羊传染性脓疱病毒通过激活PI3K/Akt通路抑制HeLa细胞凋亡
2021-04-12吕丽君黄后双关继羽周艳龙贺文琦宋德光吉林大学动物医学学院吉林长春30062深圳市药品检验研究院广东深圳58057
吕丽君,黄后双,2,关继羽,周艳龙,王 帅,贺文琦,宋德光,高 丰,赵 魁* (.吉林大学 动物医学学院,吉林 长春 30062;2.深圳市药品检验研究院,广东 深圳 58057)
羊传染性脓疱病毒(Orf virus,ORFV)为痘病毒科(Poxviridae)副痘病毒属(Parapoxvirus)成员,具有高度的嗜上皮性,主要引起感染绵羊、山羊的局部性皮肤损伤病变[1-2]。通过与病羊的直接/间接接触,人(尤其是牧民、屠夫、兽医)也可发生感染,成为影响我国乃至世界公共卫生安全的一个重要隐患[3-4]。目前,ORFV感染广泛分布于世界各地,在我国以东北、西北等主要养羊省区羊群中发生较为严重,是严重危害养羊业发展的重要动物疫病之一。尽管该病临床上具有自限性和低死亡率的特点,但在羔羊、儿童和免疫抑制动物,死亡率可高达80%[5]。
细胞凋亡是宿主细胞对抗病毒感染等多种外界刺激所做出的重要防御手段之一,通过诱导感染细胞凋亡从而有效阻止病毒对邻近细胞的感染,进而抑制病毒的复制和传播[6-7]。然而,作为大型DNA病毒,痘病毒自身能够复制编码一系列抗凋亡基因抑制感染细胞的凋亡[8],例如,ORFV ORF125基因被发现含有与Bcl-2家族成员相同的保守序列,编码蛋白具有类似Bcl-2的抗凋亡活性[9]。ORFV ORF119蛋白C端存在视网膜母细胞瘤蛋白(pRb)结合基序“LxCxE”,可通过与pRb家族成员的结合,进而参与对细胞周期、细胞凋亡等通路的调控[10]。
近年来,越来越多的研究表明,多种病毒可激活宿主细胞PI3K/Akt等抗凋亡通路,通过抑制细胞凋亡从而有助于病毒完成生命周期[11]。例如,单纯疱疹病毒1(herpes simplex virus,HSV-1)能够激活与凋亡阻断相关的PI3K/Akt信号通路,促进病毒基因的表达[12]。乙肝病毒表面抗原(HBxAg)可通过激活PI3K/Akt通路抑制人胎盘滋养层细胞凋亡[13]。鸡传染性法氏囊病病毒(infectious bursal disease virus,IBDV)通过其VP5蛋白与PI3K的p85α亚基相互作用,激活PI3K/Akt信号通路,抑制感染早期细胞的凋亡,进而促进病毒增殖[14]。肠道病毒71型(entero virus 71,EV71)也可通过激活 PI3K/Akt 通路抑制JNK介导的细胞凋亡[15]。然而,在ORFV感染过程中,PI3K/Akt信号通路与细胞凋亡的相互关系尚不明确。鉴于此,本研究运用Western blot、免疫荧光技术、流式细胞术、药物阻断等方法,系统研究ORFV感染对PI3K/Akt通路和细胞凋亡的影响,以及PI3K/Akt通路介导ORFV感染HeLa细胞凋亡抑制的途径。该研究不仅有助于加深对ORFV抑制感染细胞凋亡机制的了解,而且将为ORFV免疫逃逸机制的全面解析以及疾病防控提供重要的理论依据。
1 材料与方法
1.1 细胞和病毒羊传染性脓疱病毒(ORFV/JL/08/CC)由本实验室分离鉴定保存[16],HeLa细胞由本实验室保存。
1.2 抗体和试剂PI3K抑制剂LY294002购自Calbiochem公司;Akt、磷酸化Akt(Ser473)和β-actin抗体均购自CST公司;MDM2、GSK3β和Bad抗体均购自Abcam公司;辣根过氧化物酶(HRP)标记山羊抗小鼠IgG和山羊抗兔IgG购自Proteintech公司;MEM培养基和胎牛血清均购自GIBCO公司;MTT购自Sigma公司;annexin V-FITC/PI 凋亡检测试剂盒购自北京全式金生物。
1.3 细胞增殖试验利用MTT法测定细胞增殖能力,将96孔板细胞用不同浓度(10~50 μmol/L)的PI3K抑制剂LY294002处理48 h后,每孔加入MTT试剂使终质量浓度为0.5 g/L,37 ℃、5% CO2孵育4 h;去除培养基,加入DMSO(100 μL/孔)摇板孵育10 min。使用酶标仪(BioRad,Hercules,CA)测定活细胞在570 nm波长下的吸光度。
1.4 病毒感染与药物处理经PI3K抑制剂LY294002(25 μmol/L)预处理1 h后的Hela细胞,以MOI=1接种ORFV,或以等体积的MEM作为对照,吸附1 h后,加入含有相同浓度LY294002的MEM共培养。
1.5 免疫荧光分析使用annexin V-FITC/PI凋亡检测试剂盒,按照说明书对细胞进行凋亡检测;染核:细胞经PBS洗涤3次后,用预冷的甲醇和丙酮(1∶1)固定10 min,与Hoechst(0.5 g/L)溶液共孵育10 min,再次洗涤后,使用倒置荧光显微镜(Leica DM18,Germany)观察细胞。
1.6 流式细胞术检测将细胞按1.4方法处理后,用胰蛋白酶消化收集并重悬于annexin V结合缓冲液中,使细胞浓度为1×106个/mL。取100 μL细胞悬液(105个细胞)与annexin V-FITC(0.1 g/mL)混合室温孵育15 min后,使用500 μL结合缓冲液重悬并与10 μL的PI(30 g/mL)溶液4 ℃避光染色。利用BD FACSCalibur流式细胞仪(Becton Dickinson)检测annexin V-FITC荧光,每个样本至少分析30 000个细胞。
1.7 Western blot检测将细胞按1.4方法处理后,使用含有1 mmol/L PMSF的蛋白裂解液收集样品,经10% SDS-PAGE电泳结束后转移到PVDF膜。含5%脱脂奶的TBST(0.1% 吐温20)溶液37℃封闭1 h后,分别用Akt、p-Akt(Ser473)、MDM2、GSK-3β和β-actin抗体4℃孵育过夜,洗膜后进行HRP标记二抗孵育、ECL显色,并利用Image J软件对蛋白进行定量。
1.8 统计学分析通过SPSSV 17.0软件对结果进行t检验分析(SPSS Inc.,Chicago,IL,USA)。*P<0.05,**P<0.01,***P<0.001,差异有统计学意义。
2 结果
2.1 ORFV感染诱导依赖PI3K的Akt磷酸化Western blot检测ORFV感染Hela细胞后Akt和p-Akt的表达情况。结果显示,ORFV感染组中p-Akt(Ser 473)水平在3~48 h间先增加后减少,48 h 后p-Akt的水平与对照组细胞同一时间点的表达相似(图1A)。由于Akt是PI3K信号通路下游的主要效应分子,因此,进一步探究ORFV感染中Akt的活化是否通过PI3K信号通路实现。将ORFV接种于经PI3K特异性抑制剂LY294002(25 μmol/L)预处理1 h的HeLa细胞,吸附1 h后加入含有相同浓度LY294002的MEM共培养,收集感染后的细胞裂解液,Western blot检测p-Akt水平。结果显示,经LY294002处理Akt的总蛋白水平未发生明显改变,但能够抑制Akt的磷酸化(图1B)。该结果表明,ORFV 感染Hela 细胞能够激活依赖PI3K的Akt磷酸化。此外,MTT法检测结果显示,LY294002浓度达40 μmol/L时对Hela细胞增殖具有显著影响(图2)。
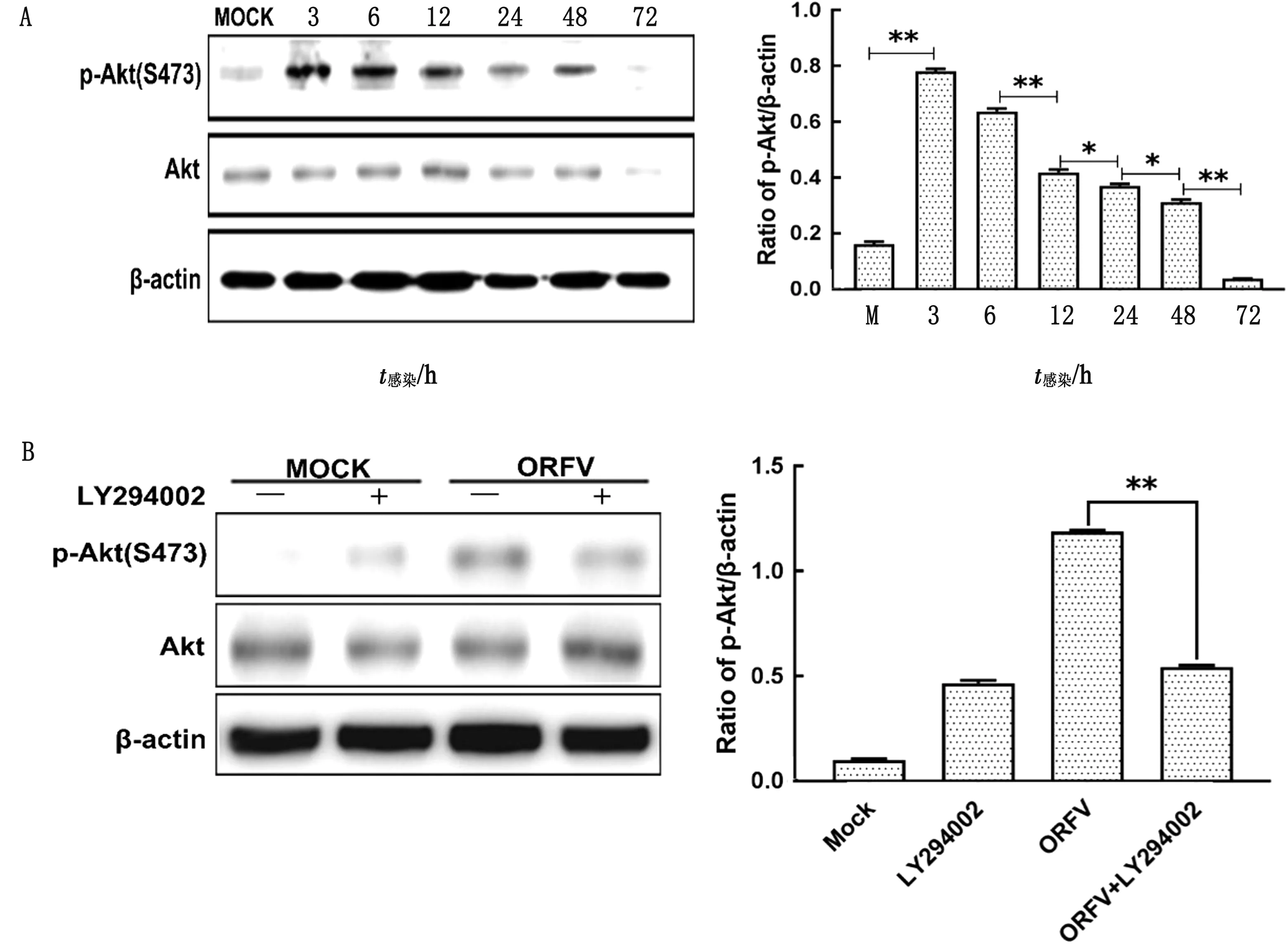
A.ORFV感染Hela细胞诱导Akt发生磷酸化;B.ORFV感染Hela细胞诱导依赖PI3K的Akt磷酸化

图2 MTT法检测PI3K特异性抑制剂LY294002对Hela细胞活性的影响
2.2 ORFV感染通过PI3K/Akt信号通路抑制Hela细胞凋亡将ORFV接种于Hela细胞,并以等量MEM作为对照,48 h后使用细胞核染色剂Hoechst 33258和Annexin V-FITC/PI 对细胞进行染色。荧光显微镜观察发现,在ORFV感染组中,Hela细胞核完整,未出现染色质浓缩、分割或边缘化,细胞膜发生裂解成块状等典型的凋亡状态,Annexin V未出现特异性染色,与对照组一致(图3A),表明ORFV感染未引起Hela细胞发生凋亡。将ORFV接种于经PI3K特异性抑制剂LY294002(25 μmol/L)预处理1 h的Hela细胞,吸附1 h后加入含有相同浓度LY294002的MEM共培养,并以等量DMSO作为对照,48 h后使用Hoechst 33258和Annexin V-FITC/PI 对细胞进行染色。结果显示,与DMSO对照组相比,LY294002处理组Hela细胞核出现蓝色浓斑颗粒状光、体积变小,细胞质浓缩,Annexin V-FITC标记绿色荧光增多,同时PI也使细胞核发生了弥散性的红染(图3B),表明经LY294002预处理抑制PI3K/Akt信号通路后,ORFV感染诱导Hela细胞发生凋亡。上述结果表明,PI3K/Akt 信号通路在ORFV感染Hela细胞的凋亡抑制过程中起着重要作用。此外,将ORFV接种于经PI3K特异性抑制剂LY294002(25 μmol/L)预处理1 h的Hela细胞,吸附1 h后加入含有相同浓度LY294002的MEM共培养,并以等量ORFV单独接种作为对照,使用Annexin V-FITC/PI双标试剂盒对LY294002和ORFV共培养24、48 h的Hela细胞进行染色标记,利用流式细胞仪检测凋亡情况。结果显示,与ORFV感染组相比,LY294002处理组细胞凋亡率明显增高,凋亡细胞数量显著增多(图4A,B)。综上所述,ORFV感染通过激活PI3K/Akt信号通路抑制Hela细胞凋亡。

A.ORFV感染Hela细胞;B.ORFV感染经PI3K抑制剂LY294002预处理的Hela细胞

A.ORFV感染Hela细胞;B.ORFV感染经LY294002预处理的Hela细胞
2.3 PI3K/Akt通过激活其下游效应分子Bad抑制Hela细胞凋亡将ORFV接种于经PI3K特异性抑制剂LY294002(25 μmol/L)预处理1 h的Hela细胞,吸附1 h后加入含有相同浓度LY294002的MEM共培养,Western blot检测6、24 h后MDM2、GSK-3β、Bad的表达情况。结果显示,与对照组相比,当PI3K被LY294002抑制后,ORFV感染Hela细胞MDM2和GSK-3β分子无显著变化,但Bad表达量显著增加(图5A,B)。结果表明Bad分子参与了ORFV感染Hela细胞的抗凋亡过程。
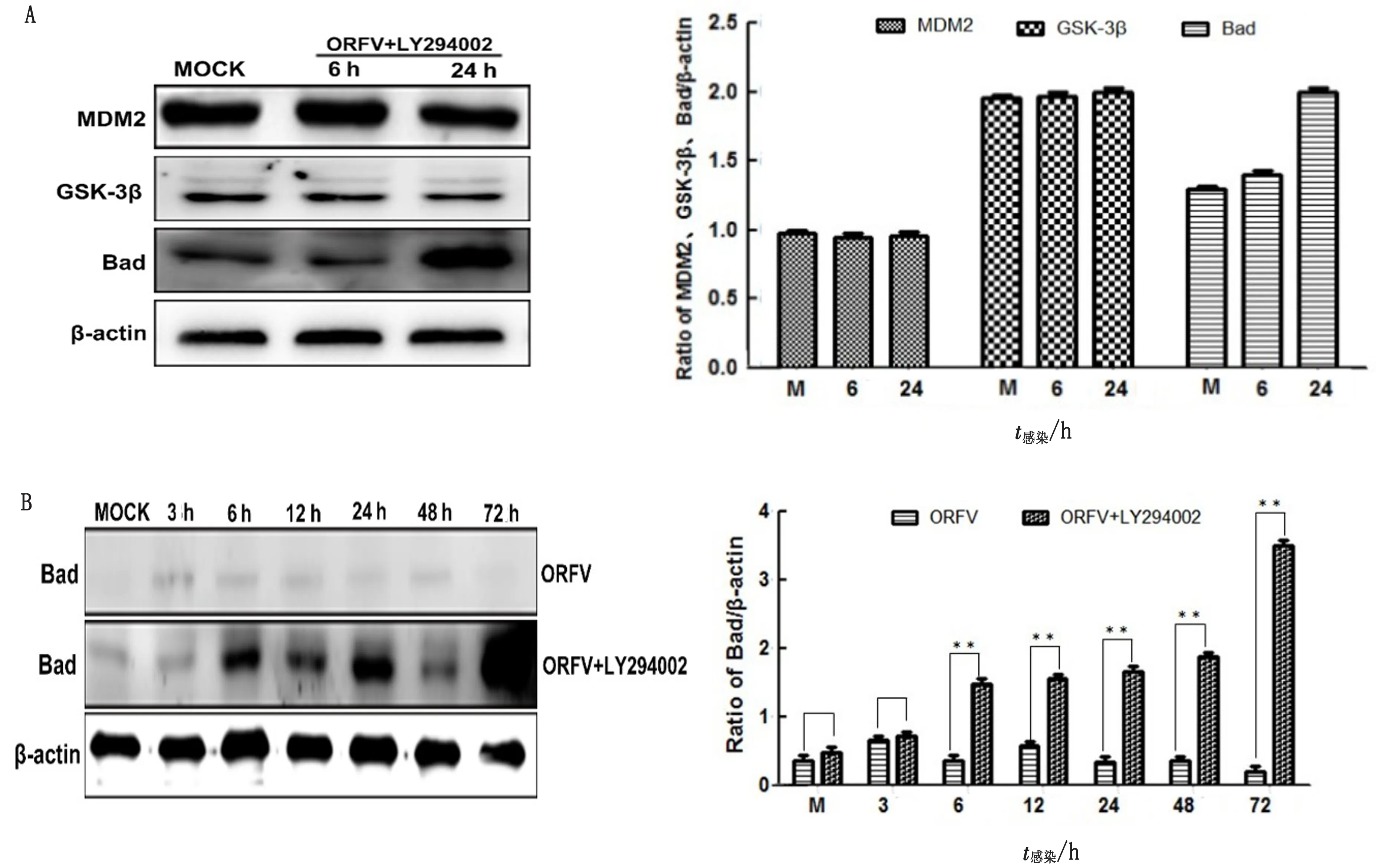
A.PI3K/Akt下游效应分子MDM2、GSK-3β、Bad表达;B.ORFV感染Hela细胞Bad分子表达
3 讨论
许多病毒感染宿主细胞后,能够破坏宿主细胞的防御系统以确保自身的生存、复制和增殖,从而达到持续性感染的目的。细胞凋亡是一种细胞程序性死亡,以细胞收缩、核浓缩和浆膜起泡为特征,在组织发育、体内平衡和各类疾病中发挥着至关重要的作用,是宿主对病毒感染反应的关键组成部分。研究表明,大量病毒已经进化出复杂的分子策略来破坏宿主细胞的凋亡防御。在宿主细胞部署的各种防御机制中,一些依赖于细胞程序性死亡的途径可被快速激活。例如,许多病毒获得了Bcl-2的同源物来破坏宿主细胞的凋亡和自噬,以防止被感染细胞的过早死亡,从而影响宿主与病毒的相互作用,最终使病毒感染成功建立和传播。
研究发现,痘病毒中1/3~1/2的病毒基因组致力于免疫逃避,其中多种痘病毒产物参与拮抗细胞凋亡反应,从而使病毒完成其生命周期并产生子代病毒粒子。如牛痘病毒(VACV)的B13和B22蛋白能够抑制Caspase(半胱天冬酶)活性从而抑制凋亡;猪痘病毒(SPV)的032蛋白抑制PKR激活能够减少或阻止Caspase-7和PARP-1的裂解。研究表明,ORFV作为痘病毒科副痘病毒属的一员,其编码的ORF125抗凋亡基因可抑制感染细胞的凋亡。实验室前期研究发现,参与细胞增殖、分化和凋亡调控相关通路的重要分子PI3K对ORFV的复制增殖具有明显的促进作用,然而具体的机制尚不清楚[22]。
本研究深入探究ORFV感染对PI3K/Akt通路和细胞凋亡的影响,以及PI3K/Akt通路介导ORFV感染Hela细胞凋亡抑制的途径。研究发现,ORFV感染Hela细胞能够激活p-Akt(Ser473),而PI3K特异性抑制剂LY294002处理则使p-Akt表达显著降低,说明ORFV感染Hela细胞能够激活PI3K/Akt信号通路。此外,免疫荧光及流式细胞术检测结果显示,ORFV感染未引起Hela细胞凋亡,但PI3K/Akt信号被LY294002抑制后,细胞出现了明显的凋亡现象,表明ORFV感染能够通过激活PI3K/Akt信号通路抑制Hela细胞发生凋亡。进一步对PI3K/Akt下游效应分子MDM2、GSK-3β、Bad表达情况检测发现,经PI3K/Akt通路特异性抑制剂LY294002预处理后,ORFV感染Hela细胞MDM2、GSK-3β表达均无显著变化,仅Bad分子表达显著增加,表明Bad分子可能参与ORFV感染Hela细胞的抗凋亡过程。综上所述,ORFV感染可通过激活PI3K/Akt/Bad信号抑制Hela细胞凋亡。该研究结果不仅有助于解析ORFV抑制感染细胞凋亡的机制,而且将为全面阐明ORFV免疫逃逸机制及疾病防控提供重要的理论依据。
