TNF-α诱导酮病奶牛脂肪组织脂解和胰岛素抵抗
2021-04-12范明赫白洪旭陈喜莹宋玉祥李心慰刘国文杜希良李小兵吉林大学动物医学学院吉林长春130062
范明赫,白洪旭,陈喜莹,宋玉祥,李心慰,刘国文,王 哲,杜希良,李小兵 (吉林大学 动物医学学院,吉林 长春 130062)
产后奶牛因干物质摄入量(DMI)下降但维持泌乳所需的营养物质明显增加,导致严重的能量负平衡(NEB)[1]。白色脂肪组织(WAT)作为主要的能量储存器官,在NEB或营养不良时能分解其存储的甘油三酯(TAG)提供脂肪酸以确保能量供应[2]。强烈和持续的NEB使脂肪组织过度分解生成大量非酯化脂肪酸(NEFA)。NEFA可被肝脏等组织代谢生成酮体,如β-羟基丁酸(BHBA),当血液中BHBA的含量超过3 mmol/L时即可诊断为临床酮病[3]。在美国的集约化牧场中,哺乳早期约有20%以上奶牛检测患有临床酮病,而在我国围产期奶牛临床酮病患病率更是可高达30%[4-5]。诸多研究表明酮病不但能降低奶牛产奶量和乳品质,而且能够增加乳房炎、子宫内膜炎和脂肪肝等其他疾病的发病风险[6]。因此,酮病是围产期奶牛疾病防控的瓶颈,而脂肪组织过度脂解是酮病发生的关键环节。
先前研究发现酮病奶牛胰岛素敏感指数低于正常奶牛,即出现胰岛素抵抗[7]。而脂肪组织是胰岛素作用的主要靶器官之一,胰岛素抵抗时表现为脂肪分解作用增强和葡萄糖摄入量减少,前者导致NEFA增多进而聚集在肝脏、肌肉等器官,NEFA具有脂毒性,能够损伤组织器官的胰岛素信号通路,进一步引起全身胰岛素抵抗并促进酮病的发生[8]。SETHI等[9]研究表明,小鼠促炎因子水平与脂肪组织的脂解速度呈正相关,而与机体的胰岛素敏感性呈负相关。本课题组先前研究发现酮病奶牛血液中促炎因子肿瘤坏死因子-α(TNF-α)、白介素(IL)-1β和IL-6浓度显著升高[10],但这是否与酮病奶牛存在过度脂肪动员以及胰岛素敏感性降低有关尚不清楚。
因此,本研究检测酮病奶牛脂肪组织脂解水平以及胰岛素敏感性变化,并探究炎性因子TNF-α对奶牛脂肪细胞脂解作用和胰岛素敏感性的影响,有助于进一步揭示奶牛酮病的发病机制,为酮病的防治提供数据支撑。
1 材料与方法
采集奶牛脂肪组织前先剪去尾根部一侧皮肤的5 cm×5 cm区域被毛并用碘酒和75%酒精消毒,皮下注射2%盐酸利多卡因麻醉。用手术刀在皮肤和皮下组织中切开1.5~2.5 cm切口,用无菌镊子及止血钳将皮肤拉开,露出组织。使用无菌手术刀刀片和镊子取出脂肪组织,用生理盐水冲洗组织样品,在液氮中快速冷冻之后,-80℃保存。
1.2 主要试剂DMEM/F12培养液、青霉素/链霉素、澳源特级胎牛血清购自Hyclone公司;牛胰岛素、地塞米松、IBMX、I型胶原酶购自Sigma公司;p-HSL、ATGL、p-IR、IR、p-Akt、Akt、β-actin抗体购自Abcam和CST公司;红细胞裂解液购自碧云天生物技术公司。
1.3 血液参数测定使用Hitachi自动分析仪以及商用试剂盒(NEFA:目录号FA115;BHB:目录号RB1008;葡萄糖:目录号GL3815;Randox Laboratories)测定采集血清中的NEFA、BHB和葡萄糖浓度。
1.4 脂肪细胞的原代分离培养将健康的1日龄荷斯坦奶牛麻醉,在无菌条件下通过外科手术方法获取腹膜网膜和肠系膜的脂肪组织。将所得脂肪组织在含有青霉素(2 500 U/mL)和链霉素(2 500 mg/L)的无菌磷酸盐缓冲盐水(PBS)中冲洗,以去除黏附组织的血液。剥离组织中可见筋膜和血块,所得组织切成约1 mm2的小块在37℃下用含胶原酶I型(1 g/L)的DMEM/F12消化液在轻微摇晃的水浴池中消化1.5 h。将消化后的混合物依次通过80和40目滤网,并将滤液在室温以175×g离心10 min,所得沉淀加入红细胞裂解液以去除残留的红细胞,再以175×g于室温离心10 min。弃去上清液,并将所得细胞再悬浮于基础培养基(BCM)中,该培养基为含10%胎牛血清和1%青霉素/链霉素的DMEM/F12。细胞计数后,将细胞悬液浓度调节至1×104/cm2,接种于细胞培养瓶中,于细胞培养箱中37°C、5%CO2的潮湿环境中培养,每48 h更换1次BCM。
为了诱导前脂肪细胞分化为成熟脂肪细胞,将细胞接种在6孔细胞培养板中。当每孔细胞数达到70%时,用分化培养基1(DCM1)代替BCM培养用以分化前脂肪细胞,DCM1是终浓度为0.5 mmol/L 3-异丁基-1-甲基黄嘌呤(IBMX)、1 μmol/L地塞米松、和1 mg/L胰岛素的BCM。培养48 h后,弃去DCM1,BCM中加入终质量浓度为1 mg/L胰岛素的分化培养基2(DCM2)代替,维持分化培养,约10 d 细胞内出现可见的脂滴,说明细胞已完全分化成熟。
1.5 细胞处理牛TNF-α(ab87904,Abcam)使用含有0.1% BSA的PBS稀释。经诱导成熟的脂肪细胞分别以0、0.1、1.0、10.0 μg/L的TNF-α处理3 h,加入含200 nmol/L胰岛素的DMEM/F12培养液作用15 min,检测胰岛素相关信号通路。
1.6 蛋白含量检测和蛋白免疫印迹使用商用蛋白提取试剂盒(C510003,生工生物技术有限公司)按照说明书从脂肪组织样本或脂肪细胞中提取蛋白。使用BCA试剂盒(P1511,普利莱有限公司)测定蛋白浓度。
蛋白免疫印迹(Western blot)检测蛋白表达,具体步骤参考DU等[1]方法。将30 μg的总蛋白质以及Marker上样到12%SDS-PAGE凝胶上,先以80 V 电泳30 min,然后再以120 V电泳45~60 min。电泳完毕后,根据目的蛋白相对分子质量大小,切取相应的胶条,以120 V,45 min将蛋白转移到0.45 μm聚偏二氟乙烯(PVDF)膜上。之后将PVDF膜放进3% BSA溶液中,在摇床室温封闭4 h。把封闭后的PVDF膜置于TBST稀释的一抗溶液中,4℃过夜孵育。一抗孵育后的PVDF膜用TBST洗涤3次,每次6 min。然后将 PVDF 膜放入对应的二抗稀释液中,室温摇床孵育45 min。二抗孵育后的PVDF膜,TBST洗涤3次,每次6 min。PVDF膜放在显影仪中,膜上均匀滴涂ECL显影液,曝光。用Image Pro Plus 6.0进行条带灰度分析。
1.7 脂肪组织总RNA提取、反转录和PCR扩增首先取约500 mg脂肪组织在研钵中加液氮研磨至粉状,研磨好的脂肪组织转移到1.5 mL无RNase的离心管,每管加入1 mL RNAiso Plus(15596026,Thermo公司),室温裂解5~10 min后—80℃冻存30 min。取出待溶解后,每管加入200 μL氯仿,上下颠倒使其充分混匀,冰上静置 5 min后,于4℃下12 000×g离心15 min。此时液体出现分层,吸取上层无色液体,转移至新的1.5 mL无RNase离心管,加入相同体积的异丙醇,轻微混匀,在冰上静置 10 min后,于4℃下12 000×g离心10 min弃掉上清,管底可见白色RNA沉淀。用DEPC水配置75%的无水乙醇,加1 mL至离心管中,洗涤RNA沉淀后,于4℃下12 000×g离心5 min。弃掉上清,在滤纸上干燥5 min,加入 20~30 μL DEPC水溶解沉淀,用N50型超微量分光光度计检测RNA浓度。使用TaKaRa 反转录试剂盒将RNA反转录成cDNA。根据测定的RNA浓度换算出单倍体系所对应的总RNA体积,参照说明书进行操作。用实时荧光定量PCR方法检测TNF-α、IL-1β mRNA的表达水平,引物信息见表1。
张伦对长辈的尊敬还会表现在对待朋友的父母上。有段时间,张伦创业失败,他决定骑着摩托车去流浪。途中经过朋友老家,想到朋友母亲已离世,而朋友久未归家扫墓,他竟跑到朋友母亲坟上祭拜。“拔草、清扫,他说弄了一个下午。”

表1 PCR引物序列
1.8 甘油(GC)含量测定根据说明书,使用商用试剂盒(E1002,普利莱有限公司)测定脂肪细胞培养基中的GC含量。收集脂肪细胞培养基,离心后在酶标微版上加入50 μL上清或标准品,再加入150 μL工作液,37℃反应10 min。使用酶标仪测定D550值。根据标准曲线计算GC浓度。

2 结果
2.1 酮病奶牛血液指标为了探究酮病奶牛的能量代谢状态,检测血液NEFA、BHBA和Glucose含量。结果发现与健康奶牛相比,酮病奶牛血清中NEFA和BHBA的浓度更高(P<0.01),相反,酮病奶牛的血糖浓度更低(P<0.01)(表2),说明酮病奶牛存在严重能量负平衡。

表2 酮病奶牛与健康奶牛血液指标(n=15) mmol/L
2.2 酮病奶牛脂肪组织脂解增强能量负平衡易诱发脂肪动员,检测酮病奶牛与健康奶牛体内脂肪组织脂解酶的表达,发现酮病奶牛p-HSL和ATGL的蛋白表达量显著增高(P<0.01)(图1A~C),表明酮病奶牛脂肪组织脂解显著增强。

图1 Western blot 检测脂肪组织p-HSL和ATGL的蛋白表达(A)及量化分析(B,C)
2.3 酮病奶牛脂肪组织胰岛素敏感性下降前期研究发现酮病奶牛血液促炎因子浓度增加,能够引起机体胰岛素抵抗。本试验结果显示,酮病奶牛脂肪组织胰岛素信号通路关键蛋白IR和Akt的磷酸化水平显著降低(P<0.01,P<0.05)(图2A~C),表明酮病奶牛脂肪组织胰岛素敏感性下降,存在胰岛素抵抗。

图2 Western blot检测脂肪组织p-IR、IR、p-Akt和Akt的蛋白表达(A)及量化分析(B,C)
2.4 酮病奶牛脂肪组织炎性因子mRNA水平增高炎性通路的过度激活是脂分解和胰岛素抵抗的潜在诱因[8-9]。结果显示,酮病奶牛脂肪组织TNF-α和IL-1β的mRNA水平升高(P<0.01),说明脂肪组织炎性反应增强,炎性因子表达量增多(图3A,B)。
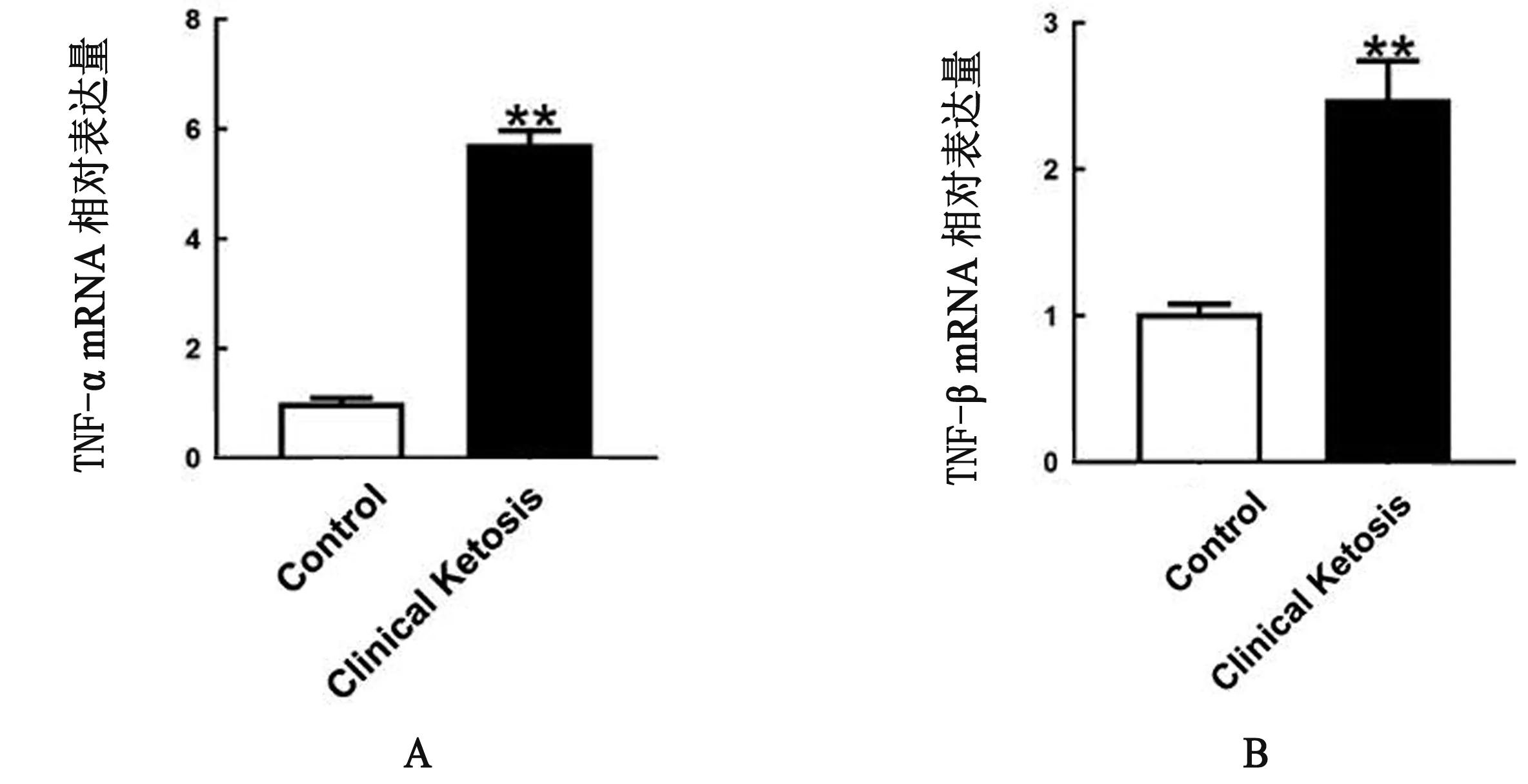
图3 奶牛脂肪组织TNF-α和IL-1β mRNA水平
2.5 TNF-α诱导奶牛脂肪细胞脂解为进一步研究酮病奶牛炎性因子增加对脂肪细胞的影响,在体外培养成熟的奶牛原代脂肪细胞中添加促炎因子TNF-α。结果显示,与对照组相比,用TNF-α(0.1、1.0和10.0 μg/L)处理后,p-HSL和ATGL蛋白丰度显著上升(P<0.01)(图4A~C)。此外脂肪细胞培养基上清中的GC含量随着TNF-α的浓度增加而上升(P<0.01,P<0.05)(图4D)。以上结果表明,促炎因子TNF-α可以显著促进脂肪细胞脂解。
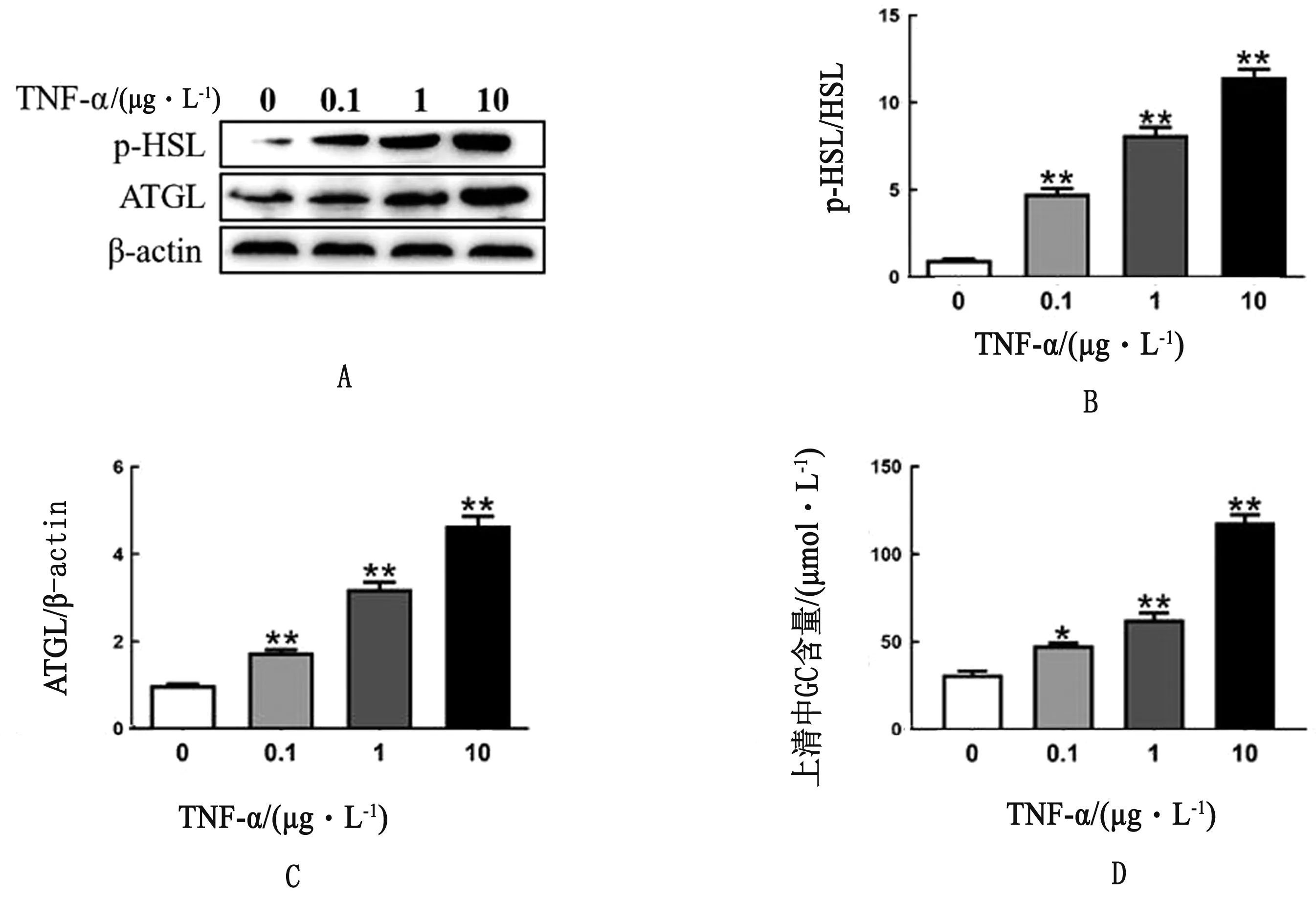
图4 Western blot 检测TNF-α处理脂肪细胞p-HSL和ATGL的蛋白表达(A)、量化分析(B,C)和培养基中GC含量(D)
2.6 TNF-α诱导奶牛脂肪细胞胰岛素敏感性下降检测TNF-α对脂肪细胞胰岛素信号通路的影响,发现TNF-α(0.1、1.0和10.0 μg/L)处理后,p-IR和p-Akt蛋白丰度显著下降(P<0.01)(图5A~C),说明促炎因子TNF-α可以显著降低奶牛脂肪细胞胰岛素敏感性,诱导胰岛素抵抗。

图5 Western blot检测TNF-α处理脂肪细胞p-IR、IR、p-Akt和Akt的蛋白表达(A)及量化分析(B,C)
3 讨论
围产期奶牛能量负平衡诱发的脂肪动员是酮病发生的病理学基础,酮病奶牛常表现为系统性炎症和胰岛素抵抗,但炎性因子能否调控酮病奶牛脂肪组织的脂解和胰岛素敏感性尚未可知。本试验结果表明,酮病奶牛脂解酶HSL和ATGL的活性和表达高于正常奶牛,但胰岛素敏感性低于正常奶牛且促炎因子TNF-α和IL-1β表达增加;脂肪细胞试验发现,TNF-α引起脂肪细胞的脂解并降低胰岛素敏感性。这些结果说明TNF-α能够诱导酮病奶牛脂肪组织脂解并降低胰岛素敏感性。
临床酮病奶牛的特点是脂肪组织过度脂解导致血液中高浓度NEFA[11]。在哺乳动物中,ATGL是脂肪分解的第1步与限速步,能将TAG水解为甘油二酯(DAG)并释放NEFA[11]。敲除ATGL小鼠与正常小鼠相比脂肪分解显著减少而且脂肪含量增多[12]。在哺乳动物体内,HSL也作为脂肪组织关键的水解酶受到可逆的磷酸化作用调节,分为有活性(磷酸化)和无活性(非磷酸化)两种形式[14]。活化的HSL能将DAG水解为甘油一酯(MAG)同时释放NEFA[15]。本试验为了确定酮病奶牛脂肪组织与健康奶牛相比脂解酶的变化情况,采用Western blot检测p-HSL与ATGL蛋白丰度,结果显示酮病奶牛脂肪组织两种酶的蛋白表达显著增加,表明酮病奶牛脂肪组织脂解增强并且HSL和ATGL起到关键的促进作用。
先前研究表明由于NEFA和BHBA的脂毒性作用,酮病奶牛和脂肪肝奶牛存在胰岛素抵抗,即胰岛素敏感组织,如脂肪、肝脏和肌肉,对胰岛素的敏感性降低[7]。脂肪细胞是炎性细胞因子的主要来源之一,而炎性因子以及NEFA的增多都将损害PI3K-Akt(磷脂酰肌醇三激酶-蛋白激酶B)胰岛素信号通路正常转导[16]。胰岛素受体(IR)属于受体酪氨酸激酶家族,在脂肪组织中胰岛素首先与胰岛素受体结合并使受体磷酸化从而激活下游胰岛素信号通路,而胰岛素信号通路的任何一步阻滞均会导致胰岛素敏感性下降[17]。通过Western blot检测发现,酮病奶牛脂肪组织磷酸化的IR和AKT蛋白丰度下调,表明酮病奶牛脂肪组织胰岛素信号通路阻滞,存在胰岛素抵抗。
脂肪组织作为一种活性内分泌器官,能够分泌包括瘦素、脂联素、TNF-α等多种脂肪因子,这些脂肪因子数量变化影响到包括脂肪组织在内的胰岛素敏感性及脂质代谢[6]。本试验发现酮病奶牛脂肪组织中促炎因子TNF-α的mRNA水平显著增高。在人类脂肪细胞中,TNF-α能够触发脂解释放NEFA的同时诱发胰岛素抵抗,说明炎症与脂解、胰岛素抵抗之间存在相互联系。先前研究曾报道过度脂肪分解产生的脂质介体能够引发3T3-L1脂肪细胞的胰岛素抵抗[18]。此外胰岛素本身能够限制脂肪组织脂解,但胰岛素抵抗导致的胰岛素敏感性下降加剧了脂肪组织进一步脂解,进而促进酮病的发生和发展。本试验通过体外培养奶牛原代脂肪细胞,添加促炎因子TNF-α模拟酮病奶牛体内炎性状态,结果显示TNF-α能够增加p-HSL与ATGL蛋白丰度和培养基中GC含量,但下调磷酸化IR和Akt的蛋白丰度,表明TNF-α可能在诱导奶牛脂肪组织脂解与胰岛素抵抗中起到重要作用。
综上所述,酮病奶牛脂肪组织存在过度脂解与胰岛素抵抗,并且炎性因子TNF-α能够诱导脂肪分解并损害胰岛素信号通路。因此本试验有助于进一步了解奶牛酮病发生的潜在机制,为预防酮病发生提供新的思路。
