青藏苔草形态特征及生物量分配沿水分梯度的变化
2021-04-09王洪斌张煜坤张大才
王洪斌, 张煜坤, 张大才
(国家林业和草原局西南地区生物多样性保育重点实验室, 西南林业大学林学院, 云南 昆明 650224)
湿地生态系统具有维持生物多样性、调节气候、涵养水源和保持区域生态平衡等多种功能,被称为“地球之肾”[1]。中国湿地类型多样、面积宽广,但大部分湿地面临过度放牧、污染和围垦等威胁[2-4],在气候干暖化及放牧等因素的长期影响下,沼泽湿地逐渐演替为沼泽化草甸、草甸、退化草甸甚至沙化草甸[5-6]。川西高原作为长江、黄河上游及其主要支流的源头,分布有大面积的沼泽湿地,生态战略地位十分重要[7-8],但湿地的退化十分严重,且退化过程由水分降低驱动[9-10]。因此,研究湿地植物对水分的响应对于沼泽湿地生态系统保护具有重要意义。
植物在适应异质生境时,功能器官的强弱变化与形态特征的分化能力常被称为表型可塑性,其强弱关系着植物对逆境的适应能力以及其在植物群落中重要值的大小[11-12]。植物可塑性越强,对生境的适应能力也越强[13]。在干旱化生境中,植物的生长高度、茎叶性状、克隆构件、根系生长及生物量都会发生相应的变化[14]。例如,华扁穗草(Blysmussinocompressus)地上、地下生物量都随土壤水分的减少而增加,干旱条件反而有利于其生物量的积累[15];高山嵩草(Kobresiapygmaea)在适应干旱化生境时,呈现出分株和根系数量及长度增加、叶片数量和茎基直径降低的形态变化,而矮生嵩草(K.humilis)会采取增加根系长度、减少分株数量和叶片数量同时减小茎基直径的适应策略[16]。此外,尽管植物器官形态特征之间存在显著的相关性,但对环境变化响应的敏感度存在差异[17]。有研究指出,旱生植物营养器官中,叶与植物生长密切相关,与外界环境进行物质能量交换最频繁,最容易发生定向进化,是可塑性最大的营养器官[18-19]。生境干旱化过程中,根系生长量增加可以增强水分的吸收,而叶片数量减小可以减小蒸腾作用[15-16],但湿地植物以怎样的机制响应生境干旱化以及其叶的可塑性是否也最强还尚待研究。
青藏苔草(Carexmoorcroftii)是川西高原高寒草甸和沼泽湿地的优势种,海拔分布范围为3 400~5 700 m[20],在水土保持、维持生态平衡方面起着重要作用[21-22]。然而,随着沼泽地水位的下降和高寒草甸生境干旱化的加剧,青藏苔草在植物群落中的重要值和生态位宽度逐渐下降[23]。沼泽与草甸的退化过程大多是土壤含水率降低的生境干旱化过程[24-25],目前,对于青藏苔草的相关研究主要在于遗传多样性[20]、生态位特征[21]、地理分布[26]及叶的形态特征随海拔梯度的变化[27]等方面,而少有研究涉及其根、茎、叶形态特征及生物量对土壤水分梯度的响应。本文以青藏苔草为研究对象,通过研究其根、茎、叶形态特征变化和生物量分配,以揭示青藏苔草营养器官形态特征沿土壤水分梯度的变化与营养器官形态特征可塑性的强弱排序,以及生境干旱化过程中植株个体生物量的分配策略。
1 材料与方法
1.1 研究地概况
研究地点位于四川省甘孜州新龙县银多乡古冰帽沼泽湿地,地理坐标为31°24′25″ N,99°49′48″ E,海拔4 300~4 450 m。据中国天气网(www.weather.com.cn)2009—2018年的资料,该区域属于青藏高原亚湿润气候区,年均降水量631 mm,年均气温7.9℃,年均最高气温18.3℃,年均最低气温1℃,全年60%以上降水量集中在6—8月。沼泽化草甸以青藏苔草为单优势种,高寒草甸以矮生嵩草、大花嵩草(K.macrantha)为共优种,伴生有青藏苔草、黑褐穗苔草(Carexatrofusca‘Minor’)、高山早熟禾(Poaalpina)、极地早熟禾(P.arctica)、丝颖针茅(Stipacapillacea)等。在放牧干扰下,高寒草甸退化严重,蓝玉簪龙胆(Gentianaveitchiorum)、大萼蓝钟花(Cyananthusmacrocalyx)、川西小黄菊(Tanacetumtatsienense)、星状风毛菊(Saussureastella)、高山唐松草(Thalictrumalpinum)、高山大戟(Euphorbiastracheyi)、委陵菜(Potentillachinensis‘Chinensis’)、珠芽蓼(Polygonumviviparum)、圆穗蓼(P.macrophyllum)等植物在干旱生境中常见,并在部分区域形成优势种。
1.2 样地设置与采样
样地位于两山之间的平缓地段,低凹区域为大面积沼泽水域,浅水区域分布有青藏苔草植物群落,高寒草甸分布于平缓坡面上(图1)。从水域开始,沿着距水域距离增加的方向,根据TSC-IW型土壤水分速测仪测量的土壤含水率,并结合青藏苔草的分布范围共设置5个4 m×4 m样地,其中2个位于有积水的沼泽化草甸,3个位于高寒草甸。青藏苔草在5个样地中均有分布,但各样地中个体的空间分布格局不尽相同:1号样地为青藏苔草沼泽化草甸,群落盖度90%以上,几乎没有其它植物分布,青藏苔草植株呈均匀分布;2~5号样地,青藏苔草植株空间分布格局逐渐从集群分布过渡到随机的单株分布,5号样地为青藏苔草分布的边缘,向样地外延伸几乎无青藏苔草植株分布(表1)。
为保证根系取样的完整性,采用挖掘法采样,对草地根系的研究表明,80%的根系分布于0~10 cm土层[28-29],因此挖取土块时,深度均为20 cm以上。样地中具有花、果的青藏苔草植株很少,部分样地甚至没有,处于花果期的植株茎会高出叶一些,因此选择茎比叶片矮(即花芽还未抽出)的植株做为研究对象。在1~2号样地,青藏苔草分布密集,盖度极高且生长状况均一,设置2个1 m×1 m样方挖取一定体积的土块,即可采集到30株以上青藏苔草;3~5号样地青藏苔草散生分布,盖度很低,各植株间距离较远且生长状况各不相同,设置3~6个1 m×1 m样方随机采集青藏苔草30株以上。将样品装入塑料袋,带回室内进行分株并测定地上、地下指标。采样时间为2018年8月。

图1 样地设置示意图Fig.1 Sketch map of sample plots setting
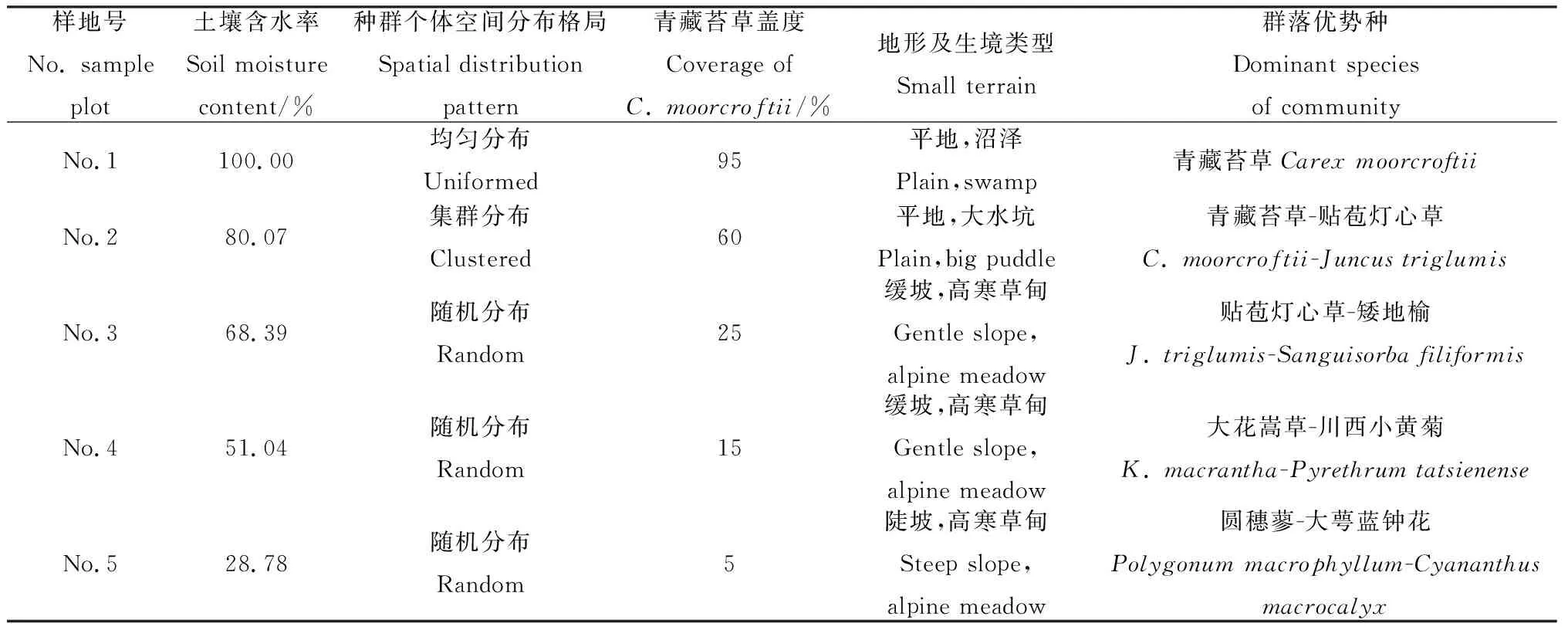
表1 样地信息Table 1 Information of sample plot
1.3 数据测量
1.3.1土壤含水率的测定 使用TSC-IW型土壤水分速测仪测量样地的土壤体积含水率,在干旱季节(9月底)测量,每个样地沿样地边界及对角线选取10个点测量土壤体积含水率,以平均值衡量每个样地的土壤含水率(表1)。
1.3.2植株形态特征的测量 将所有植株从土壤中完整地分离出来,每个样地选取长势相对均一的植株30株,分别统计叶片、残存叶基和分蘖芽数量,用游标卡尺测量植株茎基、根状茎和分蘖芽直径(mm),用精度为0.1 cm的直尺测量最长三片叶的长度(取平均值代表每株叶片的长度)和根状茎长度(cm)。完成茎、叶形态数据测量后,将根系分离出来,在尼龙网中将根系冲洗干净,采用TTC染色法区分出活根与死根,去除死根,将分离出的活根系铺开,使用Epson Perfection V700根系扫描仪进行扫描,并用配套的WinRHIZO根系分析软件获取各样地根系的总根长(cm)、表面积(cm2)、平均直径(cm)、体积(cm3)数据。
1.3.3生物量的测定 将每个样地中青藏苔草植株的地下、地上部分分装于不同的信封,在105℃下杀青30分钟后,于65℃下烘干48 h至恒重,用电子天平称量每个样地植株的地下(Belowground biomass,BB)、地上(Aboveground biomass,AB)生物量(g),并计算BB/AB值。
1.4 数据分析
获取的数据使用Excel软件记录并整理后,使用SPSS 25.0软件分析根、茎、叶形态特征及生物量与土壤含水率之间的关系。使用Origin 2019b软件进行绘图。
使用Canoco 4.5软件分析青藏苔草形态特征与土壤含水率之间的排序关系,根据去趋势对应分析(DCA)结果中“Length of gradient”判别排序模型的选择[30]。本研究中,“Length of gradient”最大值为0.26,选择RDA线性排序模型,将排序图第一轴,即横轴(—1,1)均等划分为3段,作为划分可塑性强弱等级的依据。第1段为(—0.333,0.333),可塑性弱;第2段为(—0.666,—0.333)&(0.333,0.666),可塑性中等;第3段为(—1,—0.666)&(0.666,1),可塑强。
2 结果与分析
2.1 青藏苔草形态特征与生物量对水分的响应
青藏苔草的分蘖芽直径、根系平均直径、总根体积与叶长、根系表面积、地下生物量、地上生物量分别在生境较湿润的1号和2号样地中达到最大值,残存叶基数量、总根长、根状茎长度与叶片数量、茎基直径、分蘖芽数量分别在生境较干旱的4号和5号样地中达到最大值,而根状茎直径则在3号样地中达到最大值(表2)。青藏苔草根、茎、叶形态特征在不同水分梯度中存在显著差异(P< 0.05,表2)。
2.2 形态特征与土壤含水率之间的关系
随着生境干旱化,青藏苔草根系表面积、根系平均直径、总根体积均呈减小趋势(图2A,B,C),而总根长呈增大趋势(图2D)。与最湿润生境样地(No. 1)相比,最干旱生境样地(No. 5)总根长增加了55.62%,根系表面积、根系平均直径和总根体积分别减小了25.49%,54.71%和63.37%。在生境干旱条件下,根系变细变长有利于青藏苔草吸收、利用深层土壤水分。
青藏苔草叶长随土壤含水率降低而减小(图3A),叶片数量、残存叶基数量、茎基直径均随土壤含水率降低而增大(图3B,C,D)。叶片缩短、数量增多是青藏苔草在干旱化生境中维持光合产出的适应策略。
青藏苔草分蘖芽数量、根状茎长度随土壤含水率降低呈增大趋势(图4A,B),分蘖芽和根状茎直径均随土壤含水率降低而减小(图4C,D)。克隆构件营养物质的分配存在权衡关系,随着土壤含水率降低,根状茎形态趋向更细、更长,分蘖芽趋向数量更多而形态更小,朝着占据更多生存空间的方向发展。

表2 青藏苔草形态特征及生物量Table 2 Morphological characteristics and biomass of Carex moorcroftii

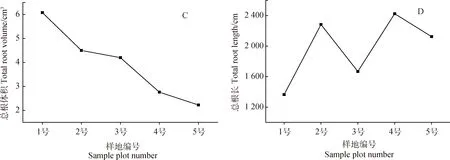
图2 根系形态特征的变化Fig.2 Variation of root morphological characteristics
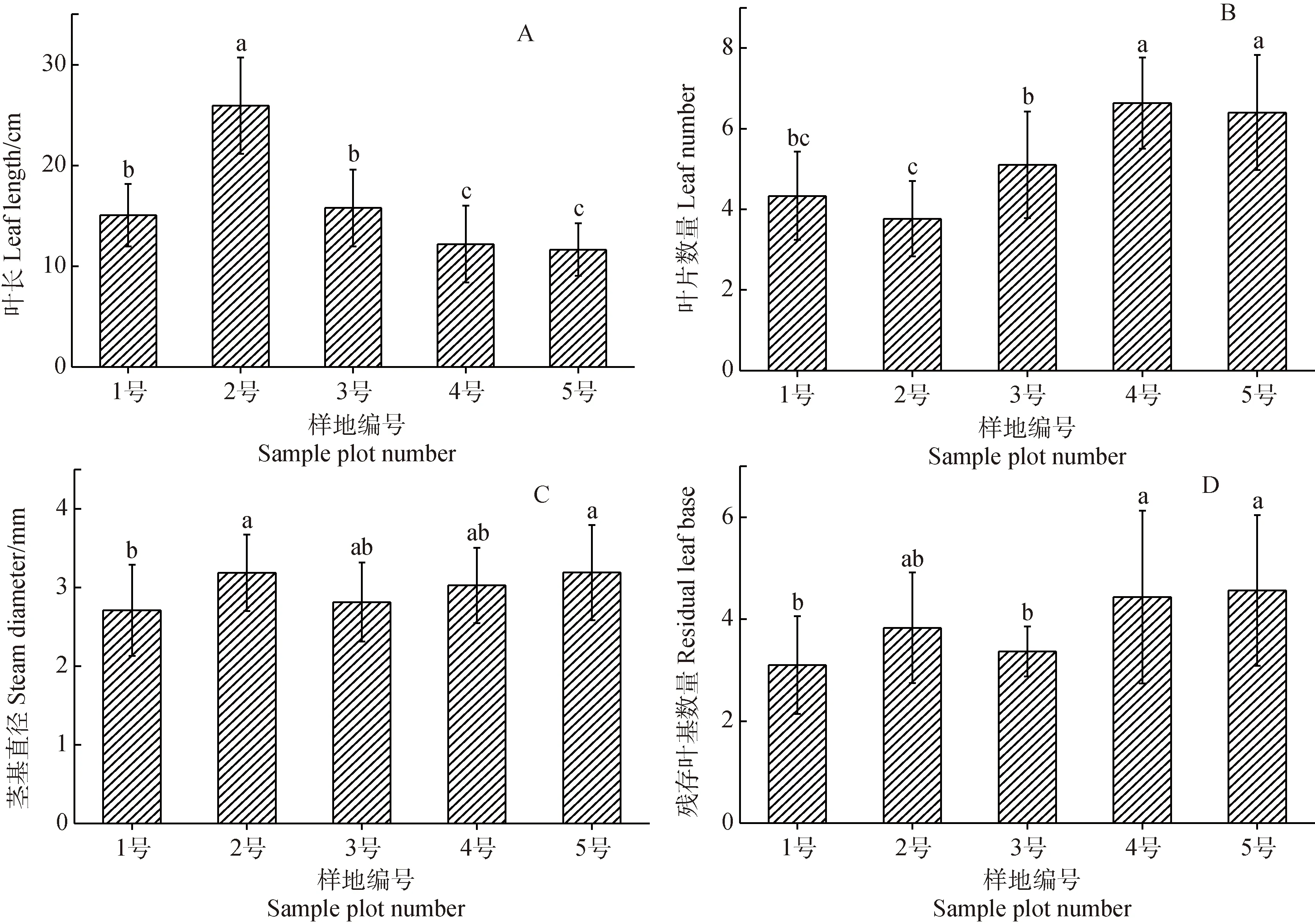
图3 茎、叶形态特征的变化Fig.3 Variation of stem and leaf morphological characteristics
2.3 水分梯度下生物量的变化与分配
青藏苔草地下、地上生物量均随土壤含水率的降低而减小,BB/AB值先减小后增大(图5)。2号样地地下、地上生物量最大,分别为4.8 g,18.21 g;5号样地地下、地上生物量最小,分别为3.36 g,13.30 g;4号样地的BB/AB值最小,为0.23,1号样地的BB/AB值最大,为0.27。
2.4 营养器官形态可塑性排序
根据RDA排序图,青藏苔草形态特征可塑性对土壤水分梯度的响应可以分为以下3类:(1)可塑性强形态特征,包括总根体积、根系平均直径、根状茎长度;(2)可塑性中等形态特征,包括叶长、分蘖芽直径、根状茎直径、根系表面积、叶片数量、残存叶基数量、总根长;(3)可塑性弱形态特征,包括地上生物量、地下生物量、茎基直径、分蘖芽数量。各个形态特征的可塑性强弱不同,因此随土壤含水率变化的程度也不同,存在非对称响应机制。综合营养器官形态特征可塑性可得出,根的可塑性最强,茎的可塑性中等,叶的可塑性最弱。

图4 克隆构件形态特征的变化Fig.4 Variation of morphological characteristics in clonal modular
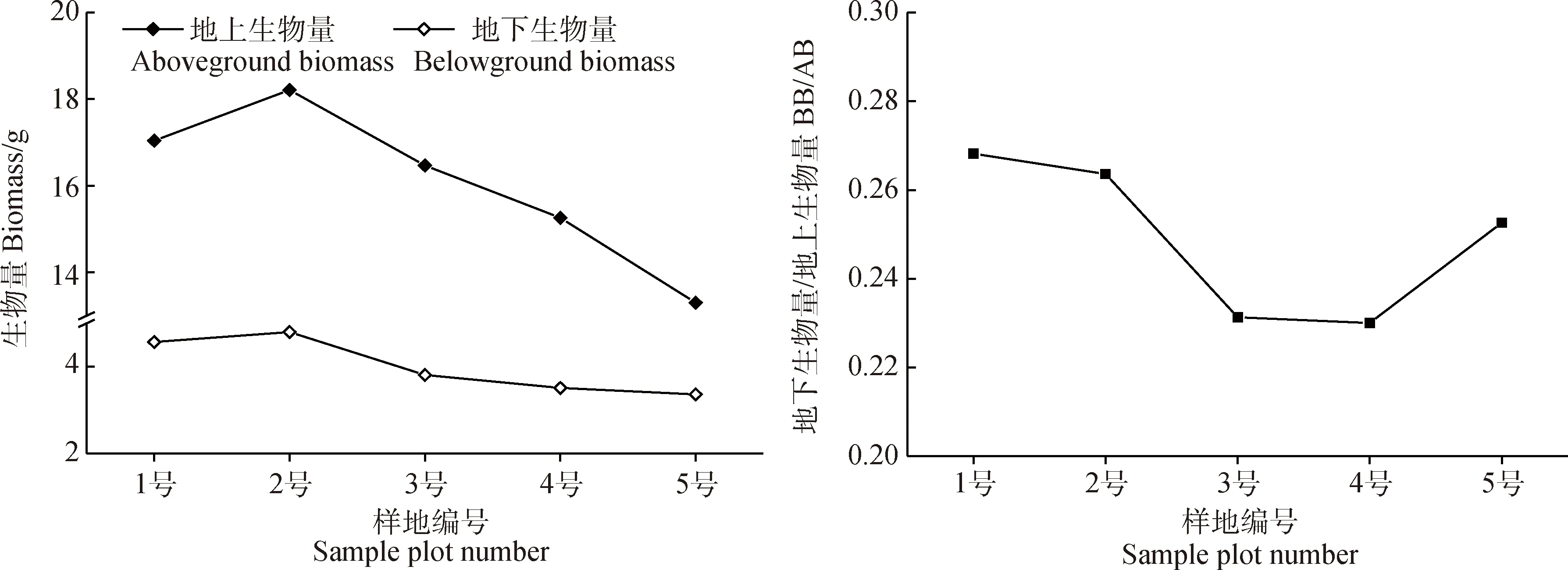
图5 生物量与BB/AB值的变化Fig.5 Variation of biomass and BB/AB ratio
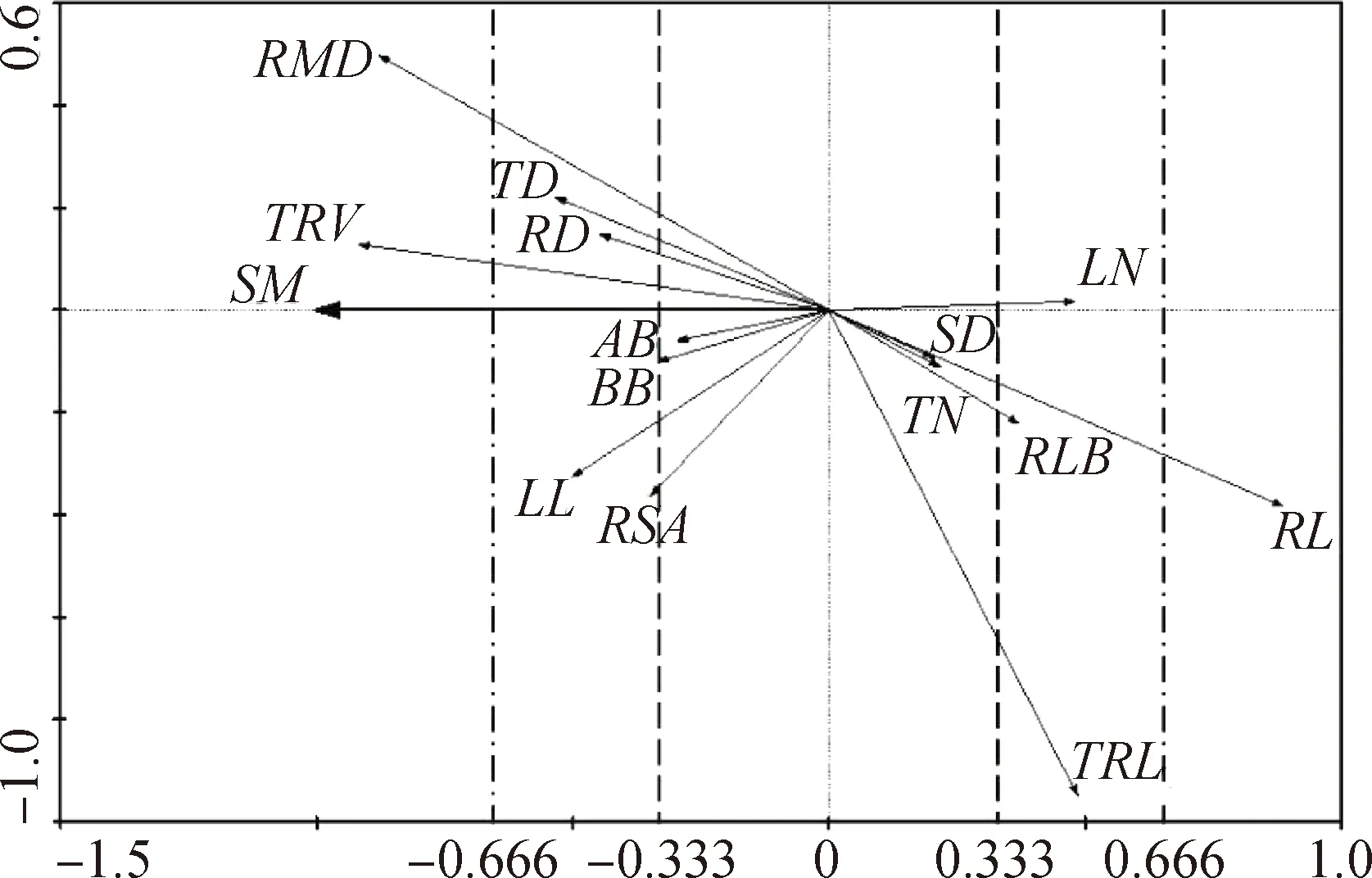
图6 形态特征与土壤含水率RDA排序图Fig.6 RDA ordination diagram of morphological characters and soil moisture content注:SM,土壤含水率Note:SM,soil moisture
3 讨论
3.1 形态特征的非对称响应
植物在受到干旱胁迫时,将调整形态特征之间的关系以适应生境变化,这将导致形态特征对水分响应敏感度的分化[31],即在生境梯度下,植物不同性状在变化的同时,变化的程度却存在较大差异,这种非对称响应机制(Asymmetric responses)在生态系统中普遍存在,已有学者进行了相关报导[32]。RDA排序分析将青藏苔草形态特征对水分响应的敏感度分为强、中、弱3类(图5),表明青藏苔草形态特征沿土壤水分梯度的变化存在非对称响应机制。随着生境干旱化,青藏苔草总根体积、根系平均直径可塑性大,地上生物量、叶片数量可塑性小,这与黄栌[33](Cotinuscoggygria)的研究结果一致,说明土壤水分的降低大大刺激了植株的根系发育,根在青藏苔草适应干旱生境的过程中起着十分重要的作用。随着生境干旱化,生物量分配朝着根茎分配、叶片分配减小的方向发展,这与根、茎、叶形态特征的非对称响应有关,根的直径、表面积、体积减小的程度大于根长增加的程度,分蘖芽、根状茎直径减小的程度大于分蘖芽数量、根状茎长度增加的程度,故其地下生物量下降;叶长、茎基直径减小的程度大于叶片数量增加的程度,故其地上生物量下降。青藏苔草对各种性状进行权衡,根、根状茎与分蘖芽偏向细长化,叶偏向数量化,以达到在干旱化生境中的最优生活对策。
3.2 营养器官形态特征对水分梯度响应的差异
根是植物吸收水分和养分的主要营养器官,当水分成为限制植物生长的条件时,植物会增加根长,使根系吸收利用深层土壤水分,从而适应干旱生境[34-35]。对青藏苔草的研究发现,生境干旱化使得青藏苔草根系形态特征发生了改变,且各个指标的变化趋势存在差异。根系平均直径、根系表面积、总根体积均随生境干旱化程度的增加呈减小趋势,而总根长随生境干旱化程度的增加呈增大趋势(图2)。高山嵩草和矮生嵩草在适应干旱化生境时,同样呈现出根系长度增加的形态变化[16],这说明青藏苔草和高山嵩草、矮生嵩草在干旱条件下的生存策略相似,都会通过调节根直径、表面积、体积与根长的关系来利用极为有限的资源。在干旱生境中,青藏苔草得不到充足的水资源,光合作用被抑制,仅能获得有限的光合产物,故其根系直径减小,将有限的资源用于根系的伸长生长,以利用深层土壤水源。
随着生境干旱化,青藏苔草呈现出叶长缩短,茎基直径增大,叶片与残存叶基数量增多的形态变化特征(图3)。在生境干旱化条件下,青藏苔草叶长缩短的形态变化与西藏嵩草(K.tibetica)的叶片变化相同[36];茎基直径的形态变化与其它高原湿地植物受到干旱胁迫后的变化相反[37];叶片数量的变化与灰化苔草(C.cinerascens)、高山嵩草和矮生嵩草在生境干旱化过程中通过减少叶片数量降低蒸腾作用,减少水分散失的变化不同[16,38],说明青藏苔草与其他植物在干旱条件下的生存策略有所不同。叶片是植物形成光合产物的重要器官,叶长缩短虽然极大的降低了青藏苔草与其他植物的竞争力[39],但水分与养分的运输距离缩短,大大提升了运输效率[40],并节省了运输的能量消耗。因此,在叶长缩短的情况下,增加叶片数量对维持植株的光合叶面积具有重要意义,这是青藏苔草在干旱胁迫生境中的最优适应策略。茎基部是多年生草本植物叶片的着生区,有着贮藏营养物质的作用,是叶片数量增加的基础,茎基直径越大,贮藏的营养物质越多,能供给叶片生长的营养就越多[16],残存叶基数量则随叶片数量的增加而增多。综上,在生境干旱化的影响下,青藏苔草缩短叶长提升运输效率,同时增加茎基直径及叶片数量以维持光合作用,残存叶基数量也随叶片数量的增多而增多,4个形态特征之间表现出很强的相关性。
克隆植物的生殖主要靠克隆构件形成新的无性系分株来完成,无性繁殖力强,无性系存活的时间也较长[41-42]。随着生境干旱化,青藏苔草的根状茎直径减小、长度增加,分蘖芽直径减小、数量增多(图4)。分蘖芽数量的变化与生境干旱化条件下高山嵩草的响应机制相同[16],说明青藏苔草与高山嵩草在干旱化生境中的无性繁殖能力都增强。随着干旱化程度增加,青藏苔草的资源积累受到限制,导致其资源进行转移,具体表现为根状茎与分蘖芽的直径减小,而根状茎长度与分蘖芽数量增加,这种资源的分配转移有利于青藏苔草占据土壤空间,增加与其他植物的竞争力。已有研究表明,苔草属植物存在生理整合现象(Physiological Integration),即在由地下根茎相连的各分株之间,营养物质和水分可以通过根茎进行运输和转移,实现整个无性系的资源共享,从而提高种群对胁迫环境的适应性[43]。青藏苔草作为典型的克隆植物,在土壤水分亏缺的情况下,既存在一般植物常有的适应生境干旱化的机制,又发挥了克隆植物的优势,占据更多生存空间,以提高对干旱环境的适应能力。
3.3 生物量分配策略
随着生境干旱化,青藏苔草的地下、地上生物量都减小,这与西藏嵩草在土壤水分减少条件下地下、地上生物量也减小的研究结果一致,而与矮生嵩草、鹅绒委陵菜(Potentillaanserina)、海乳草(Glauxmaritima)和华扁穗草地下、地上生物量与土壤含水率之间的关系相反[15,36]。矮生嵩草、鹅绒委陵菜、海乳草、和华扁穗草适宜生长于土壤水分适中的平缓滩地或山地阳坡,是耐旱性较强的植物。而西藏嵩草喜欢分布在潮湿或积水的环境中,这与青藏苔草的生境喜好相同,因此,当生境干旱化程度加剧,青藏苔草的地下、地上生物量分配与喜湿植物相同,与矮生嵩草等植物存在差异。青藏苔草BB/AB值随着生境干旱化,总体上呈现出缓慢降低的趋势,在最湿润生境(0.27)中BB/AB值最大,在半干旱半湿润生境(0.23)中BB/AB值最小,而在最干旱生境(0.25)中出现回升。其原因是随着生境干旱化,地上生物量减小的程度比地下生物量减小的程度要大的多。即青藏苔草在干旱化生境中更偏向于维持地下生物量,减小地上生物量。
生物量可以直观地反映研究对象积累有机物的多少,关系着植物的发育及其结构组成,植物会通过改变生物量的分配以适应逆境[44]。当环境发生改变时,植物将改变对不同器官的生物量分配,最大限度地提高获取各种资源的能力[45],随着生境干旱化,青藏苔草的地下、地上生物量都减小,说明其在干旱化的生境中获取维持生命和生长所需资源的能力有限,故随着土壤含水率降低,它的个体空间分布格局从均匀分布逐渐过渡到随机分布,个体数量及分布范围逐渐减小。此外,青藏苔草的地下生物量减小的程度小于地上生物量,说明在干旱化生境中,地下部分对其适应干旱生境的意义更大。
4 结论
为适应干旱化生境青藏苔草会改变自身表型特征以维持自身正常生长和繁殖,根的可塑性比茎和叶强,地下部分对其适应干旱化生境的意义更大。青藏苔草适应干旱化生境的策略为:叶片长度缩短、数量增多在缩短输导距离的同时保持植株的叶面积;发挥克隆植物的优势,增加分蘖芽数量与根状茎长度以增强对空间的占据能力;减小根系直径,增加根系长度以吸收利用深层土壤水分。根、茎、叶形态特征具有一定的可塑性,但生物量的降低减弱了青藏苔草与其他植物的竞争力,限制了青藏苔草在干旱生境中的分布。研究结果有助于深入认识干旱区湿地植物对土壤水分的响应关系,对于合理利用干旱区湿地资源、保护干旱区湿地生态系统具有重要参考价值。
