成熟过程中细齿草木樨种子结构及产量和质量的变化
2021-04-09陈立强白小明
陈立强,白小明
(1. 甘肃农业大学草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中美草地畜牧业可持续发展研究中心,甘肃 兰州 730722;2. 塔里木大学动物科学学院/新疆生产建设兵团塔里木畜牧科技重点实验室, 新疆 阿拉尔 843300)
细齿草木樨(MelilotusdentatusPers.)为豆科草木樨属一年生草本植物,产量高、品质好,具有抗寒、抗旱、耐盐碱、耐瘠薄、抗风沙等特性,是优良的牧草、绿肥和水土保持植物。细齿草木樨香豆素含量是黄花草木樨(MelilotusofficinalisPall.)的1/28~1/122,是白花草木樨(MelilotusalbusDesr.)的1/35~1/140,由于其香豆素含量较低而被誉为“无味草木樨”。细齿草木樨适口性较好,具有较高的饲用价值,在我国主要分布在东北的西部、内蒙古的中东部和黄河流域的宁夏、陕西、山西、河南和河北上东等地[1-3]。
种子发芽力是指种子成为正常种苗的能力,决定着种子能否用作播种材料,受内部因素和外部环境的双重影响,其中胚发育不完全和生理未成熟,种皮的硬实和胚乳的阻碍等内部因素都会遏制种子萌发[4-5]。为了解决这一难题,大量研究从种子结构方面对萌发限制因素进行了探索。祝明珠等[6]研究了多花黄精(PolygonatumcyrtonemaHua.)种子结构和发芽率的关系,发现栅栏细胞层、胚乳细胞和脂类物质可能都是种子萌发的限制因素;宋芸等[7]研究表明苦参(SophoraflavescensAlt.)种子的角质层和骨状石细胞层可能与种子休眠有关;桂喆和孙海群[8]对白花草木樨、黄花草木樨和其它4种豆科植物种子结构和硬实的关系进行了研究,发现栅栏细胞层和骨状石细胞层可能是影响豆科植物种子硬实的主要因素。从种子结构判断,不同物种影响其种子萌发的因素也各不相同。目前关于草木樨属种子结构的研究以黄花草木樨和白花草木樨为主[8],缺乏对细齿草木樨的研究,且相关研究报道只局限于某一阶段种子的结构分析[8],缺乏成熟过程中种子结构动态变化规律的研究。
由于不同成熟阶段种子的产量和质量都不尽相同[9-11],不恰当的收获时间会导致种子产量减少和品质下降。收获时间过早,种子的生活力较弱、产量较低、质量较差;收获时间过晚,落粒性增强[12]。前人研究表明,白花草木樨种子适宜的收获时间是盛花后33~42 d[9],但由于同属不同种或品种的花期存在差异[13-14],尚不清楚其它草木樨的种子收获时间是否与白花草木樨相吻合。目前我国草木樨种子严重缺乏,明确细齿草木樨及同属其它牧草种子适宜的收获时间,有助于提高种子的产量和品质,对其推广种植具有重要的意义。
为了使试验结果可靠实用,贴近生产,本研究以生产性能优异的细齿草木樨为材料,参考普通高等教育“十一五”国家级规划教材《牧草种子学》[15]定义的4个种子成熟阶段,依据大田种子的形态特征进行取样,通过分析成熟过程中种子细胞结构、产量和质量的变化规律,探索影响细齿草木樨种子发芽力的主要因素,为促进种子萌发,推动草木樨属牧草种子生产提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于新疆阿拉尔市科克库勒镇(81°19′ E,40°35′ N)。属暖温带极端大陆性干旱荒漠气候,干旱少雨,蒸发强烈。该地年降雨量40.1~82.4 mm,年蒸发量≥2 100 mm,年日照时数2 556~2 991 h,无霜期200 d以上,平均气温7.5~11.7℃,>10℃年积温3 800~4 700℃[16-17]。试验地前茬作物为甜高粱。
1.2 试验材料及设计
试验材料为细齿草木樨新品系,属一年生植物,原始群体为野生种质资源,分布于新疆阿拉尔市。2019年4月20日播种,小区面积30.6 m2(5.1 m×6.0 m),4个重复,条播,行距30 cm。理论播种量15 kg·hm-2(因土壤盐碱,播种量提高),播种深度1.3~1.5 cm。出苗后第10 d、第20 d、分枝期和初花期各漫灌一次。苗期人工除杂草2次。取样时间分别为绿熟期、绿熟期+5 d、绿熟期+10 d、绿熟期+15 d、黄熟前期、黄熟前期+5 d、黄熟前期+10 d、黄熟前期+15 d、黄熟前期+20 d、黄熟后期、黄熟后期+5 d和完熟期,样品编号分别为1~12号。取样时避免边行及两头50 cm之内的植株,每小区按行从前到后的顺序割取30株样品,用于测定试验指标。
1.3 测定指标及方法
1.3.1种子结构 分别选取绿熟期、黄熟前期、黄熟后期和完熟期[15]的代表性种子各3粒,用解剖针刺破种皮,置于蒸馏水中,于4℃冰箱中浸泡12 h,充分吸胀后,用10%甲醛溶液固定48 h,水洗48 h后,用酒精脱水,二甲苯透明。用YD-6D智能型生物组织包埋机进行包埋,用LEICARM2235切片机切片(纵切),厚度为10 μm,贴片和烤片后用伊红-苏木精染色[18]。在ECLIPSEE200型显微镜下观察,并采集图像。每张切片选取5个视角,用ImageJ软件测定角质层厚度、栅栏细胞层厚度、骨状石细胞层厚度和糊粉层厚度,取平均值。对种子成熟过程中子叶细胞、胚根细胞和维管组织的变化规律进行观察和描述。
1.3.2种子的产量和质量 脱粒和清选后称取种子产量。随机选取1 000粒种子,测定千粒重。将同一成熟阶段不同重复的种子充分混匀,测定生活力、发芽势、发芽率和硬实率。种子的生活力参照《草种子检验规程生活力的生物化学(四唑)测定(GB/T 2930.5-2017)》[19]进行测定,随机数取400粒种子,每重复100粒,刺破种皮后,浸入1%的四唑溶液,30℃黑暗条件下浸泡18 h后取出,统计生活力百分率。参照《草种子检验规程发芽实验(GB/T 2930.4-2017)》[20]测定发芽势、发芽率和硬实率,随机数取400粒种子,每重复100粒,整齐摆放在直径为12 cm,铺有1张滤纸的培养皿中,置于温度为20℃,光照强度为4 400 lx,光暗周期为12 h/12 h的光照培养箱中进行培养,期间保持滤纸湿润,第4天统计发芽势,第7天统计发芽率和硬实率。用Seedburo电动种子划痕机(实验用)对种子进行划痕处理后,按上述方法再次测定发芽势、发芽率和硬实率。
1.4 数据处理
用Excel 2007进行数据整理。分别用SPSS 19.0中的Duncan氏新复极差法和Pearson相关性分析法对不同收获期细齿草木樨种子的种皮、产量和质量性状进行差异显著性分析和相关性分析。
2 结果与分析
2.1 成熟过程中细齿草木樨种子结构的变化规律
2.1.1成熟过程中细齿草木樨种皮结构的变化规律 由图1可知,尽管不同成熟阶段细齿草木樨的种皮结构不尽相同,但总体上可分为角质层、栅栏细胞层、骨状石细胞层和糊粉层。角质层位于种皮的最外层。栅栏细胞冠与角质层紧密相连,呈莲花状排列,接着是明线和栅栏细胞腔,栅栏细胞呈篱笆状,与表皮细胞垂直,分布均匀,排列紧密。骨状石细胞层与栅栏细胞层紧密相连,有较大的细胞间隙。随着种子成熟,骨状石细胞的数量明显增多,与骨状石细胞层相连的是糊粉层。由表1可知,不同成熟阶段角质层厚度介于5.23~6.86 μm之间,其在绿熟期最大,黄熟前期次之,然后是黄熟后期和完熟期,随着种子成熟呈降低的变化趋势,但不同成熟阶段角质层厚度差异不显著。栅栏细胞层厚度介于50.42~54.98 μm之间,随着种子成熟呈增加的变化趋势,其在完熟期最大,极显著高于绿熟期(P<0.01),显著高于黄熟前期和黄熟后期(P<0.05)。骨状石细胞层厚度介于17.96~26.01 μm之间,其在完熟期最大,黄熟后期次之,然后是黄熟前期和绿熟期,随着种子成熟呈上升的变化趋势,但不同成熟阶段骨状石细胞层厚度差异不显著。糊粉层厚度介于10.81~17.68 μm之间,其在完熟期最大,绿熟期最小,显著低于其它成熟阶段(P<0.05)。
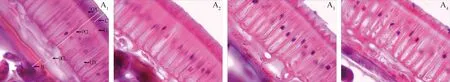
图1 不同成熟阶段细齿草木樨的种皮结构Fig.1 Seed coat structures of Melilotus dentatus in different mature stages注:A1-A4分别为绿熟期、黄熟前期、黄熟后期和完熟期的种皮结构。C-角质层,PCL-栅栏细胞层,OCL-骨状石细胞层,AL-糊粉层,CPC-栅栏细胞冠,LL-明线,LPC-栅栏细胞腔Note:A1-A4 were seed coat structures in green,early yellow,late yellow and full ripe stages,respectively. C-Cuticle,PCL-Palisade cell layer,OCL-Osteosclerid cell layer,AL-Aleurone layer,CPC-Cap of palisade cell,LL-Light line,LPC-Lumen of palisade cell
2.1.2成熟过程中细齿草木樨种子子叶细胞和胚根细胞结构的变化规律 细齿草木樨为双子叶植物,早在绿熟期,子叶中已积累大量的贮藏物质(图2-B1)。从黄熟前期到黄熟后期和完熟期,子叶中细胞明显变大(图2-B2,B3,B4)。绿熟期胚根细胞的形态不规则,呈多边形或圆形,部分细胞发生分裂,形成新的细胞(图2-C1)。黄熟前期发生分裂的细胞数量大幅增加,分裂后的细胞形态变为条形(图2-C2)。完熟期胚根中大部分细胞发生分裂,分裂后细胞呈玉米粒状排列(图2-C4)。
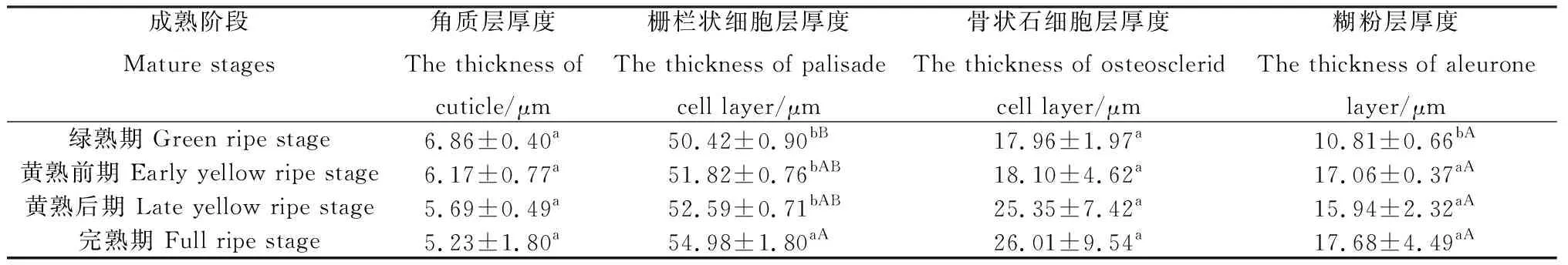
表1 不同成熟阶段细齿草木樨种皮的径向切面参数Table 1 Radial section parameters of seed coat of Melilotus dentatus in different mature stages

图2 不同成熟阶段细齿草木樨种子子叶和胚根中的细胞和维管束Fig.2 Cells and vascular bundles in seed cotyledon and radicle root of Melilotus dentatus in different mature stages注:B1-B4分别为绿熟期、黄熟前期、黄熟后期和完熟期种子子叶中的细胞和维管束,图中VB表示维管束;C1-C4和D1-D4分别为上述4个成熟阶段胚根中的细胞和维管束Note:B1-B4 were cells and vascular bundles of seed cotyledon in green,early yellow,late yellow and full ripe stages,respectively,on which vascular bundles were marked with VB;C1-C4and D1-D4were cells and vascular bundles of seed radicle root in above four mature stages,respectively
早在绿熟期子叶和胚根中均已分化出维管组织,形成维管系统的雏形,但维管组织长度较短,时断时续,间隙较大(图2-B1,图2-D1)。黄熟后期维管组织变长,间隙变小,至完熟期维管组织再次伸长,相互连接构成了完整和连贯的维管系统(图2-B3,B4;图2-D3,D4)。
2.2 成熟过程中细齿草木樨种子产量和千粒重的变化规律
供试样品的编号及其产量和质量指标如表2所示。随着种子成熟,细齿草木樨的产量和千粒重呈上升的变化趋势,种子产量介于12.32~73.85 g·(30株)-1之间,完熟期12号样品的产量最高,显著高于黄熟后期前取的1~9号样品(P<0.05),极显著高于1~8号样品(P<0.01)。成熟早期1~5号样品的种子产量介于12.32~19.26 g·(30株)-1之间,彼此间差异不显著,但显著低于黄熟前期后取的6~12号样品(P<0.05)。不同成熟阶段样品的千粒重介于1.35~2.96 g之间,完熟期12号样品的千粒重最高,极显著高于1~10号样品(P<0.01)。而1号和3号样品的千粒重较低,显著低于其它样品(P<0.05)。

表2 不同成熟阶段细齿草木樨种子的产量和质量指标Table 2 Seed yield and quality indexes of Melilotus dentatus in different mature stages
2.3 成熟过程中细齿草木樨种子生活力、发芽力和硬实率的变化规律
随着成熟,细齿草木樨种子生活力和硬实率呈上升的变化趋势,发芽势和发芽率呈先升后降的变化趋势。供试样品的生活力百分率介于48.75%~100.00%之间,完熟期12号样品的生活力百分率最高,极显著高于1~10号样品(P<0.01),显著高于11号样品(P<0.05)。而成熟早期1号和2号样品的生活力百分率最低,极显著低于3~12号样品(P<0.01)。供试样品的发芽势介于2.25%~17.00%之间,发芽率介于3.25%~19.75%之间,其中黄熟前期后5 d取的6号样品发芽势和发芽率最高,极显著高于其它供试样品(P<0.01)。而成熟早期1号、2号、3号和成熟晚期11号和12号样品的发芽势和发芽率较低。供试样品的硬实率介于38.25%~87.25%之间,成熟晚期12号和11号样品的硬实率分别为87.25%和85.25%,极显著高于1~10号样品(P<0.01)。成熟早期1~4号样品的硬实率较低,分别为38.25%,41.50%,41.50%和43.00%,极显著低于其它供试样品(P<0.01)。
2.4 划痕后细齿草木樨种子发芽力和硬实率的变化规律
划痕处理后,1~12号样品的发芽势介于28.00%~96.00%之间,发芽率介于33.00%~96.75%之间。完熟期12号样品的发芽势和发芽率仍然最高,极显著高于1~11号样品(P<0.01)。而绿熟期1号样品的发芽势和发芽率最低,极显著低于2~12号样品(P<0.01)。
2.5 相关性分析
由表3可知,细齿草木樨种子的硬实率与骨状石细胞层厚度、种子产量、千粒重、生活力百分率呈显著正相关关系,与栅栏状细胞层厚度、糊粉层厚度、发芽势和发芽率呈正相关关系,与角质层厚度呈显著负相关关系。种子的生活力百分率与发芽势和发芽率呈正相关关系,但相关系数较小。

表3 细齿草木樨种皮、产量和质量性状的相关性Table 3 Correlation of seed coat,yield and quality traits of Melilotus dentatus
3 讨论
3.1 细齿草木樨发芽力的影响因素分析
本研究发现在自然状态下细齿草木樨种子的发芽率较低,但经划痕处理后,发芽率大幅提高,这种情况与黄花草木樨[8]、白花草木樨[9]、苦豆子(SophoraalopecuroidesLinn.)、苦马豆(Sphaerophysasalsula(Pall.) DC.)、胀果甘草(GlycyrrhizainflataBatal.)[21]和草木樨状黄芪(AstragalusmelilotoidesPall.)[22]等豆科植物类似。因此,细齿草木樨的种皮硬实是影响种子发芽的主要因素。种皮是种子最外的保护层,由胚珠和珠被发育而成,不含原生质,均为无生命的死细胞[15]。崔芬芬等[23]认为种皮的角质层和栅栏细胞层可能是影响种皮硬实的主要因素。Bevilacqua等[24]和胡小文等[25]认为栅栏细胞层主要影响种皮硬实,而桂喆和孙海群[8]及Graaff等[26]认为除栅栏细胞层外,骨状石细胞层也是影响种皮硬实的主要因素。本研究结果表明,从绿熟期到完熟期,种子栅栏细胞层厚度和骨状石细胞层厚度呈持续上升的变化趋势,相关性分析结果表明种子的硬实率与骨状石细胞层厚度呈显著正相关,与栅栏状细胞层厚度呈正相关。据此判断,栅栏细胞层和骨状石细胞层可能都是影响细齿草木樨种皮硬实的主要因素。
3.2 成熟过程中细齿草木樨种子产量和质量的变化规律分析
本研究结果表明,随着种子成熟,细齿草木樨种子的千粒重呈上升的变化趋势,这与耿本仁和王金玲[27]研究白花草木樨的结果一致,表明细齿草木樨和白花草木樨种子的干物质在成熟过程中不断积累。细齿草木樨种子的产量也呈上升的变化趋势,完熟期达到最高,显著高于其它成熟阶段(P<0.05),而李鸿祥等[9]的研究结果表明,白花草木樨的种子产量随着成熟呈先升后降的变化趋势,这种差异可能是由于与白花草木樨种质资源相比,细齿草木樨落粒性相对较小。种子的生活力、发芽率和硬实率是影响种子质量的重要因素。通常生活力越高,种子适应不良环境的能力越强,生活力低,则种子出苗不整齐,甚至不能出苗[28-30]。本研究结果显示,随着成熟,细齿草木樨种子结构逐渐完善,生活力呈上升的变化趋势,这与高羊茅(FestucaelataKeng ex E. Alexeev.)[31]、垂穗披碱草(ElymusnutansGriseb.)[32]和春小麦(TriticumaestivumL.)[33]种子成熟过程中生活力的变化趋势相同,但由于种子趋于硬实,发芽势和发芽率并未随生活力的增加而持续上升。本研究中细齿草木樨种子的硬实率极高,可能与其原始群体的野生性状有关,这与桂喆和孙海群[8]研究野生白花草木樨、黄花草木樨、广布野豌豆(ViciacraccaL.)、箭筈豌豆(ViciasativaL.)、高山豆(Tibetiahimalaica(Baker) Tsui.)、紫花苜蓿(MedicagosativaL.)时所得结论一致。
3.3 细齿草木樨适宜的收获时间及解除硬实的必要性分析
由于细齿草木樨种子的硬实率过高,导致不同成熟阶段种子的发芽势和发芽率普遍较低,最大值仅为17.00%和19.75%,用作播种材料时,必须先破除种皮硬实。经划痕处理后,种子的产量、千粒重、发芽势和发芽率的最大值均出现在完熟期。因此,完熟期为细齿草木樨种子最适宜的收获时间。大量研究表明,砂纸打磨和硫酸浸泡可大幅提高豆科植物的发芽率[34-36],但前者过于耗时费力,后者所需硫酸数量过多,加之购买困难,危险性过高,用于生产尚有缺陷。由于一年生牧草每年需要重新播种,其流通方式与多年生牧草不同,农户通常采用自产留种的方式繁殖种子。因此,简易的硬实破除方法尚待进一步研究。
4 结论
种皮硬实严重抑制了细齿草木樨种子发芽,栅栏细胞层和骨状石细胞层可能是影响细齿草木樨种皮硬实的主要因素。随着种子成熟,细齿草木樨的产量、千粒重和硬实率呈上升的变化趋势,发芽势和发芽率呈先升后降的变化趋势。细齿草木樨在新疆阿拉尔市最适宜的收获时间为完熟期,用作播种材料时,必须要先破除硬实。
