Xiangbinfang granules enhance gastric antrum motility via intramuscular interstitial cells of Cajal in mice
2021-04-13QiChengChenZhiJiangJunHongZhangLiXingCaoZhiQiangChen
Qi-Cheng Chen, Zhi Jiang, Jun-Hong Zhang, Li-Xing Cao, Zhi-Qiang Chen
Abstract
Key Words: Interstitial cells of Cajal; Migrating motor complex; W/Wv; Gastric antrum motility; Xiangbinfang granules; Chinese medicine
INTRODUCTION
Interdigestive migrating motor complexes (MMC) induce periodic contractions of the gastric, small intestine, or colon smooth muscle, propagating along the gastrointestinal(GI) tract. Although the exact mechanism of MMC action still remains unclear, MMC have been classified into four stages. The MMC phase III consists of a high-frequency,high-amplitude rhythmic contraction, which has important physiological significance in normal GI motility and digestion[1]. Interstitial cells of Cajal (ICC), which are electrically coupled to smooth muscle cells (SMCs), produce slow waves that function as pacemaker activity[2]. There are several ICC subtypes based on their anatomical locations, morphology, and function in the GI tract. ICC associated with the myenteric plexus (ICC-MP) are responsible for producing slow waves. Intramuscular ICC (ICCIM), which are located in muscle bundles between muscle cells, mediate information transmission from autonomic nerves to SMCs[3]. In the small intestine of W/Wvmice lacking ICC-MP, there are no rhythmic slow waves and there are irregular contractions of longitudinal and circular muscles[4]. However, the MMC cycle still exists in the isolated intestine of W/Wvmice, suggesting that MMC do not require ICC-MP[5]. Other studies have shown that with the depletion of gastric ICC, disorders of the MMC cycle are common in diabetic gastroparesis[6,7]. This seems to indicate that ICC, especially ICC-IM, may also be involved in regulating the movement of MMC.
The Chinese herbal medicine xiangbinfang granules (XBF) is an effective treatment for promoting the post-surgical recovery of GI function[8,9]. Previous studies have shown that XBF significantly enhanced MMC activity in the antrum, pylorus,duodenum, jejunum, and the colon in beagles[10]. XBF was also found to promote contraction of the stomach, duodenum, and jejunum in healthy volunteers[11].However, the mechanism of XBF in enhancing MMC activity in the GI tract is still unclear.
In this study, we observed the MMC activity of the gastric antrum in W/Wvmice that lacked ICC-IM and analyzed the effects of the traditional Chinese medicine XBF in promoting gastric antrum motility.
MATERIALS AND METHODS
Animals
This study included 36 W/Wvand 36 wild-type (WT) mice. W/Wvmice were obtained from Jackson Laboratory (Bar Harbor, ME, United States) and bred in Guangdong Provincial Experimental Animal Center. The W/Wvand WT mice (weighing 18-25 g)were allowed free access to a standard laboratory chow diet and water and housed at a temperature of 22-24°C under a 12:12 h light/dark cycle. The study protocol was approved by the Institutional Animal Care and Ethics Committee of Guangdong Provincial Hospital of Traditional Chinese Medicine (No. 2018003). All animal procedures were conducted according to the Regulations for the Care and Use of Laboratory Animals in Guangzhou University of Chinese Medicine.
Tissue preparation
The mice were initially anesthetized with isoflurane before being killed by cervical dislocation. The whole stomach was placed in Krebs-Ringer buffer that was constantly perfused with oxygen. A cut was made along the lesser curvature of the stomach, the stomach was washed with ice-cold Krebs with the mucosa facing upward, and then the mucosa and submucosa were carefully removed.
Immunofluorescence staining of the gastric antrum
Ten W/Wvand 10 WT mice were used for immunofluorescence staining of the gastric antrum. The immunolabeling of mouse tissues was carried out on whole tissue mounts devoid of mucosa and submucosa. The whole mounts were stained with the primary antibodies for 1:100 c-kit (SC-168; Santa Cruz, CA, United States) and incubated for 1 h with 1:400 donkey anti-Rb IgG and Alexa Fluor 594. All immunostaining was imaged with a confocal microscope (Zeiss LSM 710; Göttingen, Germany) at an excitation wavelength of 594 nm. The c-kit-positive cells were observed under a 20 ×objective lens. The confocal micrographs shown are digital composites of the Z-series with a depth of 6-10 µm for ICC-MP and 15-30 µm for ICC-IM. The density of ICC-MP and ICC-IM was estimated by scanning through a 6-µm thickness of tissue area,counting the number of positive cells. The area of each micrograph was 424.3 µm ×424.3 µm = 0.18 mm2. The unit volume was 424.3 µm × 424.3 µm × 6 µm = 0.0011 mm3.The immunofluorescence staining of the gastric antrum was performed as previously described[12].
Intracellular microelectrode recording
Eighteen W/Wvand 18 WT mice were used for intracellular microelectrode recording.Full-thickness muscle strips (8 mm × 4 mm) were cut from the gastric antrum and pinned onto the base of a silica layer and continuously perfused with oxygenated Krebs-Ringer buffer at 37°C. Before recording, the strips were incubated for 2 h. The glass microelectrodes with resistance ranging between 50 and 80 MΩ were filled with 1 mol/L KCl for penetrating cells. The electrical response was recorded and amplified using a high input impedance amplifier (AXON 210 B; Molecular Devices, San Jose,CA, United States). The data were recorded on a computer using Clampfit 10.4 software.
Recording of MMC in the gastric antrum
When the 8 W/Wvand 8 WT mice were anesthetized by isoflurane inhalation, the limbs were fixed on the Baoding frame, and the abdominal hair was shaved with a razor and disinfected with iodophor. The abdominal cavity was carefully opened and the gastric antrum fully exposed. Then the micro-strain-gauge force transducers (J2 A-06-S108 N-10 C; Micro-Measurements, Raleigh, NC, United States) were sutured to the gastric serosa with a needle and nylon thread. The electrode leads were drawn from the back of the neck through the subcutaneous layer before closing the abdomen. After monitoring the animal’s wakefulness for more than 15 min, the micro-strain-gauge force transducers were connected to the Porti physiological recorder (TMSI,Netherlands) to measure the gastric contractile motion. The first 30 min were recorded for baseline control and the next 30 min for treatment. The gastric contractile frequency and amplitude index (sum of amplitudes within 20 min) were analyzed.
Solutions and drugs
XBF was produced by Hunan Academy of Chinese Medicine and contains Areca catechu L. (Binlang), Ginseng (Renshen), Fructus amomi (Sharen), Radix linderae (Wuyao)and Prunus persica Batsch (Taoren). Atropine (Lot A0132), tetrodotoxin (TTX, Lot T8024)and other chemicals were all acquired from Sigma (Sigma-Aldrich, St. Louis, MO,United States) unless indicated otherwise. The XBF, atropine, and TTX were dissolved in dH2O and further diluted in Krebs-Ringer buffer to the final concentration. Krebs buffer consisted of 121.9 mmol/L NaCl, 15.5 mmol/L NaHCO3, 5.9 mmol/L KCl, 1.2 mmol/L MgSO4, 1.2 mmol/L KH2PO4, 11.5 mmol/L glucose, and 2.5 mmol/L CaCl2.The pH of the Krebs buffer was 7.3-7.4 when bubbled with 95% O2-5% CO2at 37 ± 0.5°C.
Statistical analysis
The data are shown as mean ± SD. Welch’s t-test was used to compare the difference between WT and W/Wvmice. The paired t-test was used to analyze the data before and after administration. The number of MMC was compared by Fisher’s exact test. A P value < 0.05 was considered significant. Data statistics were calculated with GraphPad Prism 5 (GraphPad Software, La Jolla, CA, United States).
RESULTS
Immunogenicity of c-kit-positive cells in the gastric antrum of WT and W/Wv mice
Immunofluorescence showed that there were many c-kit-positive ICC in the myenteric plexus of the gastric antrum in WT mice. The c-kit-positive cells were connected to form a rich and dense ICC-MP network (Figure 1 Aa). In W/Wvmice, the c-kit immunogenicity of ICC-MP decreased (Figure 1 Ab). Compared with WT mice, the density of ICC-MP in the gastric antrum of W/Wvmice was significantly decreased(Figure 1 B; P < 0.001). A large number of bipolar ICC were found in the longitudinal and circular muscles of the gastric antrum in WT mice (Figure 1 Ac). Although sparse c-kit immunoreactivity was visible occasionally, no ICC-IM network was found in the antrum of W/Wvmice (Figure 1 Ad). There was a significant difference in ICC-IM density between WT and W/Wvmice (Figure 1 C; P < 0.001).
Characteristics of the gastric antrum slow waves in WT and W/Wv mice
The spontaneous periodic depolarization slow wave of the gastric antrum was observed in WT mice (Figure 2 A). In WT mice, the resting membrane potential (RMP)was –(56.49 ± 3.58) mV, the amplitude was 22.62 ± 2.23 mV, and the frequency was 6.16 ± 1.12 cpm. Spontaneous periodic depolarization slow waves with low amplitude were also observed in the antrum of W/Wvmice (Figure 2 B). In W/Wvmice, the RMP was –(52.95 ± 2.34) mV, the amplitude was 2.92 ± 0.52 mV, and the frequency was 7.48± 0.66 cpm. The difference determined was statistically significant between WT and W/Wvmice (Figure 2 C-E;aP < 0.05,cP < 0.0001).
Effect of XBF on the gastric antrum slow wave in WT and W/Wv mice
With perfused XBF at the concentration of 10-2g/L, the slow waves of the gastric antrum began to depolarize in WT and W/Wvmice (Figure 3 A and B). After treatment with concentrations of 10-2g/L, 5 × 10-2g/L, and 10-1g/L XBF, RMP decreased from−(56.49 ± 0.53) mV to −(52.31 ± 0.82) mV, −(51.08 ± 0.61) mV, and −(49.87 ± 1.58) mV,respectively. The frequency changed from 5.99 ± 0.18 cpm to 5.95 ± 0.44 cpm, 6.47 ±0.37 cpm, and 6.79 ± 0.14 cpm, respectively, and the amplitude decreased from 21.78 ±0.65 mV to 20.85 ± 0.90 mV, 19.56 ± 1.60 mV, and 18.15 ± 0.65 mV, respectively in WT mice. The differences in RMP and amplitude were significantly different at all XBF concentrations between the control and treatment groups (P < 0.05 or 0.01), and the frequency increased significantly except at the concentration of 10-2g/L (Figure 3 C-E).In W/Wvmice, the RMP decreased from −(53.13 ± 0.53) mV to −(49.47 ± 0.79) mV,−(48.41 ± 1.24) mV, and −(46.98 ± 1.40) mV, respectively. The frequency changed from 7.55 ± 0.12 cpm to 8.02 ± 0.19 cpm, 8.25 ± 0.18 cpm, and 7.65 ± 0.15 cpm, respectively,and the amplitude decreased from 2.84 ± 0.08 mV to 2.71 ± 0.11 mV, 2.87 ± 0.10 mV,and 2.66 ± 0.21 mV, respectively, at concentrations of 10-2g/L, 5 × 10-2g/L, and 10-1g/L. Compared to the WT controls, the differences in RMP were statistically significant at all concentrations (P < 0.01). At concentrations of 10-2g/L and 5 × 10-2g/L, the slow-wave frequency increased significantly (P < 0.05). However, the amplitude was not significantly altered at any XBF concentration in W/Wvmice.
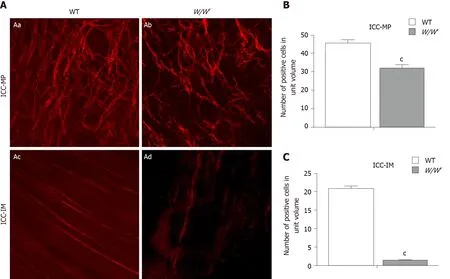
Figure 1 Immunogenicity of c-kit positive cells in the gastric antrum of wild-type and W/Wv mice. The c-kit positive cells were observed under a confocal microscope. The unit volume was 424.3 µm × 424.3 µm × 6 µm ≈ 0.0011 mm3. A: Immunostaining images showing the networks of interstitial cells of Cajal myenteric plexus (ICC-MP) and intramuscular interstitial cells of Cajal (ICC-IM), respectively: The abundant c-kit positive ICC-MP network of gastric antrum in wildtype (WT) mice (Aa); the sparse c-kit positive ICC-MP network of the gastric antrum in W/Wv mice (Ab); c-kit positive ICC-IM network of the gastric antrum in WT mice (Ac); no ICC-IM network distribution in the antrum of W/Wv mice (Ad); B and C: The number of c-kit positive cells in ICC-MP and ICC-IM was compared in the gastric antrum of WT and W/Wv mice under unit volume (424.3 µm × 424.3 µm) (n = 10, cP < 0.001). ICC-MP: Interstitial cells of Cajal myenteric plexus; ICC-IM:Intramuscular interstitial cells of Cajal; WT: Wild-type.
Effects of atropine and TTX on gastric antral slow waves induced by XBF
The results of intracellular recording showed that atropine could completely abolish the effect of XBF on gastric antrum slow waves in WT and W/Wvmice (Figure 4 A and B). After pretreatment with 0.5 mmol/L atropine, perfusion of XBF at a concentration of 10-1g/L did not alter the RMP, frequency, or amplitude of the gastric antrum slow waves in WT and W/Wvmice (Figure 4 C-E). In the presence of TTX 10-7M, perfusion of XBF at a concentration of 10-1g/L was able to decrease the RMP of gastric antrum slow waves in WT and W/Wvmice (Figure 4 F and G). TTX did not block the enhancement of gastric antrum slow waves in WT and W/Wvmice (Figure 4 H-J).
Characteristics of gastric antrum motility in WT and W/Wv mice

Figure 2 Characteristics of the gastric antrum slow waves in wild-type and W/Wv mice. A: Spontaneous rhythmic slow waves in the antrum of wildtype (WT) mice; B: Spontaneous rhythmic slow waves with the lower amplitude in the antrum of W/Wv mice; C-E: Comparison of resting membrane potential,frequency and amplitude of the gastric antrum slow waves between WT and W/Wv mice (n = 18, Welch’s t-test, aP < 0.05, cP < 0.0001). WT: Wild-type.
The MMCs in the gastric antrum were recorded by a micro-strain-gauge force transducer for 30 min. As shown in Figure 5, it was very difficult to distinguish between MMC phase I and phase II in WT. With the increase of irregular contraction frequency, regular contractions with the frequency 3.53 ± 0.18 cpm and amplitude index 23014.26 ± 1798.65 mV·20 min occurred in the antrum of WT mice. In the 30 min recording, there were only sporadic contractions in the gastric antrum of W/Wv.Occasionally, there was a strong paroxysmal contraction with a frequency of 1.28 ±0.12 cpm and an amplitude index of 3782.16 ± 407.13 mV·20 min. The differences in the frequency and amplitude index were all statistically significant (P < 0.0001). We observed that 7 out of 8 WT mice developed a total of 13 MMC III phase contractions,while in the 8 W/Wvmice, only 2 developed 3 MMC phase III-like contractions of shorter duration, lower amplitude, and lower frequency. There was a significant difference in the number of MMCs between both groups (Fisher’s exact test, P =0.0406). For the gastric antrum of WT mice, the duration of the MMC phase III was 151.08 ± 8.87 s, the amplitude was 315.45 ± 5.55 mV, and the interval between MMCs was 10.75 ± 0.61 min. In the gastric antrum of W/Wvmice, the duration of the MMC phase III-like contractions was 123.67 ± 2.96 s, the amplitude was 194.12 ± 4.76 mV,and during the 30 min observation, no phase III-like contractions were found twice in the same W/Wvmice. Compared to WT mice, the duration and amplitude of the MMC III phase in the gastric antrum was significantly reduced in W/Wvmice (P = 0.0117 and 0.0020, respectively), suggesting there was no typical MMC cycle in the gastric antrum of W/Wvmice.
Effect of XBF on gastric antrum motility in WT and W/Wv mice
After the administration of XBF (5 mg), strong contractions in the antrum of WT mice were observed instantaneously (Figure 6). The periodic MMC movement turned into a high frequency and high amplitude MMC III phase contraction. However, XBF did not significantly enhance the contraction of the gastric antrum in W/Wvmice (Figure 6).After treatment in turn with 0.6 mg, 1.25 mg, and 5 mg of XBF, the frequency index of the gastric antrum in WT mice was 3.16 ± 0.39 cpm, 3.30 ± 0.26 cpm, and 5.58 ± 0.62 cpm and the amplitude index was 13415.25 ± 1694.38 mV·20 min, 24537.89 ± 2406.33 mV·20 min, and 51807.48 ± 9255.04 mV·20 min, respectively, while the frequency index of the gastric antrum in W/Wvmice was 1.85 ± 0.48 cpm, 1.93 ± 0.37 cpm, and 1.99 ± 0.37 cpm and the amplitude index was 6488.43 ± 1490.04 mV·20 min, 7733.07 ±1469.27 mV·20 min, and 6901.26 ± 807.34 mV·20 min, respectively. These results demonstrated that with high-dose XBF treatment, there was a significant difference in the frequency and amplitude indices of the gastric antrum between WT and W/Wvmice (all P < 0.05 or 0.01).


Figure 3 Effect of Xiangbinfang granule on gastric antrum slow wave in wild-type and W/Wv mice. A: Representative trace of the effect of different concentrations of Champaign formula on gastric antrum slow waves in wild-type (WT) mice. After infusion of Xiangbinfang granule (XBF) at the concentration of 10-2 g/L (Aa), 5 × 10-2 g/L (Ab) and 10-1 g/L (Ac), the slow waves resting membrane potential (RMP) decreased, frequency increased and amplitude decreased; B:Representative trace of the effect of different concentrations of Champaign formula on gastric antrum slow waves in W/Wv mice. Infused XBF at the concentrations of 10-2 g/L (Ba), 5 × 10-2 g/L (Bb) and 10-1 g/L (Bc) decreased the RMP in W/Wv gastric antrum slow waves; C-E: Histogram of XBF for gastric slow wave RMP (C),frequency (D) and amplitude (E) in WT and W/Wv mice. (Control, n = 18; treatment, n = 6. Paired t-test was used to compare the difference before and after administration, aP < 0.05, bP < 0.001, cP < 0.0001). WT: Wild type; XBF: Xiangbinfang granule.
Effects of atropine and TTX on the gastric antrum motility induced by XBF
To clarify the mechanism of XBF in promoting antral motility, atropine and TTX were injected intraperitoneally, and then XBF was administered by gavage. The results indicated that contractions were inhibited in the antrum of WT and W/Wvmice after intraperitoneal injection of 0.1 mg/kg of atropine, and subsequent treatment with 5 mg of XBF did not increase the contractile activity of the gastric antrum (Figure 7 A and C). These results suggest that atropine could completely eliminate the motilitypromoting effect of XBF in both WT and W/Wvmice (Figure 7 E). The frequency and amplitude of gastric antral contractions were abolished by 0.05 mg/kg of TTX.Subsequent treatment with 5 mg of XBF induced a contraction enhancement in WT and W/Wvmice (Figure 7 B and D). In comparison with XBF without TTX, TTX pretreatment could partially reduce the gastric antrum motility-promoting effect mediated by XBF (Figure 7 F).
DISCUSSION
C-kit is a necessary condition for the development of a GI pacemaker system[13].Previous studies have shown that the W-mutation causes loss of the ICC network in the GI tract of W/Wvmice, such as the disappearance of ICC-MP and decreased ICCMP in the jejunum and ileum[4]. In the W/Wvcolon, the density of c-kit positive ICCMP was reduced by 50%-60% compared to that of WT mice[12]. While a normal ICC-MP network was observed, the ICC-IM disappeared completely in the gastric antrum of W/Wvmice[14]. We scanned whole muscle specimens to obtain a more accurate number of ICC and compared them to WT mice; about 60% of ICC-MP was retained in the antrum of W/Wvmice, and no ICC-IM network was observed. GI slow waves are mainly produced by ICC-MP[2], and ICC-IM is responsible for the regenerative component of the slow wave[15]. Our immunofluorescent staining showed that the ICCMP network density decreased but did not disappear, and the slow-wave activity in W/Wvmice was preserved. However, due to the lack of an ICC-IM network in the antrum of W/Wvmice, there is no regenerative component in the W/Wvantrum slow wave. Therefore, W/Wvmice showed a low-amplitude slow wave and disturbed movement in the antrum. ICC-IM were also innervated and provided mechanisms for neural signal transduction to the gastric musculature[16]. ICC-IM were densely innervated by excitatory and inhibitory enteric motor neurons and in close contact with nerve terminals. ICC-IM played a role in both nitrergic inhibitor and cholinergic excitatory motor neurotransmission in the gastric antrum[17,18]. Excitatory junctional potential and inhibitory junctional potential after intrinsic nerve stimulation were greatly attenuated in the antrum of W/Wvmice, and the reduced density of ICC-IM leads to reduced neural regulation in the W/Wvantrum[19]. Hirst et al[19]have shown that excitatory vagal stimulation response resembles the regenerative response which was initiated in this tissue by ICC-IM. Regenerative responses were the dominant responses produced by neural stimulation. This suggested that ICC-IM is regulated by vagus nerve.


Figure 4 Effects of atropine and tetrodotoxin on the enhancement of gastric antrum slow wave by Xiangbinfang granule. A and B:Representative trace of the effect of Xiangbinfang granule (XBF) 10-1 g/L on slow waves of the gastric antrum pretreated with atropine 5 mmol/L in wild-type (WT) (A)and W/Wv mice (B); C-G: Histogram of atropine blocking the effect of XBF on the enhancement of resting membrane potential (C), frequency (D) and amplitude (E) of antral slow waves. Representative trace of the effect of 10-1 g/L of XBF on slow waves of the gastric antrum pretreated with 10-7 M of tetrodotoxin (TTX) in WT (F) and W/Wv (G) mice; H-J: Histogram of TTX blocking the effect of XBF on the enhancement of the resting membrane potential (H), frequency (I) and amplitude (J) of antral slow waves. (Compared with pretreatment, n = 3, aP < 0.05). WT: Wild-type; XBF: Xiangbinfang granule; TTX: Tetrodotoxin; CON: Basic control; ATR: Atropine.
The MMC has been found to be a complex system that may be regulated by motilin[20], the enteric nervous system, and the vagus nerve. Mondal et al[21]found that treatment with motilin induced phase III contractions in vivo and in vitro, while motilin antagonists can abolish the occurrence of spontaneous gastric phase III contractions.Their other experiment showed that motilin-induced gastric contractions were mediated through the myenteric plexus in a vagus-independent manner[22]. ICC-IM may play an important role in this process.

Figure 5 Characteristics of gastric antrum motility in conscious wild-type and W/Wv mice. Micro strain-gauge force transducers were implanted into the gastric antrum to observe the migrating motor complex (MMC). A: Representative trace of the gastric antrum contractions in conscious wild-type (WT) and W/Wv mice. The black arrow shows the MMC III phase contraction; B and C: The histograms of the comparison of the contractile frequency (B) and amplitude index(C) of the gastric antrum in WT and W/Wv mice. (n = 8, Welch’s t-test, cP < 0.0001). WT: Wild-type.
The MMC have a temporally coordinated cyclic motor pattern during the interdigestive state of the stomach and small intestine in many animals. In the human and dog gastrointestinal tracts, the occurrence of MMC is regulated at 90-120-min intervals[23,24]. Because of motilin and motilin receptor pseudogenes[25], mice and rats have shown different MMC patterns. Strain-gauge force transducer implantation is a crucial technique for recording GI MMC movement in conscious animals[23,24,26,27].Takayama et al[26]examined the gastrointestinal motility of W-mutant rats by an extraluminal strain-gauge force transducer method. They found the duration of MMC was 2.5 ± 2.3 min and the interval of MMC was 5.4 ± 2.9 min. Similar results were observed by Taniguchi et al[27]in their study, spontaneous phase III contractions were observed every 13-16 min. It was difficult to record the gastrointestinal motor pattern of the mice in situ. Spencer et al[5]tried to record MMC in the isolated small intestine of mice. They found the interval between MMCs in the mouse small intestine was 5 ± 1 min, and the durations of MMC contractions were about 30 s. The MMC was regulated by motilin, the enteric nervous system, and the vagus nerve. Motilin-induced contractions are much less potent than those of MMC in vivo[28]. The complex regulatory system of MMC was not complete in vitro. Therefore, we miniaturized the strain-gauge force transducer to facilitate MMC recording in the gastric antrum of conscious mice. In this study, we found that the duration of the MMC phase III was 151.08 ± 8.87 s, the amplitude was 315.45 ± 5.55 mV, and the interval between MMCs was 10.75 ± 0.61 min in WT mice. There was no complete MMC cycle in the gastric antrum of W/Wvmice. The gastric antrum of W/Wvmice lacked ICC-IM and the corresponding motor nerve conduction was inhibited[29]. The typical MMC disappeared in W/Wvmice, suggesting that ICC-IM is an important factor in regulating the MMC activity.
In this study, we found that XBF enhanced the contractions of the gastric antrum in WT mice through slow-wave depolarization of SMCs. The effect of XBF on enhancing the antral motility was greatly reduced in W/Wvmice. ICC specifically express calcium-activated chloride channels (CACC), which are regulated by anoctamine 1(Ano1). Exogenous nerve stimulation or GI hormones could act on ICC to produce slow waves by CACC[30]. SMCs respond to slow waves to generate action potentials by activating voltage-dependent L-type calcium channels[31]. Under physiological conditions, cell-membrane potential depolarization increased the probability of the Ltype calcium channels opening, allowing calcium influx to induce SMCs to generate action potentials and contractions[32-34]. In Ano1-knockout mice, the excitability of smooth muscle contraction induced by carbachol was decreased. This is caused by cholinergic nerve stimulation that first activates muscarinic receptors on ICC-IM and then stimulates the CACC channel, thereby causing the SMCs to depolarize and generate action potentials through gap junctions[35]. Thus, the response of the distal stomach to cholinergic stimulation was weakened and blocked the neural activation of regenerative potentials in W/Wvmice deficient in ICC-IM, and the low level of potential depolarization during the slow-wave plateau period reduced the probability of L-type calcium channels opening. Therefore, XBF depolarized the SMCs slow waves(Figure 3) but did not significantly enhance the mechanical contraction (Figure 6) of the gastric antrum in W/Wvmice. Our study showed that atropine completely blocked the gastric antral motility induced by XBF, but TTX just partially reduced the effect of XBF in both WT and W/Wvmice (Figure 7). This suggested that XBF enhances the smooth muscle contraction of the gastric antrum through the cholinergic pathway of ICC-IM and the enteric nervous system.

Figure 6 Effect of Xiangbinfang granule on gastric antrum motility in wild-type and W/Wv mice. A: Representative trace of the enhancement of the gastric antrum motility in wild-type (WT) and W/Wv by Xiangbinfang granule (XBF); B: Comparison of the antral contractile frequency of WT and W/Wv mice after intragastric administration of 0.6 mg, 1.25 mg and 5 mg XBF; C: Comparison of antral contractile amplitude index of WT and W/Wv mice after intragastric administration of 0.6 mg, 1.25 mg and 5 mg XBF (n = 8, Welch’s t-test, aP < 0.05, bP < 0.01). WT: Wild-type; XBF: Xiangbinfang granule.
The Chinese medicine XBF, composed of Areca catechu L., Ginseng, Fructus amomi,Radix linderae, and Prunus persica Batsch, promotes entire GI motility in the treatment of postoperative ileus[8-11]. Previous studies have shown that Areca catechu L. enhances gastric motility in healthy people[36]. Arecoline, the main active component of Areca,with an XBF content of 0.112 mg/g[37], promoted GI motility through the M3 receptor[38,39]. Ginsenoside RF, an extract of Ginseng, induced membrane depolarization of ICC[40]and Fructus amomi promoted the contraction of the antrum and duodenum in beagle dogs[41]. The promotion of GI motility by XBF is the result of the interaction of these Chinese herbs, and the most important active components still need to be further characterized.
CONCLUSION
Our study shows that ICC-IM play a crucial role in regulating gastric antral MMC activity. They may be an important bridge between the vagus nerve, the enteric nervous system, and motilin in regulating smooth muscle contraction. Through the muscarinic receptor pathway on ICC-IM, XBF depolarizes SMCs and initiates an action potential, changing the periodic motion of MMC into a phase III-like contraction pattern in the gastric antrum of mice.

Figure 7 Atropine and tetrodotoxin block the enhancement of antrum motility induced by Xiangbinfang granule. A-D: Intraperitoneal injection of 0.1 mg/kg atropine (ATR) completely blocked the enhancement effect of Xiangbinfang granule (XBF, 5 mg) on gastric antrum in wild-type (WT) (A) and W/Wv (C)mice. Tetrodotoxin (TTX) partially reduced the gastric antrum motility enhancement mediated by XBF in WT (B) and W/Wv (D) mice; E-F: Comparison of the effect of XBF 5 mg of antral contractile amplitude index in WT and W/Wv mice after pretreatment with ATR 0.1 mg/kg (E) and TTX 0.05 mg/kg (F). (n = 4, Welch’s t-test, aP <0.05, bP < 0.01, cP < 0.001). WT: Wild-type; XBF: Xiangbinfang granule; TTX: Tetrodotoxin; CON: Basic control; ATR: Atropine.
ARTICLE HIGHLIGHTS
Research background
The Chinese medicine Xiangbinfang granule (XBF) is an effective prescribed treatment for promoting the recovery of gastrointestinal (GI) function post-surgery. In previous studies, we found that XBF mediated the phase III contraction of migrating motor complexes (MMC). However, the mechanism of XBF in enhancing MMC activity in the GI tract is still unclear.
Research motivation
In this study, we observed the MMC activity of gastric antrum in W/Wvmice that lack intramuscular interstitial cells of Cajal (ICC-IM) and analyzed the effect of the traditional Chinese medicine XBF in promoting gastric antrum movement. From this study, we will further understand the role of ICC-IM in MMC activities. Meanwhile,the mechanism of XBF promoting gastrointestinal motility through ICC-IM was discussed, so as to provide the basis for the development and application of XBF.
Research objectives
W/Wvmice were used to observe the effects of ICC-IM on gastric antrum motility and to establish the mechanism of XBF in promoting gastric antrum motility. We further investigated the correlation between ICC-IM and MMC in mouse gastric antrum.
Research methods
The density of c-kit positive ICC myenteric plexus and ICC-IM in the antral muscularis of W/Wvand wild-type (WT) mice was examined by confocal microscopy. The effects of XBF on the gastric antrum slow waves in W/Wvand WT mice were recorded by intracellular amplification recording. The micro-strain-gauge force transducers were implanted into the gastric antrum to monitor the MMC and the effect of XBF on gastric antrum motility in conscious W/Wvand WT mice.
Research results
In the gastric antrum of W/Wvmice, no ICC-IM network was observed. Spontaneous rhythmic slow waves with the low amplitude also appeared in the antrum of W/Wvmice. In this study, we found that the duration of MMC phase III was 151.08 ± 8.87 s,the amplitude was 315.45 ± 5.55 mg, and the interval between MMCs was 10.75 ± 0.61 min in the gastric antrum of WT mice. There was no complete MMC cycle in W/Wvgastric antrum lacking ICC-IM. The gastric antrum motility in WT and W/Wvantrum was significantly increased after treatment with XBF. Atropine blocked the enhancement of XBF completely, while tetrodotoxin partially inhibited the enhancement of XBF.
Research conclusions
In this study, we first examined the gastrointestinal motility of W-mutant mice by an extraluminal strain-gauge force transducer method. It showed that ICC-IM plays an important role in the regulation of gastric antrum MMC.
Research perspectives
In this study, MMC were recorded only at a single site in the gastric antrum. It was impossible to describe the propulsion of gastrointestinal movement. Therefore,recording at multiple gastrointestinal sites is important to further clarify the motility of gastrointestinal MMC in mice.
杂志排行

World Journal of Gastroenterology的其它文章
- Digestive system involvement of infections with SARS-CoV-2 and other coronaviruses: Clinical manifestations and potential mechanisms
- Sinapic acid ameliorates D-galactosamine/lipopolysaccharideinduced fulminant hepatitis in rats: Role of nuclear factor erythroidrelated factor 2/heme oxygenase-1 pathways
- Quantitative multiparametric magnetic resonance imaging can aid non-alcoholic steatohepatitis diagnosis in a Japanese cohort
- Clinicopathological features and prognostic factors associated with gastroenteropancreatic mixed neuroendocrine non-neuroendocrine neoplasms in Chinese patients
- Effect of liver inflammation on accuracy of FibroScan device in assessing liver fibrosis stage in patients with chronic hepatitis B virus infection
- Simultaneous partial splenectomy during liver transplantation for advanced cirrhosis patients combined with severe splenomegaly and hypersplenism