舞毒蛾取食诱导小黑杨苯丙烷代谢酶活性及其相关基因的表达*
2021-04-10周心怡闫丽琼吕云彤孙丽丽朱靖闻曹传旺
周心怡 闫丽琼 吕云彤 孙丽丽 朱靖闻 曹传旺
东北林业大学森林生态系统可持续经营教育部重点实验室 哈尔滨 150040)
植物和昆虫在地球上共存已有数亿年,在长期的进化过程中,双方形成了一系列相互防御的策略(陈澄宇等, 2015)。虫害诱导的植物防御反应及其复杂,不同植物、不同刺激方式和强度下的防御反应机制不同(张斌等, 2016)。1980年,Bell等(1980)提出了“植物次生代谢”概念,植物次生代谢产物是指植物中一大类对于细胞生命活动或植物生长发育正常进行并非必需的小分子有机化合物,其在植物体内含量不等,且均有自己独特的代谢途径,通常由初生代谢派生而来(Dixon, 2001)。参与植物防御反应的次生代谢产物主要有酚类、萜类和生物碱类,其对植食性昆虫具有引诱、驱避、毒杀、拒食和不育等作用(王景顺等, 2015),苯丙氨酸是许多植物化合物的前体,对植物繁殖、生长、发育和防御不同类型的胁迫至关重要,苯丙氨酸代谢途径是植物体内最重要的次生代谢途径之一(陈晓亚等, 1996),能够将碳源大量转移到苯丙氨酸衍生化合物的生物合成中,特别是木质素(Pascualetal., 2016)。在高等植物中,肉桂酸-4-羟化酶(C4H)、苯丙氨酸氨裂解酶(PAL)和4-香豆酸CoA连接酶(4CL)参与苯丙烷新陈代谢过程中的核心反应(Mizutanietal., 1993)。C4H行使功能需氧且依赖NADPH,催化苯丙氨酸途径的第二步反应,同时也是该途径的第一个氧化反应,在肉桂酸的对位点上催化位置特异性的羟化反应,将反式肉桂酸催化生成对-香豆酸(李莉等, 2007)。目前,在水稻(Oryzasativa)、杜仲(Eucommiaulmoides)和青稞(Hordeumvulgare)等植物中已经克隆鉴定出C4H,并对其表达特性表达了分析(罗小娇等, 2014; 李铁柱等, 2014; 刘亚洲等, 2019)。4CL作用于苯丙氨酸代谢途径中最后一步反应,催化各种羟基肉桂酸生成相应的硫酯,这些硫酯处于苯丙酸代谢途径和各种末端产物特异合成途径的分支点。4CL主要以香豆酸、咖啡酸和阿魏酸为催化底物,通常以基因家族形式存在(李莉等, 2007),进而调控木质素、类黄酮等次生代谢物质的形成(饶国栋等, 2012)。研究发现,拟南芥(Arabidopsisthaliana)的11个4CL重组蛋白中,4个在体外存在催化活性,具有较广的底物特异性(Costaetal., 2005);水稻中在功能上存在分化的5个4CL基因(Guietal., 2011);毛果杨×美洲黑杨(Populustrichocarpa×P.deltoides)杂交杨仅有2个4CL基因,且同源性很高,但功能各异(Allinaetal., 1998)。然而,植物苯丙氨酸代谢途径调控次生物质响应病虫害胁迫的基因功能亟待进一步研究。
舞毒蛾(Lymantriadispar)属鳞翅目(Lepidoptera)毒蛾科(Liparidae)毒蛾属(Lymantria),是一种世界性森林食叶害虫,其分布广、食性杂,可取食500多种寄主植物,给农林业生产造成了巨大损失(Lazarevicetal., 1998)。在食叶害虫与寄主植物的互作中,寄主植物的次生代谢产物起着关键的“化学防御”作用,次生代谢产物大多是非挥发性物质,影响昆虫对食物的选择、利用和消化进程,延缓发育。据报道,分月扇舟蛾(Closteraanastomosis)取食虫害处理的美洲黑杨后,幼虫各发育历期延长,对食物的利用率下降,死亡率升高(周艳琼等, 2010);多种信号途径参与昆虫取食诱导的植物防御反应,其相互作用为协同或拮抗(秦秋菊等, 2005)。小黑杨(Populussimonii×P.nigra)是从小叶杨(P.simonii)与欧洲黑杨(P.nigra)杂交组合中选出的优良单株,具有生长速度快、抗寒、抗旱、抗病虫害等优良特性(沈清越等, 1979)。本研究基于小黑杨转录组文库中获得的C4H和4CL基因序列,测定舞毒蛾取食和机械损伤胁迫下苯丙氨酸代谢途径中关键基因C4H、4CL的表达量以及酶活性变化情况,从转录和蛋白水平上揭示小黑杨诱导抗性的形成和发展规律,以期为明确杨树抗虫性分子机制提供理论依据,并为利用基因工程手段提高寄主抗虫性提供基因材料。
1 材料与方法
1.1 供试材料
舞毒蛾卵块和人工饲料购自中国林业科学研究院森林生态环境与保护研究所。将舞毒蛾卵块包裹于纱布中,使用体积分数为10%为甲醛溶液浸泡1 h,用蒸馏水漂洗,打开纱布,晾干卵块表面水分后将卵块放入塑料培养盒(直径9 cm),置于温度(25 ±1)℃、光照14L∶10D、相对湿度75%的人工培养箱内,并及时将新孵出的舞毒蛾幼虫转入盛有人工饲料的培养盒中。选取健康、大小一致的4龄幼虫进行试验。
小黑杨植株由东北林业大学林木遗传育种国家重点实验室提供。参照刘桂丰等(2002)方法,待组培幼苗在培养瓶中长出完整根系后,打开瓶盖,置于塑料大棚炼苗3~4天。从瓶中取出幼苗,小心洗去附着在根部的培养基,种植于经过除菌剂代森锌(80%可湿性粉剂,沈阳农药有限公司生产,质量浓度4 g·L-1)处理3天后的草炭土与沙子等量混合的基质中,置于相对湿度60%~70%的塑料大棚遮荫培养(姜静等, 2004)。选用平均直径18.31 mm±2.15 mm、平均株高110.27 cm±3.79 cm的植株进行胁迫处理。
1.2 C4H和4CL基因克隆与分析
采用CTAB法提取健康小黑杨的总RNA(曾凡锁等, 2007),用于构建转录组文库,采用IIlumina HiSeqTM2000进行测序(深圳华大基因科技有限公司)。C4H和4CL基因克隆与分析参考刘鹏等(2017)方法,首先利用NCBI中Blastx和Blastn分析转录组文库中的Unigenes 功能,根据注释结果,查找并获得C4H和4CL基因序列,然后设计特异性引物进行RT-PCR验证,通过测序获得C4H和4CL基因全长序列。开放阅读框采用ORF Founder (https:∥www.ncbi.nlm.nih.gov/orffinder)程序确定,蛋白质分子质量和理论等电点用ProtParam (http:∥au.expasy.org/tools/protparam.html) 软件计算推导,利用SignlP4.1 Server (http:∥www.cbs.dtu.dk/services/SignalP)分析氨基酸序列中信号肽,运用NCBI的Conserved Domains程序(https:∥www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)预测蛋白保守区,通过Blast (http:∥www.ncbi.nlm.nih.gov/BLAST/)进行序列同源性搜索,选择与其相似程度高的不同植物C4H和4CL氨基酸序列,用ClustalW进行多序列比对。应用Clustalx (1.83)和MEGA5.1,采用邻接法(neighbor-joining, N-J)构建系统发育树(Brodskyetal., 1993)。
1.3 舞毒蛾取食和机械损伤处理
试验于2018年7月下旬进行。选用株高一致、无病虫害的健康小黑杨植株,随机选择健康、大小一致的舞毒蛾4龄幼虫(不区分雌雄)。设计舞毒蛾取食处理(采用每株放置30头舞毒蛾幼虫,使其在植株上均匀分布)、机械损伤处理(高压蒸汽灭菌剪刀仿照舞毒蛾取食造成的损伤形状对小黑杨叶片进行机械损伤处理)和对照(未进行任何损伤)3种处理, 处理期间所有植株均用100网目尼龙纱网套笼,以防止其他昆虫取食和舞毒蛾幼虫逃逸。根据舞毒蛾幼虫取食小黑杨情况,处理24 h后,剪取全株叶片作为测试样本,并迅速放入液氮中,带回置于-80 ℃冰箱内备用。
1.4 C4H和4CL活性测定
C4H活性按照试剂盒说明书(北京雷根公司)测定。从冰箱中取0.5 g待测样品,加入2 mL C4H Lysis Buffer,冰浴下玻璃匀浆器充分匀浆,4 ℃下 10 000 r·min-1离心15~20 min,取上清液,-20 ℃冻存,用于C4H活性测定。测定管中加入1.64 mL蒸馏水、0.05 mL酶液、0.25 mL C4H Lysis Buffer、NADPH工作液(取试剂盒中1支NADPH加入1 mL蒸馏水充分溶解,即为NADPH工作液)和0.05 mL C4H Assay Buffer,对照管中不加入C4H Lysis Buffer,其他成分均与测定管相同。以对照管调零,立即用分光光度计(比色杯光径为1 cm)测定OD 290 nm处吸光度。37 ℃准确孵育1 h,立即用分光光度计测定OD 290 nm处吸光度。以每分钟每毫克蛋白变化0.01OD为1个酶活性单位(U)。
4CL活性参照Knobloch等(1977)方法测定。准确称取1 g待测样品,加入预冷提取液(含50 mmol·L-1Tris-HCl,pH 8.9,15 mmol·L-1β-巯基乙醇,5 mmol·L-1EDTA,5 mmol·L-1Vc,10 mmol·L-1leupeptin,1 mmol·L-1PMSF,0.15% PVP,30%甘油),冰浴下玻璃匀浆器充分匀浆,4 ℃下 4 000 r·min-1离心15 min,取上清液作为4CL粗酶液。测定管反应体系为0.45 mL 15 μmol·mL-1Mg2+、0.15 mL 5 μmol·mL-1香豆酸、0.15 mL 50 μmol·mL-1ATP、0.15 mL 1 μmol·mL-1CoA和0.5 mL酶液,对照管同等条件下以0.15 mL ddH2O替代香豆酸。于40 ℃下反应10 min,波长333 nm处测定吸光度,以每分钟每毫克蛋白变化0.000 1OD为1个酶活性单位(U)。蛋白质含量测定参照Bradford(1976)的考马斯亮蓝G-250法。
1.5 实时荧光定量RT-PCR
随机挑选舞毒蛾取食组、机械损伤组和对照组的小黑杨叶片,采取CTAB法提取总RNA(曾凡锁等, 2007),用DNase Ⅰ(Promega)消化总RNA中的DNA,测定质量浓度,采用PrimeScriptTMRT reagent Kit(TaKaRa)合成cDNA,将cDNA稀释10倍,作为模板备用,使用试剂盒TransStart Tip Green qPCR SuperMix(TransGen)进行实时荧光定量RT-PCR。内参基因(EF1β和UBQ)、C4H-1、C4H-2和4CL基因引物序列见表1。实时荧光定量RT-PCR反应体系为10 μL 2 ×TransStart Tip Green qPCR SuperMix酶、正向和反向引物(10 μmol·L-1)各1 μL、2 μL稀释的cDNA,加去离子水补足20 μL; 反应条件为94 ℃预变性30 s,94 ℃变性5 s,60 ℃退火15 s,72 ℃延伸10 s,81 ℃读板1 s,45个循环,每处理重复3次,采用2-ΔΔCt方法进行基因相对表达量分析(吕云彤, 2020; Pfaffletal., 2002)。

表1 实时荧光定量RT-PCR引物序列
2 结果与分析
2.1 小黑杨C4H和4CL基因特性分析
从小黑杨转录组文库中克隆鉴定获得小黑杨C4H-1、C4H-2和4CL基因,全长基因开放阅读框大小为1 518~1 740 bp,编码505~579个氨基酸,蛋白分子质量为58.00~63.03 kDa,理论等电点pI为5.63~9.09,C4H-1和C4H-2为碱性蛋白,4CL为酸性蛋白(表2)。Blastp对3个基因保守区的预测结果表明,C4H-1和C4H-2蛋白属于p450家族基因蛋白,4CL属于多结构域蛋白超家族(AFD-class-Ⅰ)。
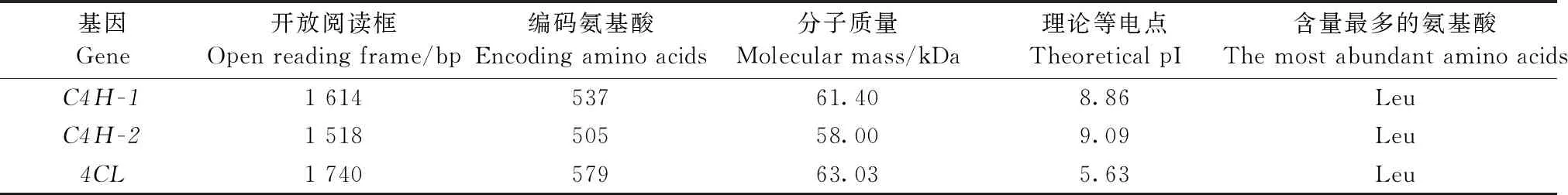
表2 小黑杨C4H和4CL基因特性
通过Blastp进行序列同源性搜索,选择与小黑杨C4H-1、C4H-2同源的35种植物的C4H蛋白以及与小黑杨4CL同源的27种植物的4CL蛋白进行多序列比对,分别构建C4H和4CL系统进化树。系统进化树分析表明,小黑杨C4H-1与银白杨(P.alba)TKR74890.1同源性为98.87%,亲缘关系近聚为一类, C4H-2与毛白杨(P.tomentosa)APR63996.1、欧洲山杨(P.tremuloides)ABF69 102.1、毛果杨×美洲黑杨AAG50231.1和毛果杨XP002319974.1具有较高同源性聚为一类(图1);小黑杨4CL与毛果杨×美洲黑杨AAK58909.1和毛果杨XP002325815.1亲缘关系较近(图2)。

图1 小黑杨C4H蛋白与35种植物C4H蛋白的系统进化树
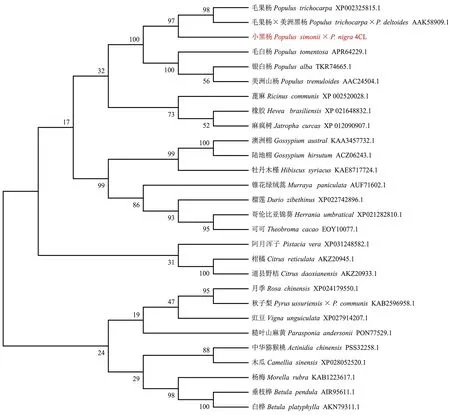
图2 小黑杨4CL蛋白与27种植物4CL蛋白的系统进化树
2.2 舞毒蛾取食和机械损伤对小黑杨C4H和4CL基因表达量的影响
如图3所示, 舞毒蛾取食诱导小黑杨C4H-1、C4H-2和4CL基因表达量增加; 机械损伤诱导小黑杨C4H-1基因表达上调,而对C4H-2和4CL基因表达下调。舞毒蛾取食24 h后,小黑杨C4H-1、C4H-2和4CL基因相对表达量分别为对照组的6.86、1.15和1.50倍; 机械损伤24 h后,小黑杨C4H-1、C4H-2和4CL基因相对表达量分别为对照的1.80、0.71和0.60倍。
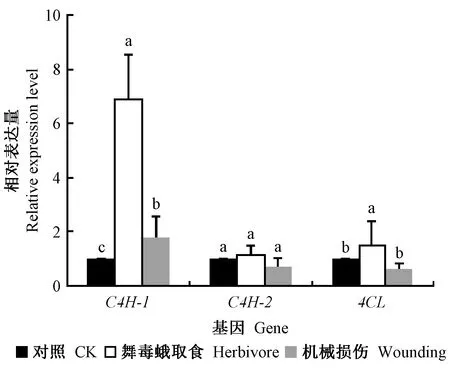
图3 舞毒蛾取食和机械损伤对小黑杨C4H和4CL
2.3 舞毒蛾取食和机械损伤对小黑杨C4H和4CL活性的影响
如图4所示,舞毒蛾取食C4H活性为748.60 U·mg-1,是对照的16.69倍; 4CL活性为2 610.36 U·mg-1,是对照的17.05倍。机械损伤C4H活性为219.19 U·mg-1,是对照的4.89倍; 4CL活性为1 196.93 U·mg-1,是对照的7.82倍。虽然舞毒蛾取食和机械损伤均能诱导小黑杨C4H和4CL活性,但取食的诱导作用显著高于机械损伤,分别为机械损伤的3.42和2.18倍。
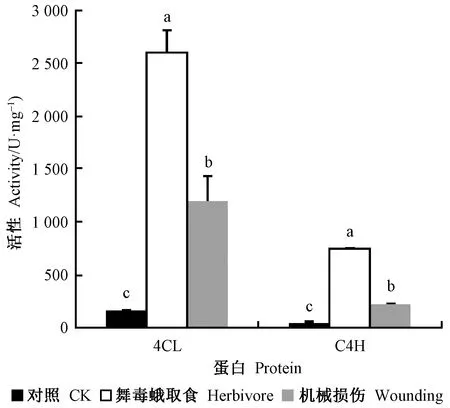
图4 舞毒蛾取食和机械损伤对小黑杨C4H和4CL活性的影响
3 讨论
植物在进化过程中对昆虫和病原菌危害以及植食性动物取食形成了多种防御机制,一般可分为组成型防御机制和诱导型防御机制,其中诱导防御机制是植物防御侵害过程中的主要机制(Waretal., 2012)。植物防御外界侵害时通常会产生包括蛋白酶抑制剂、氧化酶系、苯丙烷类代谢途径酶、糖结合蛋白和病程相关蛋白等一系列防御蛋白(秦秋菊等, 2005),植物苯丙氨酸生物合成代谢途径在逆境胁迫条件下,相关酶基因的转录水平会发生一系列改变,以诱导相应化合物的合成与积累(Fahrendorfetal., 1993; 陈鸿翰等, 2013)。Chaman等(2013)研究发现蚜虫侵染大麦(Hordeumvulgare)后会引起苯丙氨酸酶(phenyalanine ammonia-lyase,PAL)活性升高,且在具有抗蚜虫作用的大麦中PAL活性高于敏感品种。Cao等(2011)研究表明,小黑杨被舞毒蛾取食和机械损伤后PAL活性和mRNA表达水平增加。木质素是植物防御害虫取食的重要物质,C4H和4CL是木质素生物合成途径中的关键酶,通过调控C4H和4CL基因表达量可以影响木质素的合成量(李伟等, 2003);同时,木质素与细胞木质化以及植物损伤部位伤口愈合密切相关,能够间接参与植物对植食性害虫的防御作用(苏晶等, 2014)。马尾松(Pinusmassoniana)受到害虫取食后,产生的木质素对植食性害虫具有毒杀作用(任琴等, 2007)。
苯丙氨酸在PAL作用下形成反式肉桂酸,经C4H和4CL催化的一系列反应后成木质素单体,最后木质素单体聚合形成木质素。木质素作为植物防御外界侵害的物理屏障,充实细胞壁的同时也增强了细胞壁抗真菌穿透和抗酶溶解的能力(郭艳玲等, 2012)。本研究表明,小黑杨C4H-1和C4H-2属于p450家族基因蛋白,4CL属于多结构域蛋白超家族(AFD-class-Ⅰ)。舞毒蛾取食对小黑杨C4H-1、C4H-2和4CL基因表达量的影响主要表现为诱导上调,机械损伤对小黑杨C4H-1基因表现为诱导上调,而对C4H-2和4CL基因主要表现为抑制其表达下调。此外,舞毒蛾取食和机械损伤均能诱导C4H和4CL活性增加且舞毒蛾取食的活性变化显著高于机械损伤,这表明小黑杨机体可能通过增加木质素合成途径中C4H和4CL的活性以提高木质素含量来抵御损伤逆境胁迫。已有研究表明,木质素尽管是组成型的结构物质,但具有相当强的受诱导合成特性(Buchananetal., 2015)。
小黑杨对舞毒蛾取食和机械损伤处理表现出不同的应激反应,这可能是由于舞毒蛾唾液中存在的效应因子在调控植物防御反应中发挥重要作用,而机械损伤仅仅是一个物理因素。研究发现,在昆虫与植物的互作过程中,植食性昆虫的唾液起重要作用。植物识别不同种昆虫唾液中的特异性激发子从而对其防御反应进行精准调控(禹海鑫等, 2015)。甜菜夜蛾(Spodopteraexigua)口腔内的分泌物N-(17-羟基亚麻酸基)-L-谷氨酸可诱导植物释放挥发性信号物质,其中包括可以吸引天敌的诱导素,但机械损伤单独诱导不能产生此类诱导素(Albometal., 1997)。进一步研究,昆虫的唾液还能降解植物防御次生代谢物质,如水稻褐飞虱(Nilaparvatalugens)唾液腺中的一种纤维素酶内切-β-1,4-葡聚糖酶(endo-β-1,4-glucanase,NIEG1),能够帮助褐飞虱适应水稻细胞壁的物理屏障,同时可以水解细胞壁中的纤维素,提高其在水稻韧皮部的取食效率(Jietal., 2017);巢菜修尾蚜(Megouraviciae)利用口器刺穿蚕豆(Viciafaba)韧皮部时,会分泌唾液封闭植物筛管细胞所形成的伤口,导致Ca2+在筛管细胞内外无法产生浓度差,进而阻断Ca2+信号传递途径,抑制植物防御反应(Willetal., 2007)。在一定程度上,诱导抗性强弱与诱导因子类型相关,且在大多数情况下为正相关,诱导因子强度越高,诱导抗性表现越明显(张斌等, 2016)。因此,本研究舞毒蛾取食处理小黑杨体内的C4H和4CL基因表达水平高于机械损伤处理,可能是由于昆虫取食在一定程度上抑制了植物次生代谢产物的抗逆作用,而黄酮类、萜类和多糖类等相关产物的基因被诱导增强,可产生更多的次生代谢产物。机械损伤对小黑杨的伤害是瞬时且相对简单的,当产生的木质素达到一定程度时,相关基因表达量降低,这可能是苯丙氨酸代谢途径下游基因4CL表达量下调的原因。小黑杨C4H-1基因表达可能是取食和机械损伤作用下C4H活性升高的主要来源,而机械损伤导致4CL活性增加可能存在其他更多4CL家族基因表达导致的。
4 结论
舞毒蛾取食和机械损伤均能诱导小黑杨C4H和4CL活性增加,且取食诱导显著高于机械损伤; 舞毒蛾取食和机械损伤对C4H和4CL基因表达影响存在差异性。应进一步研究C4H和4CL家族基因功能及蛋白互作机制,以助于认识昆虫与寄主植物互作机制并通过遗传工程开发抗性植物控制害虫。
