的脱网率达到76.7%,表明大部分的茶小绿叶蝉成虫都可以脱离蛛网的困扰,两个不同处理组间脱网率存在显著差异(df=1,P=0.047)。
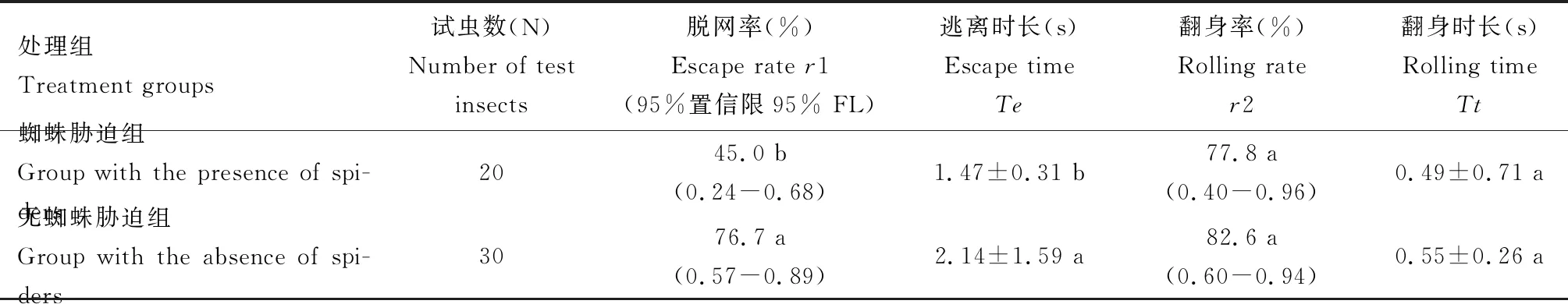
表1 茶小绿叶蝉成虫不同处理下逃脱蛛网的主要行为参数
从蜘蛛捕食视频可以看出,茶小绿叶蝉成虫挣扎后立即引起蜘蛛的猛扑行为。两个不同处理组中,存在差异显著的是逃离时长,蜘蛛胁迫组的逃离时长仅1.47±0.31 s,显著短于无蜘蛛胁迫组(t=-1.249,df=28,P=0.003),说明蜘蛛胁迫有助于叶蝉试虫快速脱离蛛网。对成功逃离蛛网的试虫脱网行为进行分析,我们发现茶小绿叶蝉成虫有一定比例可以不翻身情况下直接逃离蛛网,约80%左右的试虫都偏向于翻身后逃离,翻身率在两个不同处理组中不存在显著差异(df=1,P=1.000),翻身时长在两个不同处理组都很短,仅仅约0.5 s,不同处理组的翻身时长差异不显著(t=-1.01,df=28,P=0.315)。从翻身时长很短可以看出,成虫背面对蛛网似乎不存在黏力现象。两个处理组的逃离时长接近翻身时长的3倍,表明叶蝉成虫翻身后虫体仍有某些部位与蛛网的黏力较强,需经过一定时间的挣扎才能摆脱蛛网。
2.2 茶小绿叶蝉成虫陷入蛛网的行为解析
观察蜘蛛胁迫组,茶小绿叶蝉成虫表现出2种脱网策略:一种是翻身时触碰蛛网,引发蜘蛛过来捕食,但在捕食行为中叶蝉可快速跳跃逃脱,与蛛网的最后脱离部位为足的前跗节(图1: A),该种情况一般出现在蜘蛛从蛛网下方过来捕食,这在草间小黑蛛的捕食行为中很常见;另一种情况是蜘蛛还在旁边守候时,茶小绿叶蝉无翻身直接飞行逃离(图1: B)。无蜘蛛胁迫组,茶小绿叶蝉逃离蛛网的行为策略可分为4种:最常见的是快速翻身后向下跳跃逃离(图1: C);其次是成虫不翻身情况下通过调节腿部姿势直接飞行逃离(图1: D);第3种是翻身后在蛛网中挣扎,调节3对足与蛛丝的结合后腾空跳跃逃离蛛网(图1: E);第4种是叶蝉成虫翻身后移动到蛛网边缘,采用悬挂法挣扎后利用重力跳跃逃离蛛网(图1: F)。从以上行为分析可以看出,叶蝉背面尤其是翅面与蛛网的黏力较小,翻身时长很短,翻身十分容易,而足与蛛丝的黏力在不同部位存在差异,如果仅足的前跗节触网,叶蝉成虫仍可在蛛网上进行移动(图1: C)。
对茶小绿叶蝉,最难挣脱的部位是前足和中足,如利用悬挂法脱网的茶小绿叶蝉个体挣扎脱离蛛网的部位是前足和中足(图1: F)。分析蛛网上逃脱失败的茶小绿叶蝉个体,发现茶小绿叶蝉常因前足和中足被蛛丝粘黏无法及时挣脱而被蜘蛛捕食(图2: A),无蜘蛛胁迫组有的叶蝉个体挣扎时前足及中足拉扯蛛丝仍难以挣脱蛛网,超过60 min保持不动,在本试验中被记为脱网失败(图2: B)。

图1 茶小绿叶蝉成虫陷入蛛网后的逃离行为

图2 茶小绿叶蝉成虫陷入蛛网后逃脱失败的行为动作
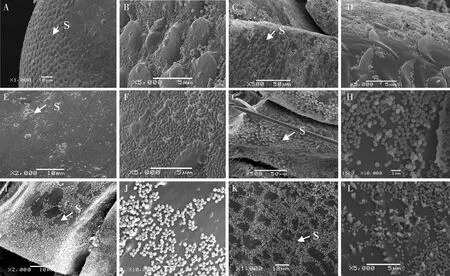
图3 扫描电镜观察的茶小绿叶蝉成虫头、胸和腹虫体部位的网粒体分布
2.3 茶小绿叶蝉成虫体表网粒体的分布特征
茶小绿叶蝉头、胸和腹3个虫体部位中,都出现虫体背面网粒体的分布数量明显少于腹面的情况。茶小绿叶蝉头部并非光滑,而是有微小凸起,但头部的网粒体分布数量较为稀疏(图3: A, B),分布密度仅为18±10粒/25 μm2,而头部腹面喙管和触角部位则堆积较厚的网粒体层,但分布不均匀(图3: C, D),网粒体堆积部位的分布密度>300粒/25 μm2;胸部背面的微观结构类似皮革纹,网粒体分布较为稀少(图3: E, F),表面覆盖的网粒体分布密度为22±12粒/25 μm2,胸部腹面有较多的微刺,堆积较多网粒体,分布也较不均匀(图3: G, H),分布密度>300粒/25 μm2;腹部背面微观结构较为光滑,网粒体呈现薄薄一层状,存在较多的无网粒体空隙(图3: I, J),表面覆盖的网粒体分布密度为90±42粒/25 μm2,腹部腹面微观结构为鱼鳞状排列的微刺,微刺周围上面覆盖较多的网粒体(图3: K, L),分布密度为>300粒/25 μm2。
从茶小绿叶蝉陷入蛛网的行为解析中,我们发现茶小绿叶蝉前足难以脱离蛛网,而后足则很少出现类似情况。通过扫描电镜比较观察茶小绿叶蝉前足和后足的显微结构(图4),前足腿节表面虽然有鳞片状结构的微小凸起,表面也有网粒体分布,但却覆盖不均匀(图4: B),网粒体稀少部位的分布密度为4±4粒/25 μm2,网粒体堆积部位的分布密度则>300粒/25 μm2。前跗节尤其是爪垫部位(Pu),相比后足更加肥厚,且光滑无网粒体(图4: C),后足的腿节则覆盖着厚厚一层网粒体(图4: E),后足前跗节爪垫较薄,且背面也是一层厚厚的网粒体(图4: F),网粒体堆积部位的分布密度>300粒/25 μm2,叶蝉后足的转动容易使表面的网粒体脱落包裹蛛丝。
从野外采集的茶小绿叶蝉,其前翅和后翅在扫描电镜下观察,观察到正面和背面都均匀覆盖一层白色的网粒体颗粒(图5: A, D)。前后翅虽然质地不同,前翅正面的显微结构为皮革纹路状(图5: C),后翅正面的显微结构为平滑膜质具微小乳突(图5: F),前后翅表面的微观结构虽然不同,但都均匀黏附微米级的网粒体颗粒(图5: B, E)。电镜扫描下放大10 000倍可清楚观察到网粒体的蜂窝状中空球体外观,直径约为0.30±0.10 μm,蜂窝孔小于100 nm(图5: C)。前后翅表面覆盖的网粒体分布密度为280±17粒/25 μm2。
2.4 茶小绿叶蝉翅面网粒体触碰蛛网后的脱落情况
茶小绿叶蝉前翅触碰草间小黑蛛的不规则蛛网后,可导致网粒体脱落留下一些空白痕迹,单独从这些空白痕迹无法判断网粒体是否因蛛丝粘黏而脱落,因此在本试验中将前翅触碰后的蛛丝保留在翅面一同进行扫描电镜观察,能在叶蝉翅面上看到蛛丝下方对应与蛛丝形状相似的空白区域(图6, LB所指的深色部位),同时便于分析网粒体与蛛丝接触的微观形态。草间小黑蛛的不规则蛛网中含有两种蛛丝,一种是含黏状梭型液滴的黏丝(图7: B),一种是不带黏性的拖丝(图7: F),利于蜘蛛行走。将图6中的S点放大获得的微观形态如图7(A)所示,交叉的两根黏丝碰巧都是黏珠部位,两滴黏珠重叠融合在一起,形成大滴黏珠触碰到翅面,底部空白,显示网粒体脱落并包裹到黏珠上。单根黏丝形态如图7(B)所示,可以看到大小相间黏珠,最后一滴黏珠触碰了叶蝉翅面;图7(C)是图7(B)最后一滴黏珠的放大图,展示单颗黏珠触碰翅面后被网粒体包裹的形态,图7(D)显示整根黏丝都被网粒体包裹,图7(E)将黏丝放大,可以观察到黏丝表面具有凹槽,网粒体可以附着在黏丝上方,草间小黑蛛的的拖丝表面光滑无黏性, 如图7(F)所示, 其表面则没有任何的网粒体。从图6和7可以看出,茶小绿叶蝉翅面与蛛丝触碰后,翅面的网粒体可脱落并有效包裹蛛网中的整根黏丝,包括液滴状黏珠和丝状部位。

图6 扫描电镜观察的茶小绿叶蝉成虫前翅网粒体触碰蛛网脱落留下的空白痕迹
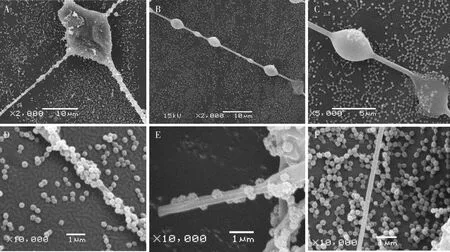
图7 扫描电镜观察的草间小黑蛛不规则网黏丝和茶小绿叶蝉成虫网粒体包裹黏丝的微观形态
3 讨论
昆虫利用体表覆盖物保护自己逃离天敌的捕食在自然界中并不少见,在蜘蛛网上已见报道的如鳞翅目昆虫的鳞片脱落(Diazetal., 2020)以及粉虱体表蜡粉颗粒的脱落对其逃离蛛网的帮助(Eisner, 2005)。叶蝉科昆虫的网粒体属于微纳米级颗粒,肉眼不可见,与鳞片或蜡粉颗粒相比,直径仅约为0.3 μm的网粒体是否能有效保护叶蝉逃离蛛网并不十分肯定。通过本研究,我们观察到体表覆盖的网粒体通过脱落并包裹蛛网中的黏丝,可以有效帮助叶蝉成虫脱离蛛网。
在本试验中,利用草间小黑蛛进行胁迫时,茶小绿叶蝉成虫成功躲避蜘蛛捕食的比例可达45.0%,无蜘蛛胁迫时,76.7%成虫可顺利脱离蛛网(表1),表明茶小绿叶蝉成虫具有较强的脱网能力,该结果与王沅江等(2008)的生态位调查结果相符,草间小黑蛛在茶树冠层所结的蛛网更适合捕食茶小绿叶蝉若虫。我们还观察到茶小绿叶蝉成虫苏醒后触动蛛网容易引起守候蛛网旁边的蜘蛛注意,蜘蛛胁迫组叶蝉逃离时长显著短于无蜘蛛胁迫组(表1),表明在危急状态下茶小绿叶蝉具有更快的脱网速度。从陷网行为分析中茶小绿叶蝉成虫至少有4种不同的脱身方法,其中悬挂法脱离蛛网的昆虫不止茶小绿叶蝉一种,Eisner(2005)报道草蛉成虫落入蛛网也采用类似的悬挂法成功脱离蛛网。
茶小绿叶蝉成虫陷入蛛网后的翻身时长很短,仅需0.55±0.26 s,约20%的叶蝉成虫甚至不翻身情况下也可直接跳离蛛网(表1),表明翅面与蛛网之间的黏力很小。茶小绿叶蝉前翅覆盖整个虫体背面的大部分面积,在陷入蛛网时翅面经常是粘黏面积最大的部位。通过扫描电镜的观察,我们发现前翅触碰到蛛丝时网粒体可从翅面脱落,在翅面上形成与蛛丝形态相同的空白斑,同时在蛛丝上可观察到网粒体包裹整根蛛丝的形态(图6)。Dong和Huang(2013)研究发现叶蝉科昆虫前翅存在网粒体储存区,叶蝉通过后足腿节和胫节的微毛对网粒体进行修饰涂抹。叶蝉翅面是虫体表面最平整的部位,网粒体覆盖最为均匀(图5: B, E),本研究中显示翅面分布的网粒体数量足够保护虫体脱离所粘黏的蛛丝,帮助叶蝉顺利翻身(图1: C)。但在本试验中,叶蝉成虫是经麻醉后转移到蛛网,人为统一将其背面触碰蛛网,因麻醉后的试虫翅面并未展开,该姿势有利于叶蝉通过翅面网粒体脱落性保护而快速翻身或不翻身直接挣脱蛛网,因此叶蝉的翻身时长和逃离时长在本试验中都相对较短。而在田间,叶蝉虫体展翅飞行触碰蛛网时可能露出腹面或头部被蛛网困住,这将给叶蝉脱离蛛网增加难度。Eisner(2005)观察草蛉成虫落入蛛网后的逃离策略时,发现草蛉除了翅面覆盖的浓密短毛可避免翅面与蛛网黏丝的直接接触,当草蛉其他虫体部位被蛛丝缠绕时,通过抽动腿部,利用口器和腿将黏性的丝线推到身体末端,集中切断缠绕在头部、足和触角的丝线进行脱身。本试验仅观察了茶小绿叶蝉体背触网时的有利脱网姿势,当茶小绿叶蝉其他触网姿势导致虫体头部或腹部被蛛网困住时,是否对其脱网率造成影响,或者茶小绿叶蝉是否存在其他高明的脱身策略都值得我们后期再深入研究。
扫描电镜结果显示,茶小绿叶蝉虫体背面除双翅以外的部位,头部背面、胸部背板以及腹部背面的网粒体分布数量较为稀少,而虫体腹面的网粒体分布数量相对较多(图3)。茶小绿叶蝉陷入蛛网后,约80%的试虫选择翻身后逃离蛛网。叶蝉翻身后除了3对足容易与蛛网接触,由于蛛丝与蛛丝间的空隙较大,虫体的腹面也容易接触蛛网。我们观察到叶蝉虫体腹面的网粒体分布数量比虫体背面更加密集,网粒体在腹面的分布极有可能对叶蝉腹面脱离蛛网具有帮助作用,这也是茶小绿叶蝉更偏向于选择翻身后脱离蛛网的可能原因。本试验目前所研究的触网姿势较为单一,尚未验证成虫腹部触网这种模式下网粒体在脱离蛛网中所发挥的作用,需在今后的研究中进一步完善。在前期研究网粒体的疏水性功能时,我们发现叶蝉腹部腹面较为密集的网粒体有利于茶小绿叶蝉成虫脱离茶叶表面的水滴,而若虫则因腹部腹面网粒体分布较少而难以脱离水滴(Linetal., 2016)。在研究中我们观察到草间小黑蛛捕食时喜欢将猎物拖拽离开蛛网后进行取食,若虫腹部与蛛丝接触表现出很强的黏力,导致草间小黑蛛拖拽若虫离开蛛网十分费劲,这可能与若虫腹部网粒体的分布较少有关,也值得我们进一步验证。
草间小黑蛛的蛛网具有黏丝和拖牵丝,黏丝结构与金蛛科相似,具有大小相间的黏珠(蒋平等, 2014),从扫描电镜图片看到整根黏丝都可被网粒体包裹,大滴黏珠触碰叶蝉翅面也会被网粒体包裹而无法直接触碰翅面。茶小绿叶蝉陷入蛛网需要脱离黏丝的困扰,在本研究中茶小绿叶蝉成虫仍有一定比例被蛛网困住而被草间小黑蛛捕食。对该部分试虫与蛛网的接触部位进行分析,发现叶蝉成虫前足和中足是虫体各部位中最难以脱离蛛网的部位,Dong和Huang(2013)报道叶蝉通过后足腿节和胫节的微毛对网粒体进行修饰涂抹,本研究中我们也发现后足腿节及前跗节覆盖较厚的一层网粒体,而前足的网粒体则数量较少,覆盖不均匀(图4: B),由此导致成虫前足是最容易被蛛丝困住的部位。
通过本研究,我们初步明确了网粒体对茶小绿叶蝉具有摆脱蛛网困扰的防御功能。网粒体除了脱落性可以起到防护外,Yang等(2017)报道网粒体蜂窝状的纳米孔可以反射波长250~2 000 nm的光波,对天敌可能具有隐身防御功能,但目前尚未有直接验证的相关报道。探索网粒体在茶小绿叶蝉防御天敌方面的生物学功能,有助于我们进一步了解该害虫,从更多角度考虑对该害虫的生态防控策略。
