达乌里胡枝子花粉萌发特性研究
2021-04-09佟莉蓉倪顺刚
佟莉蓉, 王 娟, 宋 雨, 倪顺刚
(山西农业大学草业学院, 山西 太谷 030801)
达乌里胡枝子(Lespedezadaurica)为豆科胡枝子属的多年生草本状半灌木植物;其花序腋生,花冠蝶形,花为白色或黄白色[1],花期为7—8月;具有耐寒冷和耐贫瘠等优良特性[2],在我国分布广泛;具有很高的营养和药用价值以及生态效应[3]。
花粉的质量和生活力是影响受精成功的重要因素,花粉离体培养能为花粉提供与柱头相似的萌发条件[4]。花粉离体培养分为液体培养和固体培养基,研究表明使用液体培养基和固体培养基的效果差异不显著,而液体培养基易观察、操作简便,相比于固体培养基更能反映花粉的发育状况和实际活力[5-6]。其中花粉萌发率和花粉管长度能反映花粉生活力的高低[5]。在离体培养过程中,培养基中的营养成分如蔗糖、硼酸和钙的质量分数以及外界条件如培养时间和贮藏温度等均可以影响花粉离体萌发生长,不同植物不同品种不尽相同[7-8]。
近年来,国内外对花粉萌发和花粉管生长的研究主要集中在农业作物和经济作物上。通过对二倍体马铃薯(Solanumtuberosum)[9]、海仙花(Weigelacoraeensis)[10]、桃花(Prunuspersica)[11]和云南油杉(Keteleeriaevelyniana)[12]的花粉进行离体萌发培养,确定了最适培养基的组分以及培养时间。目前对胡枝子属的研究主要集中在开花习性、种子发芽特性、遗传多样性、药用成分等方面[13],有关达乌里胡枝子花粉离体萌发条件的研究普遍较少。本试验对影响达乌里胡枝子花粉生活力的因素进行测定,筛选出达乌里胡枝子花粉萌发的培养配方及最适培养条件,以期为其杂交育种和种质资源保存提供参考。
1 材料方法
1.1 试验材料
试验所用‘晋农一号’达乌里胡枝子(Lespedezadaurica‘Jinnong No.1’,缩写为LD)为山西农业大学草业学院自主选育品种,花粉于2019年8月采自草业学院试验田(112°34′ E,37°21′ N,海拔800 m)。
1.2 试验方法
1.2.1花粉采集与离体培养 选取生长健壮,长势良好的植株,于盛花期(8—9月)选取含苞待放的花朵带回实验室,用解剖针和镊子将花粉抖落到硫酸纸上包好放入硅胶中阴干备用。本试验所有处理所用花粉均现采现用。
参考邓衍明等[14]的方法对达乌里胡枝子的花粉进行离体培养。待花粉散粉后,在单凹载玻片凹槽上方用尖头镊子和解剖针反复刮取花丝上的花粉,确保花丝上的花粉全部落入凹槽内,取适量液体培养基质,在凹槽部位滴2滴,将载玻片置于底部垫有2层浸透水的滤纸的培养皿中,盖上培养皿盖子后置于恒温培养箱(温度25℃±1℃,相对湿度60%)内培养[6]。
1.2.2培养条件筛选 进行单因素试验,设置蔗糖浓度5个处理(0,50,100,150,200 g·L-1),硼酸浓度5个处理(0,20,40,80,120 mg·L-1),氯化钙浓度5个处理(0,10,30,50,80 mg·L-1),其中蒸馏水作对照,记为0(CK)。每个处理重复3次,培养时间为4 h和6 h。分别将上述不同浓度的蔗糖、硼酸和氯化钙溶液添加到液体培养基中,找出花粉萌发和花粉管生长的最适浓度和最佳培养时间。
在单因素的基础上分别选取对达乌里胡枝子影响较大的蔗糖、硼酸和氯化钙各2个浓度水平采用3因素2水平试验,确定适宜达乌里胡枝子的最佳处理组合,每个处理重复3次。
在最适培养基的基础上,分别在光照培养箱和冰箱中进行不同培养温度20℃,25℃,30℃,35℃和40℃)和不同贮藏温度(-20℃,4℃)试验,在之前筛选的条件下进行不同采样时间(8:00,12:00,18:00)以及光照(40 μmol·(m2·s)-1)和黑暗(0 μmol·(m2·s)-1)两种光照环境的筛选,每个处理设3次重复,最终确定花粉培养的最适温度、采样时间和光照条件。
1.2.3花粉萌发测定 每个玻片随机选取5个不重复的视野,每个视野花粉数量不少于50粒,每片随机测量20个花粉管,以视野内花粉管长度大于花粉直径为花粉萌发标准[15]。用显微测微尺测量花粉管长度,统计和计算平均萌发率和花粉管长度。
花粉萌发率=已萌发的花粉数/花粉粒总数×100%。
1.3 数据分析
利用Excel 2010对测定的数据进行整理,用SPSS 23.0软件对数据进行单因素方差分析(One-way ANOVA),并运用Duncan法对各个处理之间的指标进行差异性显著分析,运用Origin 2017软件作图。
2 结果分析
2.1 培养基组分对达乌里胡枝子花粉离体萌发的影响
2.1.1蔗糖浓度对花粉离体培养的影响 各浓度培养4 h的花粉萌发率和花粉管长度均低于培养6 h,且花粉萌发率随着蔗糖浓度的升高呈先升后降的趋势(图1)。培养6 h后,各处理组的花粉萌发率和花粉管长度均高于CK,浓度为15 g·L-1时花粉萌发率和花粉管长度均高于其他处理组;10 g·L-1和20 g·L-1处理的花粉萌发率无显著差异,其中20 g·L-1处理的花粉管长度高于10 g·L-1;5 g·L-1处理的萌发率和花粉管长度略高于CK,可以看出过高或过低的蔗糖浓度都不利于花粉萌发,但花粉管长度在浓度为0~15 g·L-1的范围内呈现出持续升高的趋势。由此可知,培养基中添加浓度15 g·L-1和20 g·L-1的蔗糖,能有效提高花粉萌发率和花粉管长度,达乌里胡枝子花粉最适培养时间为6 h,最适蔗糖浓度为15 g·L-1或20 g·L-1。
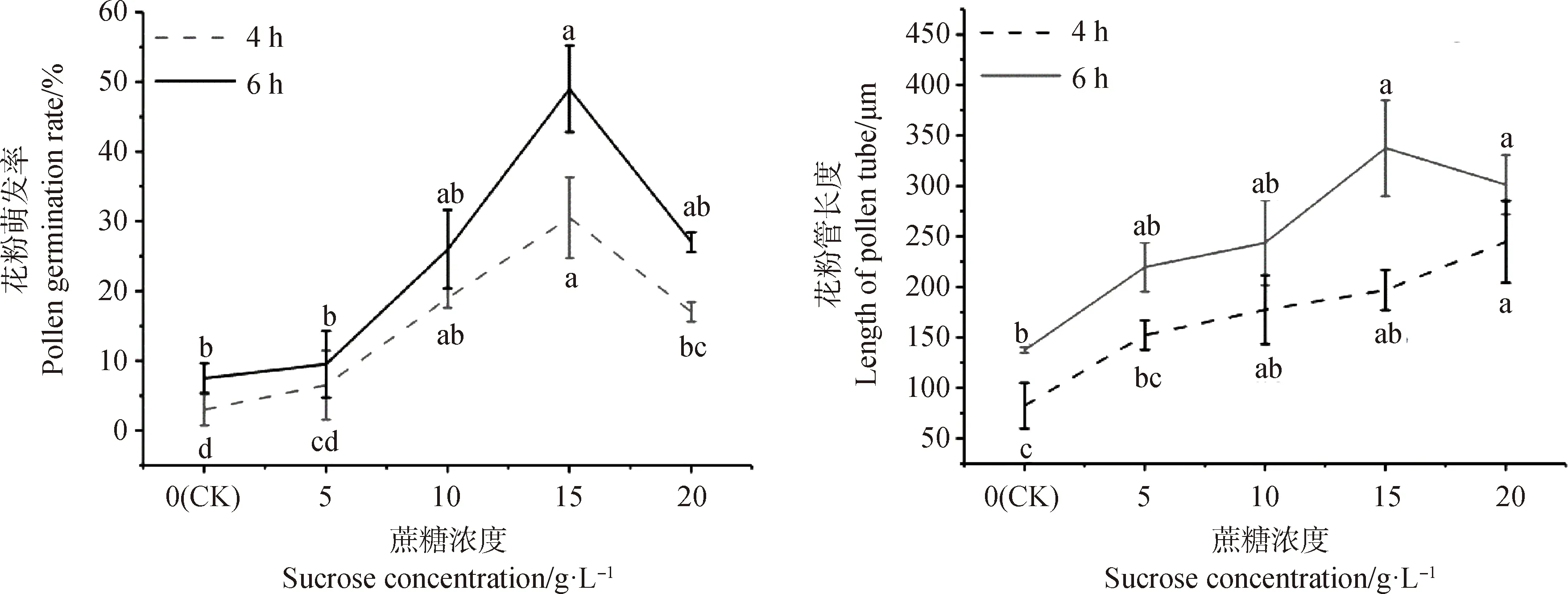
图1 蔗糖对达乌里胡枝子花粉萌发和花粉管生长的影响Fig.1 Effects of sucrose on Pollen Germination and pollen tube growth of Lespedeza davurica注:不同小写字母表示差异显著(P<0.05),下同Note:Different lowercase letters show significant differences at the 0.05 level. The same as below
2.1.2硼酸浓度对花粉萌发率及花粉管长度的影响 花粉萌发率和花粉管长度随着硼酸浓度的升高呈先升后降的趋势,且培养6 h后的花粉萌发率和花粉管长度均高于培养4 h(图2)。培养6 h后,CK的花粉萌发率和花粉管长度最低,随着浓度的增加,20 mg·L-1和40 mg·L-1处理的花粉萌发率和花粉管长度均无显著差异,且20 mg·L-1处理的花粉萌发率高于其他处理组,40 mg·L-1处理的花粉管长度和其他处理组相比最长;浓度为80 mg·L-1和120 mg·L-1处理的花粉萌发率与CK相比均无显著差异,且组间无显著差异,但其花粉管长度均高于CK。综上,花粉的最适培养时间为6 h,硼酸最适浓度为20 mg·L-1或40 mg·L-1。
2.1.3氯化钙浓度对花粉萌发率及花粉管长度的影响 花粉萌发率随着氯化钙浓度的升高呈先升后降的趋势,且培养6 h后的花粉萌发率和花粉管长度均高于培养4 h(图3)。培养6 h时,在氯化钙浓度为0~50 mg·L-1的范围内,花粉萌发率随着浓度的增加而增加。氯化钙浓度为50 mg·L-1时花粉萌发率显著高于CK和除80 mg·L-1处理外的其他处理组(P<0.05),达到68%;80 mg·L-1时,花粉萌发率有所降低,但高于10 mg·L-1和30 mg·L-1处理组。花粉管长度随着浓度的增加呈现出不规则变化,其中氯化钙浓度为10 mg·L-1,50 mg·L-1和80 mg·L-1的花粉管长度均高于30 mg·L-1和CK处理组,且3个处理组间无显著差异,其中50 mg·L-1处理的花粉管最长,为187.04 μm。综上,适宜用于测定达乌里胡枝子花粉活力的氯化钙浓度为50 mg·L-1和80 mg·L-1。
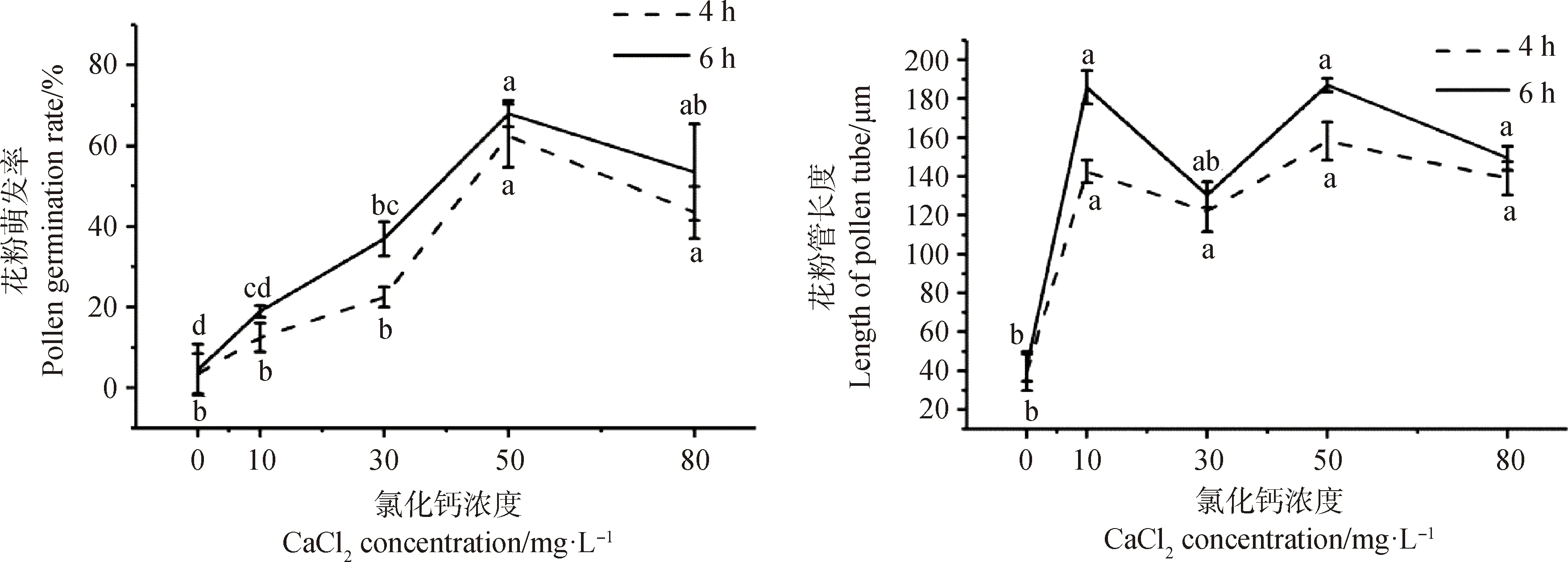
图3 氯化钙对达乌里胡枝子花粉萌发和花粉管生长的影响Fig.3 Effects of CaCl2 on Pollen Germination and pollen tube growth of Lespedeza davurica
2.1.4最佳培养基选择 在单因素试验的基础上,将蔗糖、硼酸和氯化钙设为3个因素,将每个因素筛选出的2个浓度设为水平,采用3因素2水平试验(表1),寻找最佳培养基配方。
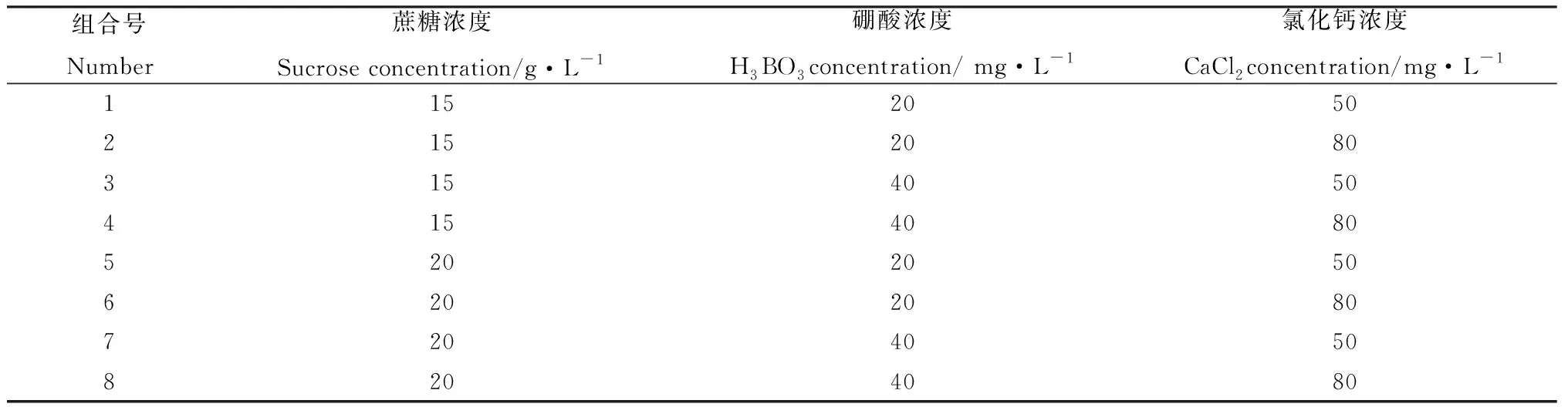
表1 试验因素及浓度水平Table 1 Test factors and concentration levels
8个处理组间的花粉萌发率和花粉管长度有明显的差异(图4)。其中,组合7的花粉萌发率最低,组合2,3和5的萌发率次之,且2,3和5处理组组间差异不显著;组合1,4和6的萌发率较高,且组间差异不显著;组合8的花粉萌发率和花粉管长度均显著高于其他处理组(P<0.05),且花粉长势良好,萌发率为69%,花粉管长度为263.88 μm;组合3和6的花粉管长度显著低于其他处理组(P<0.05),其中与组合5的差异不显著;组合1,2,5的花粉管长度组间差异不显著,且低于组合4和7,其中组合4花粉管长度略低于组合8,为213.11 μm。基于对达乌里胡枝子花粉萌发率和花粉管长度的影响,最优组合培养基为组合8,故后续试验的培养基配方均以组合8的配方进行开展。
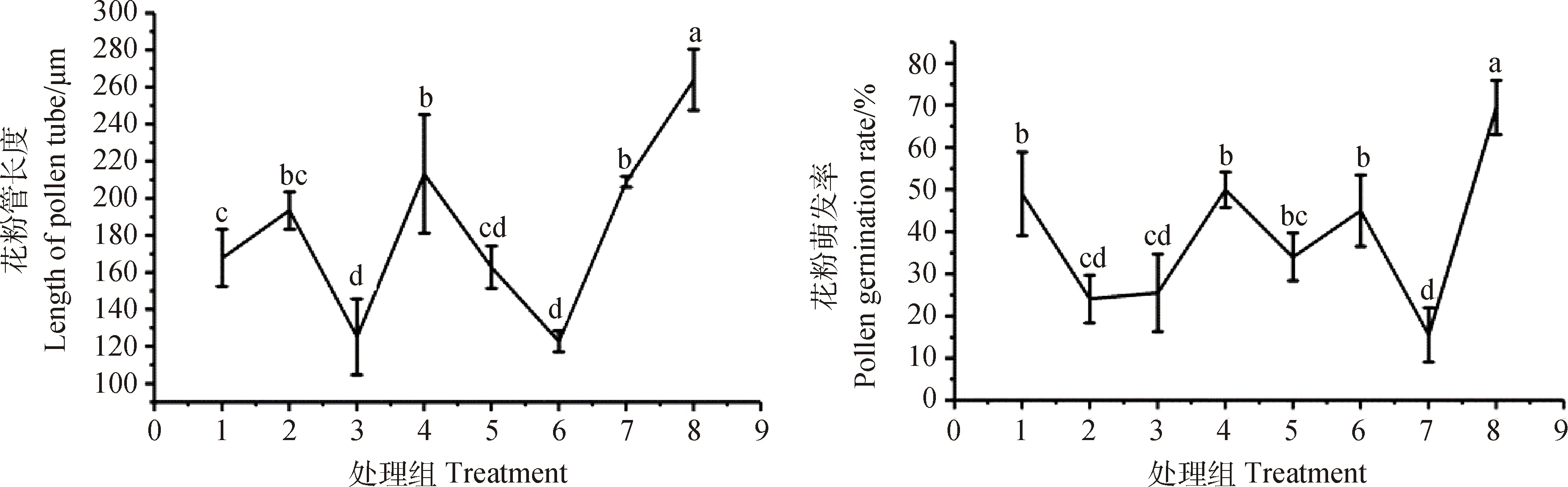
图4 不同培养基处理组对达乌里胡枝子花粉萌发和花粉管生长的影响Fig.4 Effects of different media treatments on pollen germination and pollen tube growth of Lespedeza davurica by different media treatment

图5 达乌里胡枝子花粉萌发及花粉管生长的动态变化Fig.5 Dynamic changes of pollen germination and pollen tube growth of Lespedeza davurica注:a,蒸馏水中培养6小时;b,培养基中培养6小时;c,花粉萌发初始阶段;d-e,花粉管伸长,内含物质进入花粉管;f,花粉3个萌发沟最终只有一条花粉管伸长;g-i,花粉管异常,出现扭曲、内含物散出、花粉管分支和缠绕成束Note:a,Culture in distilled water for 6 hours;b,Culture in medium for 6 hours;c,Initial stage of pollen germination;d-e,Pollen tube elongation,and the contents enter into the pollen tube;f,Only one pollen tube is elongated in the three pollen germination grooves;g-i,Abnormal pollen tube,distortion,content dispersion,pollen tube branching and twining into bundles
在显微镜中观察到,达乌里胡枝子花粉为圆球形,直径大约为50~70 μm(图5a),通常一粒花粉只有一条花粉管(图5b),萌发初期3个萌发孔均能萌发(5c),但最终只有一个萌发孔的花粉管伸长(图5f)。萌发初期,萌发沟处有透明的突起物(图5c),随着花粉管的伸长生长,花粉管中散布着内含物(图5-d-e),最终全部集中在花粉管的顶端。在花粉管的生长过程中,花粉管会出现解体,内含物散出、花粉管扭曲、花粉管有多个分支和众多花粉管缠绕成束的现象(图5 g-i)。
2.2 培养条件对达乌里胡枝子萌发生长的影响
不同采样时间和光照条件对花粉萌发率和花粉管长度的影响也不同(图6)。在相同培养基上,暗培养对花粉萌发率的影响在各个时间段均高于光照培养,其中,以中午12点的花粉萌发效果最佳,其次为早上8点;下午18点的花粉萌发率与早上8点和中午12点相比显著下降(P<0.05),且出现花粉内含物渗出的现象。在暗培养的条件下,中午12点的花粉管长度最长,为223 μm,早上8点的花粉管长度最短,故选择中午12点作为达乌里胡枝子花粉的最佳取样时间。
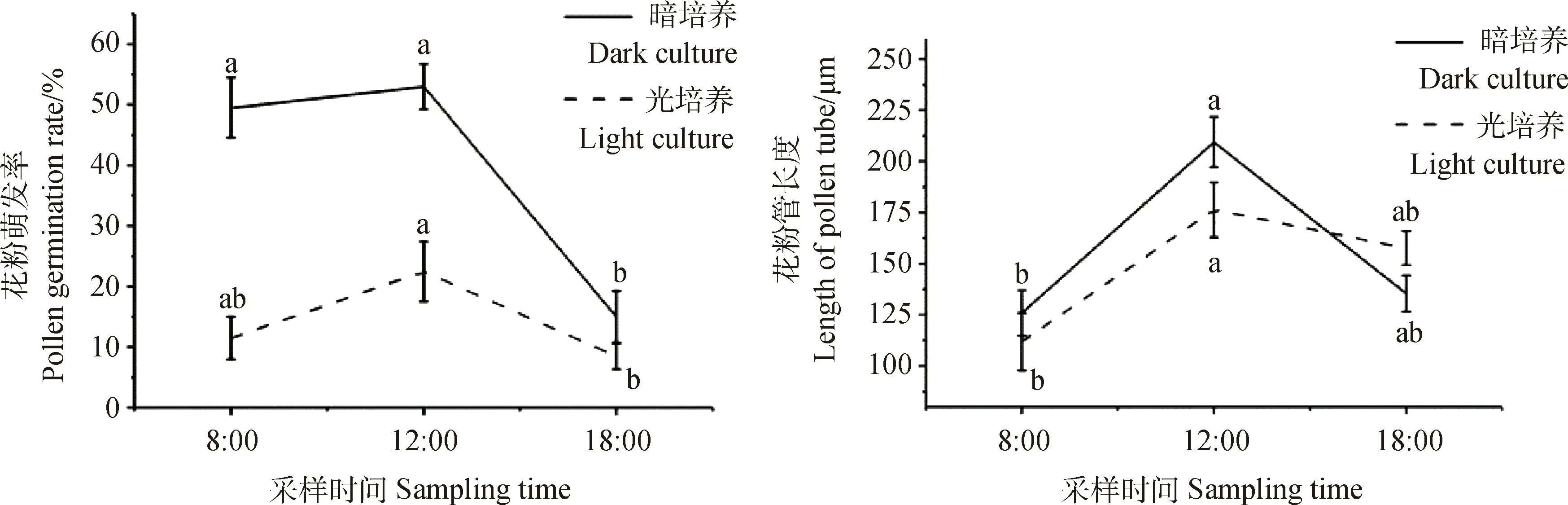
图6 培养条件对达乌里胡枝子花粉萌发及花粉管生长的影响Fig.6 Effects of culture conditions on Pollen Germination and pollen tube growth of Lespedeza davurica
2.3 不同培养温度及贮藏条件对达乌里胡枝子萌发生长的影响
不同培养温度显著影响达乌里胡枝子花粉的萌发生长(P<0.05)。总体来看,温度在20℃~40℃的范围内时,花粉萌发率和花粉管长度随着温度的升高呈现先升高后降的趋势(图7)。在20℃~25℃内,花粉萌发率和花粉管长度均随着温度的升高而升高,萌发率在25℃时达到最高;在25℃~40℃之间,随着温度的升高花粉萌发率和花粉管长度呈下降趋势,40℃时花粉萌发率和花粉管长度均最低。贮藏温度为4℃的花粉萌发率显著高于—20℃(P<0.05),但花粉管长度却显著低于—20℃(P<0.05)。由此可知,达乌里胡枝子花粉的最佳贮藏温度为—20℃,最佳培养温度为25℃。
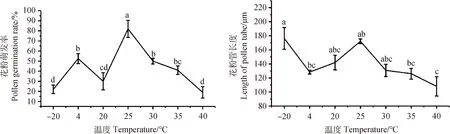
图7 培养温度及贮藏条件对达乌里胡枝子花粉萌发及花粉管生长的影响Fig.7 Effects of culture temperature and storage conditions on Pollen Germination and pollen tube growth of Lespedeza davurica注:-20℃和4℃为贮藏温度;20℃~40℃为培养温度试验Note:-20℃ and 4℃ is the storage temperature;20℃~ 40℃ is the culture temperature test
3 讨论
花粉离体萌发法具有结果准确、简单易行的优点。除了受自身条件限制外,花粉的萌发在很大程度上受其培养基成分、温度和光照条件等的影响,只有在适宜的外界条件下,成熟的花粉才能正常的萌发及生长。
蔗糖能通过调节花粉渗透压从而促进其萌发和生长,但过高的浓度会使花粉管渗透压不平衡导致破裂,影响萌发[16]。研究发现,蔗糖浓度对花粉萌发的影响具有显著性[17]。花粉萌发对蔗糖浓度的需求根据不同的植物而有所差异,一般以质量分数10%~25%居多[18-19],低浓度的如二乔玉兰(Magnoliasoulangeana)为5%[20],而贯月忍冬(Lonicerasempervirens)花粉在蔗糖溶液浓度为30%时萌发率最高[21]。尉倩等[22]研究表明蔗糖浓度过高会导致渗透势下降,花粉因失水过多而萌发率下降,这与本试验的研究结果相似。本试验中蔗糖浓度过高或偏低都会使达乌里胡枝子花粉萌发率降低,花粉的萌发率和花粉管长度均随蔗糖浓度的增加呈先升后降的趋势,其中15 g·L-1为峰值,说明在10~15 g·L-1的范围内蔗糖能为花粉萌发生长提供充足的营养物质;当蔗糖浓度为20 g·L-1时,花粉萌发率出现下降趋势,但其花粉管长度无显著降低,可能是因为花粉粒个体间原生质不同组分的含量存在差异,导致部分花粉在较高的渗透压环境中仍然能萌发和生长,从而出现抑制花粉管生长的质量分数也相应升高的现象[23]。
硼作为植物有性生殖过程不可缺少的微量元素,在花粉萌发生长等方面都发挥着重要的作用[18],花粉缺硼可导致花粉管破裂[24]。在自然生长条件下,花粉中的硼是不足以维持自身萌发生长的,但柱头和花柱中的硼可以补偿这种不足,确保花粉可以在柱头上正常生长[25]。吴斌[17]研究发现,低浓度的硼酸能促进小黄金西番莲(Passifloraedulis)花粉萌发,当硼酸浓度过高时通过影响花粉萌发进而导致产量下降,所以适量浓度的硼酸有助于花粉萌发和花粉管生长。本研究发现,硼酸浓度为20~40 mg·L-1时,会促进花粉萌发和花粉管生长,当浓度为80~120 mg·L-1时,花粉的萌发生长则会受到抑制,这与前人[10]的研究结果相似。但本研究中花粉萌发和花粉管生长所需的最适硼酸浓度分别是20 mg·L-1和40 mg·L-1,造成浓度不同的原因可能是花粉自身的硼酸可以满足萌发的需求,但不足以维持花粉管的伸长,而硼是构建花粉管顶端细胞壁不可缺少的物质,故花粉管生长相对于花粉萌发所需的外源硼量较多。因本试验设置的硼酸浓度范围较窄,故抑制达乌里胡枝子花粉管生长的浓度尚待进一步探究。
Ca2+对促进花粉萌发生长具有重要作用[26]。研究发现在培养基中加入Ca2+通道阻塞剂会干扰花粉萌发[27];外源Ca2+与花粉细胞内游离钙存在互补关系,当自身游离钙缺乏,适量的外源Ca2+能有效促进花粉萌发生长,但外源Ca2+浓度过高时会导致细胞膨压抑制花粉萌发生长[28]。芦娟[16]研究发现Ca2+浓度为40 mg·L-1时能显著提高油橄榄(Oleaeuropaea)的萌发率,当Ca2+浓度为120 mg·L-1时,其花粉萌发率显著降低,且低于对照,这与本试验研究结果相一致。本研究中Ca2+浓度为0~30 mg·L-1时,花粉萌发率无显著变化;当Ca2+浓度为50 mg·L-1时,其花粉萌发率处于峰值;当Ca2+浓度为80 mg·L-1时其花粉萌发率呈下降趋势,说明Ca2+浓度过高或过低均会抑制花粉萌发。但花粉管长度在Ca2+浓度处于10,50和80 mg·L-1时均显著升高,而50 mg·L-1时处于最高,可能是花粉细胞内个体间Ca2+含量存在差异,因此对不同浓度的Ca2+浓度的需求不同,进而导致出现多个花粉管长度峰值。本试验研究结果表明达乌里胡枝子花粉细胞内游离钙含量不均等,故自然条件下,花粉管长度与外源钙的关系有待于后期进行深入研究。
硼酸与蔗糖之间存在交互作用。H3BO3能与蔗糖形成络合物,促进花粉对糖类的吸收、转运和代谢[29-30];硼离子可以通过影响质外体和钙离子的结合,增加胞内游离钙离子至花粉萌发最适浓度[31]。通过对郁香忍冬(Lonicerafragrantissima)花粉进行离体萌发研究,发现单一蔗糖溶液浓度为35%时萌发率最高,而对蔗糖和硼酸进行多因素试验时,30%蔗糖的萌发率达到最高[22]。本试验中,单因素蔗糖浓度为15 g·L-1花粉萌发率最高,而在多因素试验中,组合8的蔗糖20 g·L-1+硼酸40 mg·L-1+氯化钙80 mg·L-1的处理能显著促进花粉萌发和花粉管生长,加入硼酸后,需要的蔗糖浓度也相应增加,可能是因为花粉萌发初期所需蔗糖和硼量较少,硼作为花粉管顶端构建不可缺少的元素,因此在花粉管伸长生长的过程中需要的蔗糖和硼酸量较多[10]。因本实验设置的浓度较少,花粉萌发率下降趋势未体现,故后续应对组合中各组分配比进一步探究。
不同植物花粉离体培养最适采样时间以及培养条件均会有所不同。邓衍明等[14]通过对茉莉(Jasminumsambac)花粉进行离体培养发现,早上花粉活力显著高于中午和晚上的活力;吕晋慧等[32]研究发现地被菊(groundcoverchrysanthemum)的花粉活力在中午13:00的时候最高,与本试验研究结果相似。本研究发现,8:00~12:00的花粉萌发率显著高于18:00,且中午得萌发率最高,故中午花粉活力最好,花粉管长度也以中午12:00所取的花粉最长,可能是因为不同的时间段所处的外界环境中的温度、湿度对花粉活力的影响较大[14]。李佛莲等[33]研究发现黑暗条件下山玉兰(Magnoliadelavayi)花粉萌发率高于光照条件,这与本试验的研究结果相一致,本研究发现达乌里胡枝子花粉更适宜在黑暗条件下萌发,光照条件下更适宜花粉管生长。
温度是影响花粉萌发的重要因素。适宜的温度能促进花粉萌发,温度过高或过低均会抑制其萌发生长[34],且植物不同,花粉萌发所需的最适温度也不同。研究发现对于温带植物来说25℃~30℃为适宜温度,低于25℃或高于35℃其萌发则会受到抑制[14];还有研究发现地梢瓜(Cynanchumthesioides)[35]最适萌发温度为30℃,而地被菊花粉的最适萌发温度为10℃或15℃[32];杨澜等[36]通过对多肉植物红司(Echeverianodulosa)花粉离体萌发研究发现,温度与花粉萌发率呈负相关,这与本试验研究结果一致。本研究发现温度在25℃~40℃范围内时,花粉萌发率和温度呈负相关,且25℃时花粉萌发率最高,说明达乌里胡枝子花粉对高温具有一定的耐受性。
合适的贮藏条件对花粉活力的保持至关重要。一般花粉贮藏温度有常温、4℃,—20℃,—70℃,—80℃和—196℃[37]。实际生产中常温、4℃和—20℃由于所需条件简单故经常用到。朱雪晨[38]研究发现—20℃和—70℃均为薄壳山核桃(Caryaillinoinensis)的适宜贮藏温度,这与本试验研究结果一致。本研究发现,—20℃花粉萌发率显著低于4℃,但—20℃下花粉管长度显著高于其他处理组,可能是因为低温导致花粉酶活性减弱,生活力下降,花粉萌发率降低,而花粉管生长主要受硼的影响,可能低温未对其伸长生长造成影响。关于低温贮藏环境和贮藏时间对达乌里胡枝子花粉萌发和花粉管生长和各生理机制的影响,有待于后续深入研究。
4 结论
本研究综合单因素和多因素试验结果分析和比较,得出‘晋农一号’达乌里胡枝子花粉离体萌发的最适液体培养基配方为20 g·L-1蔗糖+40 mg·L-1硼酸+ 80 mg·L-1氯化钙,最适采样时间为中午12:00,最适培养条件为黑暗条件下25℃,最适贮藏温度为—20℃,最佳镜检时间为6 h。
