高寒地区人工草地土壤微生物群落对不同种植方式和年限的响应
2021-04-08张杰雪王占青全小龙史惠兰陈梦词乔有明
张杰雪, 王占青, 全小龙, 梁 军, 史惠兰, 陈梦词, 乔有明*
(1. 青海大学省部共建三江源生态和高原农牧业国家重点实验室, 青海 西宁 810016;2. 青海大学农牧学院, 青海 西宁 810016;3. 青海大学生态环境工程学院, 青海 西宁 810016)
土壤微生物是土壤生态系统的重要组成部分,参与并推动环境中物质与能量的流动和转换[1]。健康的土壤微生物群落不仅有助于维持土壤生态系统的稳定,还能提高土壤对生态环境恶化的缓冲能力,维护生态平衡[2-3]。由于自然和人为因素,人工草地和天然草地一样面临着退化的问题。人工草地的退化不仅会引起草地植物群落结构变化和牧草产量、质量下降[4],还会导致土壤养分、土壤微生物群落结构等变化[5-7]。研究高寒地区多年生人工草地土壤微生物多样性变化对保护和管理该地区人工草地、改善土壤状况、保持土壤稳定性具有重要实践意义。
植物群落可通过其根际的生理过程以及根系分泌物等化感物质影响土壤微生物群落的结构和组成,而土壤微生物也会通过其自身的生理生化作用或分泌一些激素等物质对植被产生影响[8]。通过对川西北高寒牧区紫花苜蓿(Medicagosativa)人工草地、燕麦(Avenasativa)人工草地以及天然草地土壤微生物多样性研究,伍文宪等[9]认为草地植被类型可改变地下土壤微生物的群落结构。有研究发现,不同种植方式或不同生长年限的人工草地之间土壤微生物特性及其养分含量也存在显著差异[10-11]。随着生长年限的增长,细菌逐渐减少,而真菌逐渐增加,进一步影响土壤肥力和生态系统平衡[12]。这些研究主要利用传统的培养方法或采用磷脂脂肪酸(Phospholipid fatty acid,PLFA)法对微生物进行测定,只能判断细菌、放线菌和真菌3大类的变化情况,不能鉴别到门及以下的分类水平。
高通量测序技术根据细菌16S和真菌ITS区域,经提取、扩增、纯化、定量、均一化细菌和真菌的基因序列,再经测序、过滤、优化、聚类,对比基因库中的已知序列,从而鉴别细菌和真菌种(属)类,并可鉴别到那些室内不可培养的微生物种类,是传统培养方法所获得微生物数量的十倍甚至数百倍,能准确灵敏地检测土壤细菌和真菌。青海高寒地区人工草地建植和退化草地的恢复主要通过种植多年生禾本科牧草,如垂穗披碱草(Elymusnutans)、草地早熟禾(Poapratensis)、中华羊茅(Festucasinensis)和冷地早熟禾(Poacrymophila)等。除种子生产外,绝大多数采用混播方式,因管理利用不当或土壤肥力和化感等方面的因素,存在人工草地种植2~3年后快速退化的问题[13]。为此,本研究采用高通量测序技术,从土壤微生物群落的角度,研究土壤微生物在牧草不同种植方式下随生长年限的变化,以期揭示高寒地区人工草地退化的潜在驱动原因。
1 材料与方法
1.1 研究地概况
研究地点位于青海省牧草良种繁殖场(35°09′ N,100°39′ E),属于青海省海南州同德县巴滩地区,总面积243 km2,其中耕地面积100 km2,天然草地129 km2。海拔3 300~3 500 m,年均降水量429.8 mm,年均温0.2℃,牧草生长季内活动积温1 309℃,无绝对无霜期。冬春干旱、寒冷,夏季凉爽,雨热同季,有利于优良牧草生长。场内地势平坦,土层厚度70~150 cm,土壤为暗栗钙土,适宜种植优良牧草种子繁殖,如垂穗披碱草、冷地早熟禾、中华羊茅、‘同德’老芒麦(Elymussibiricus‘Tongde’)等。青海省牧草良种繁殖场于每年五月中下旬播种,种子田播种前施底肥磷酸二铵150 kg·km-2和尿素112.5 kg·km-2,每年八月下旬进行种子收获。种子生产田中牧草生长3年后更换种植的牧草种类,多年生人工草地在前3年有施肥,3年后不再进行施肥,无除草、追肥等管理措施。
1.2 样品采集
于2019年8月底在青海省牧草良种繁殖场进行样品采集,采样前3天未下雨,每个样地面积为100 m × 200 m。其中单播处理为种子生产田,混播处理为多年生人工草地。调查并记录每个样地种植的牧草种类等信息(表1)。在每个样地随机选取5个采样点进行土壤取样,样点相隔距离15 m以上,采集土壤深度为0~20 cm的土壤。每个样点分别采集土壤微生物分析样品和化学性质分析样品,微生物分析样装于无菌管中,放入—4℃的便携式冰箱中,带回实验室保存于—20℃的贮藏箱中;化学性质土样装入自封袋带回实验室,风干过筛后进行土壤pH、有机碳、硝态氮、铵态氮和速效磷含量测定。
1.3 测定方法及指标
1.3.1土壤化学性质的测定 土壤酸碱度用pH计(PHS-3E)测定,土壤有机碳含量用重铬酸钾-硫酸容量法[14]测定,总氮含量用凯氏定氮法测定,硝态氮含量用氯化钾-硫酸肼还原法测定,铵态氮含量用氯化钾-水杨酸钠法测定,速效磷含量用碳酸氢钠-钼锑抗比色法测定[15]。以每个样地的5个样品作为土壤化学性质测定的5次重复。

表1 样品采集地信息
1.3.2土壤微生物的测定 从—20℃的贮藏箱中取出装有土样的无菌管,在超净工作台中取无菌管中的土壤5 g左右,去除植物根和石砾后装于5 mL的无菌管中,放置在装有干冰的泡沫箱中寄到诺禾致源公司天津总部,采用16S/ITS扩增子测序分析技术测定土壤微生物的多样性。每个样地随机选取了3个样品,即3次重复。不同处理的样品编号见表1。
1.4 数据处理与分析
土壤化学性质数据利用SPSS 20.0进行单因素方差分析(One-way ANOVA),并采用最小显著性差异法(LSD)进行处理间差异显著性比较。对同一种植方式不同生长年限土壤的微生物数据进行合并处理,然后进行比较。微生物数据的作图通过R3.6.1完成,RDA分析用Canoco 5.0完成。
2 结果与分析
2.1 土壤化学性质
本研究的6个样地土壤化学性质如表2所示,土壤pH在8.5~9.0之间,偏碱性。土壤总氮含量介于2.17~2.75 g·kg-1之间,有机碳含量介于13.05~22.28 g·kg-1之间,速效磷含量介于9.78~37.12 mg·kg-1之间。中华羊茅和短芒披碱草单播草地不同生长年限土壤有机碳、总氮和铵态氮含量无显著差异,硝态氮和速效磷含量随生长年限增长显著降低(P<0.05);混播草地土壤总氮含量无显著变化,土壤有机碳、铵态氮和硝态氮含量随生长年限增长显著降低(P<0.05),速效磷显著增加(P<0.05)。
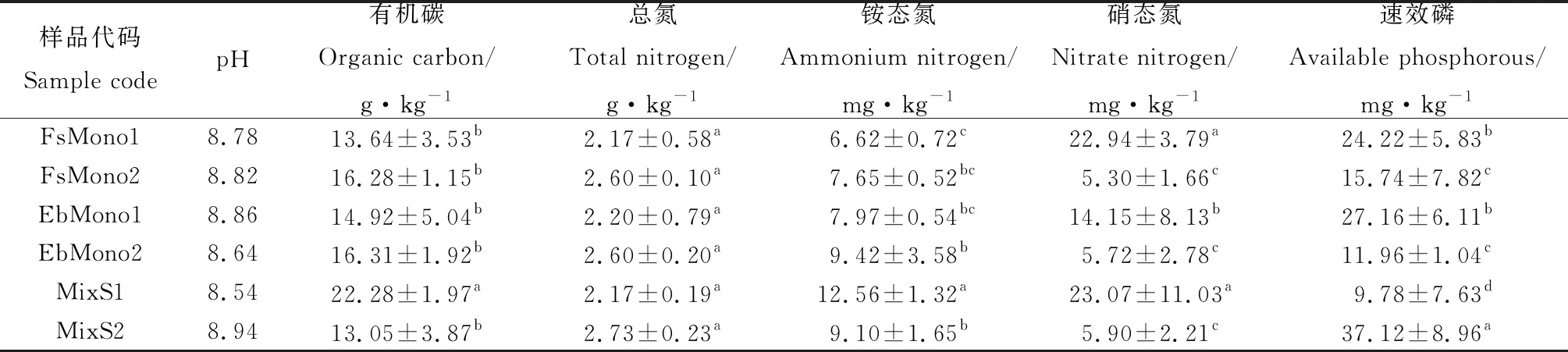
表2 土壤的基本化学性质
2.2 土壤微生物群落比较
2.2.1微生物OTU数目 细菌OTU聚类分析发现:不同种植方式土壤共有7 963个OUT,共同包含的OTU数为4 526个,单播草地和混播草地土壤的OUT均在6 100~6 300个之间,没有表现出明显的差异。6个样地土壤共同包含的OTU数为2 763个(图1)。无论是单播草地还是混播草地,土壤细菌种类随生长年限而减少,即FsMono1 > FsMono2,EbMono1 > EbMono2,MixS1 > MixS2。
真菌OTU聚类分析表明,不同种植方式土壤共有5 609个OUT,共同包含的OTU数为1 608个,中华羊茅和中华羊茅+短芒披碱草+冷地早熟禾混播草地土壤的真菌OUT数在3 500~3 600个之间,而短芒披碱草单播草地的土壤真菌OUT数明显少于前两者,不到3 000个。6个样地土壤共同包含的真菌OTU数有658个(图1)。种植中华羊茅的草地土壤真菌类群数量较接近,一致性高;但短芒披碱草和混播群落土壤真菌类群呈现随生长年限增长而增加的趋势,即土壤中所含OTU数MixS2 > MixS1,EbMono2 > EbMono1。
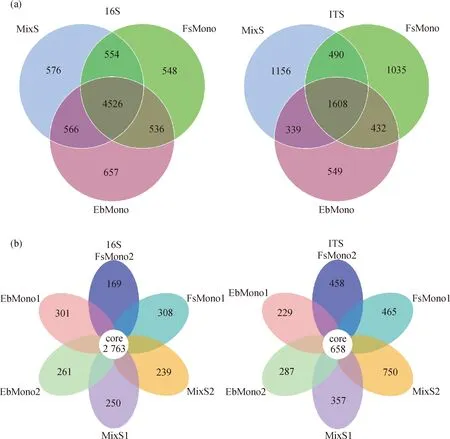
图1 土壤细菌(16S)和真菌(ITS) OTU数量的韦恩图
2.2.2微生物多样性 土壤微生物的多样性结果显示,不同种植方式间的土壤细菌Shannon指数和Chao1指数均没有显著差异,中华羊茅单播草地土壤细菌丰富度随生长年限增长而减小(表3)。土壤真菌Shannon指数以混播草地最高,中华羊茅单播草地次之,混播草地土壤真菌多样性显著高于短芒披碱草草地的(P<0.05),土壤真菌丰富度Chao1指数以中华羊茅单播草地最高,混播草地次之,混播草地土壤真菌丰富度显著高于短芒披碱草草地的(P<0.05),同一种植方式下生长年限对土壤真菌多样性和丰富度无显著影响(表4)。表3与表4的结果显示各样点的覆盖率均在99.0%以上,说明各样地的微生物物种信息基本得到了充分的体现。
2.2.3微生物群落结构 细菌分析(16S)共检测出63个门,68个纲,143个目,270个科,684个属,478个种,对细菌OTU的注释结果显示,在所有的处理中,变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、酸杆菌门(Acidobacteria)、厚壁菌门(Firmicutes)、拟杆菌门(Bacteroidetes)、绿弯菌门(Chloroflexi)、芽单胞菌门(Gemmatimonadetes)、奇古菌门(Thaumarchaeota)、疣微菌门(Verrucomicrobia)和己科河菌门(Rokubacteria)等10个门的细菌占了所有细菌的95%,其中,变形菌门、放线菌门和酸杆菌门占了大约67%的比例(表5),是不同生长年限的人工草地土壤中的优势细菌。在变形菌门中,α变形菌(Alphaproteobacteria)占比最高,其次为γ变形菌(Gammaproteobacteria),而δ变形菌(Deltaproteobacteria)占比最低。不同种植方式和年限土壤细菌占比分析显示(表5,表6),不同种植方式单播和混播草地土壤细菌门无显著差异,短芒披碱草单播草地土壤短波单胞菌属(Brevundimonas)显著高于中华羊茅单播草地和混播草地(P<0.05)。混播草地土壤绿弯菌门占比随生长年限显著增加(P<0.05),短芒披碱草单播草地短波单胞菌属和芽孢八叠球菌属(Sporosarcina)占比随生长年限显著减小(P<0.05),中华羊茅单播草地和混播草地土壤细菌门和属无显著变化。鞘氨醇单胞菌属(Sphingomonas)为6个样地土壤共同拥有的优势菌属。
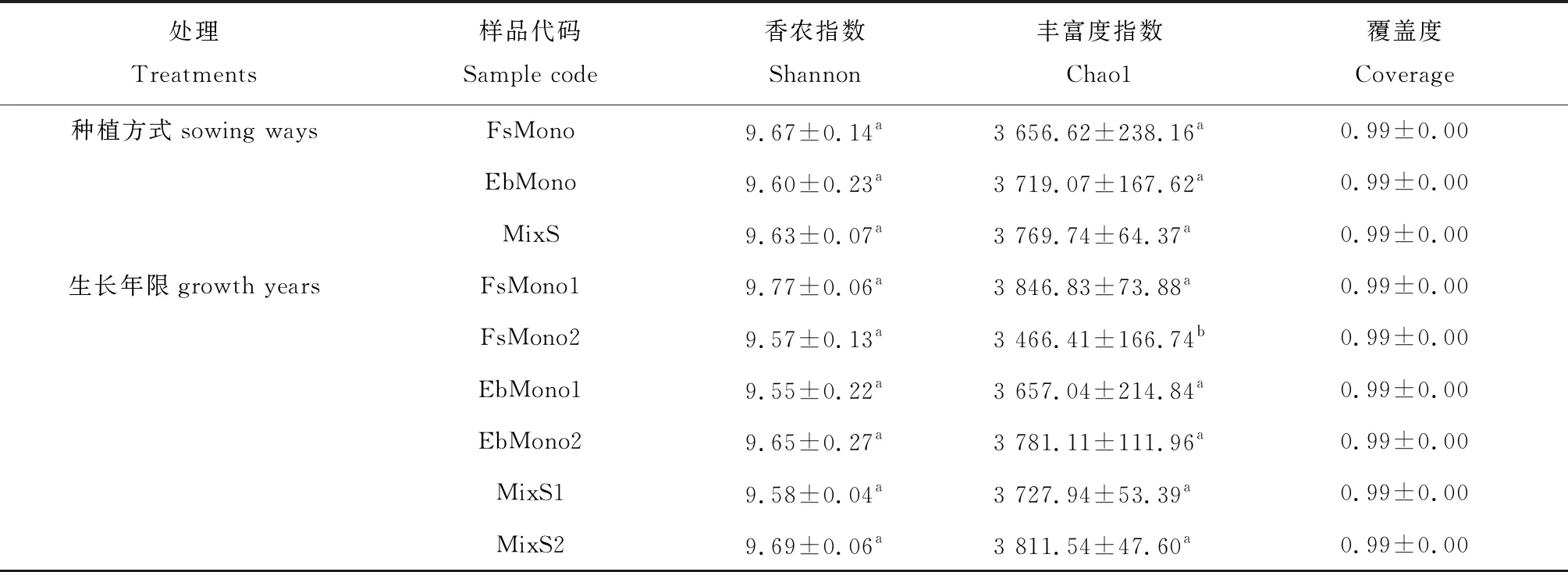
表3 不同种植方式和年限土壤细菌多样性指数比较
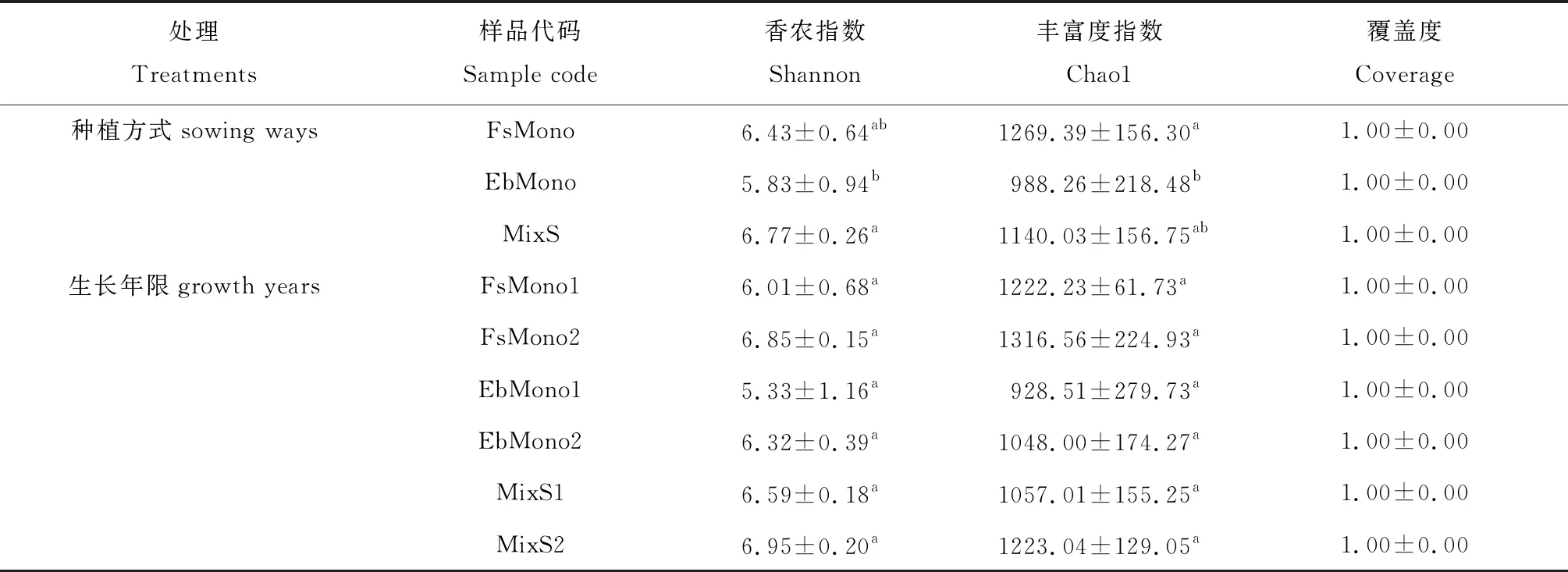
表4 不同种植方式和年限土壤真菌多样性指数比较
真菌分析(ITS)共检测出17个门,56个纲,128个目,275个科,558个属,646个种,真菌在样品间的变异较大,在所有处理中,子囊菌门(Ascomycota)为绝对的优势门,约占到了所有真菌的31%,担子菌门(Basidiomycota)位居其次,所占比例大约为10%,被孢霉门(Mortierellomycota)约占7%,还有约50%未鉴定到门水平的真菌,而其余的门所占比例不到2%。不同种植方式和年限土壤真菌占比分析显示(表5,表6),混播草地子囊菌门占比显著高于短芒披碱草单播草地(P<0.05),中华羊茅单播草地土壤镰刀菌属(Fusarium)占比显著高于短芒披碱草和混播草地的占比(P<0.05)。中华羊茅单播草地土壤担子菌门的占比随年限显著减小(P<0.05),混播草地土壤镰刀菌属占比随年限显著减小(P<0.05)。
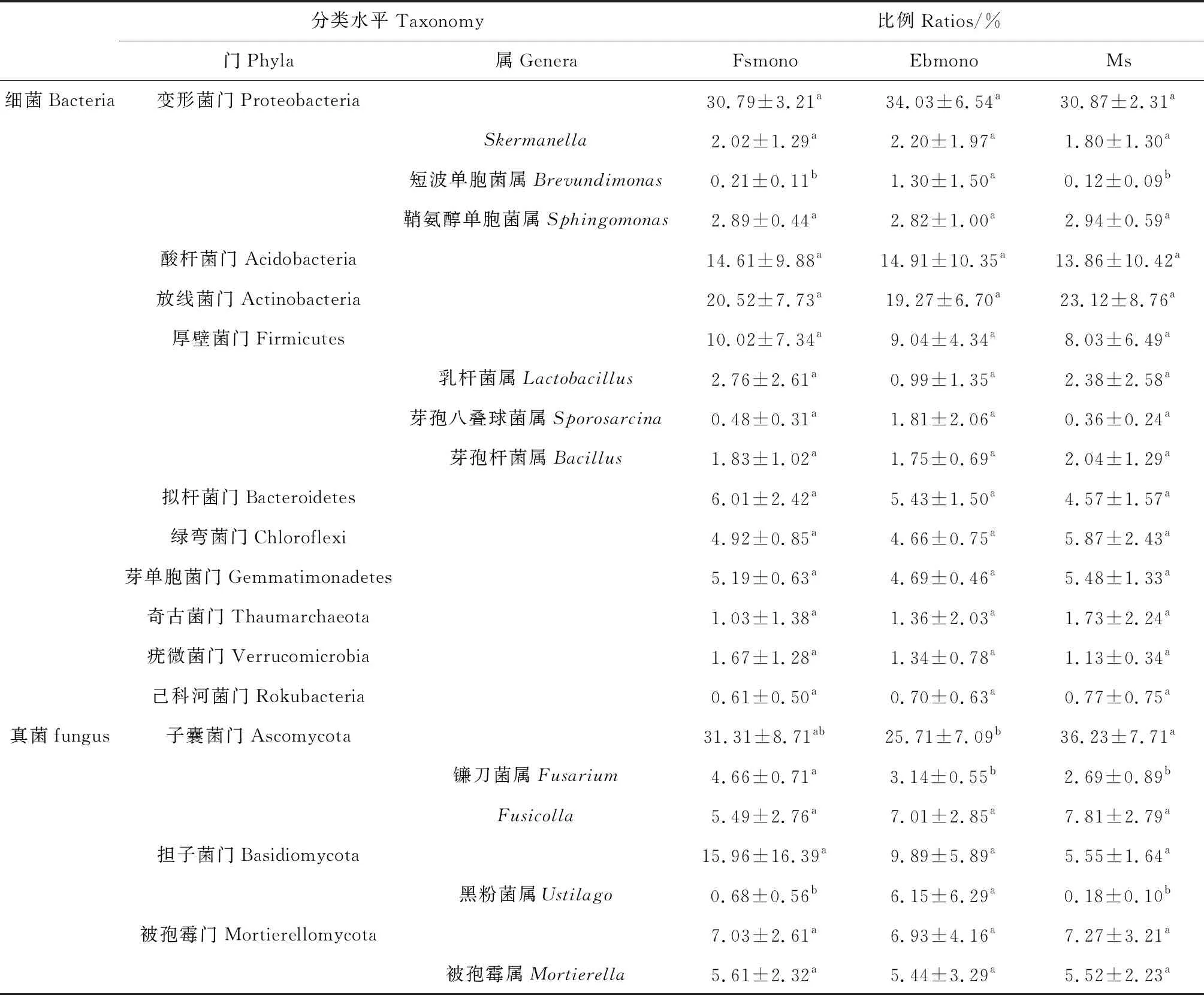
表5 不同种植方式土壤微生物在门和属水平上的丰度变化
2.3 土壤化学性质与微生物群落的关系
土壤细菌群落与土壤化学性质RDA分析显示,轴一和轴二的解释变量分别为27.75%和23.03%(图2),环境因子中土壤pH、总氮、硝态氮和速效磷对细菌群落的影响最大,而土壤铵态氮、有机碳对细菌群落的影响不显著;芽单胞菌门、己科河菌门、奇古菌门和酸杆菌门与土壤总氮、铵态氮和有机碳含量呈正相关,绿弯菌门和放线菌门与土壤pH、速效磷含量呈正相关,变形菌门、厚壁菌门和拟杆菌门与硝态氮含量呈正相关,拟杆菌门与土壤有机碳含量呈正相关。
土壤真菌群落与土壤化学性质RDA分析显示,轴一和轴二的解释变量分别为34.17%和25.97%(图2),土壤pH、总氮、有机碳和硝态氮对真菌群落影响最大,土壤速效磷和铵态氮对真菌群落的影响不显著;担子菌门与土壤pH、硝态氮和速效磷含量呈正相关,与有机碳、铵态氮、总氮呈负相关,壶菌门(Chytridiomycota)、油壶菌门(Olpidiomycota)、芽枝霉门(Blastocladiomycota)和毛霉门(Mucoromycota)与土壤pH呈正相关,土壤总氮含量与子囊菌门、被孢霉门、捕虫霉门(Zoopagomycota)、Aphelidiomycota、球囊菌门(Glomeromycota)、壶菌门、油壶菌门、芽枝霉门和毛霉门呈正相关,子囊菌门、被孢霉门和捕虫霉门与土壤有机碳和铵态氮含量呈正相关。
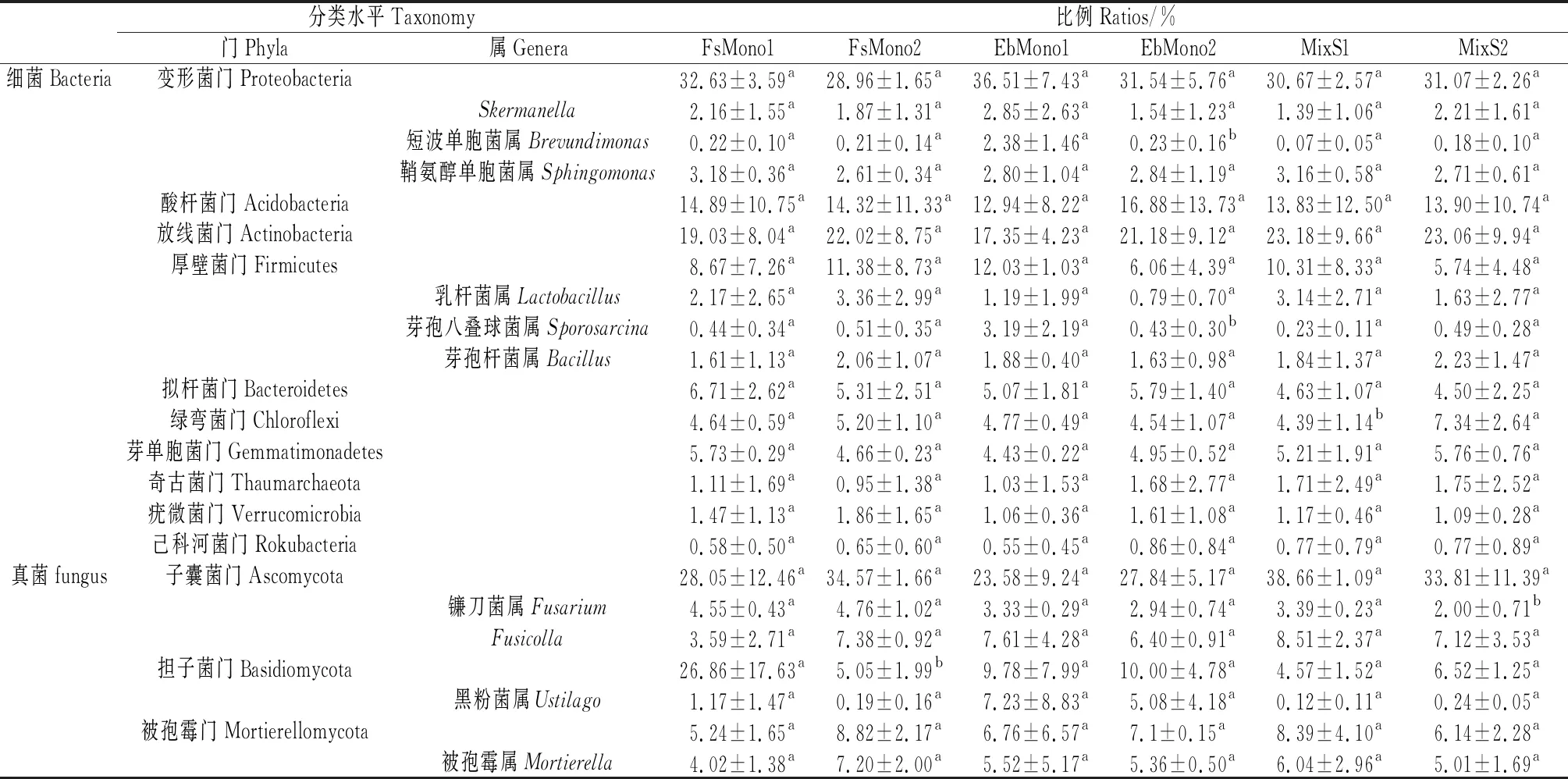
表6 不同样地土壤微生物在门和属水平上的丰度变化
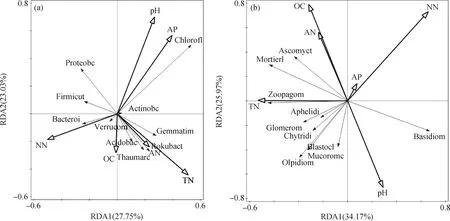
图2 土壤微生物群落与土壤化学性质的冗余分析
丰度排名前35的细菌属与环境因子之间的Spearman分析表明(图3),鞘氨醇单胞菌属(Sphingomonas)与土壤总氮含量呈极显著负相关关系(P<0.01),Haliangium与土壤总氮呈显著负相关关系(P<0.05);Acidibacter与土壤铵态氮含量呈显著负相关关系(P<0.05),慢生根瘤菌属(Bradyrhizobium)与土壤硝态氮含量呈显著正相关关系(P<0.05)。
丰度排名前35的真菌属与环境因子之间的Spearman分析结果显示(图4),被孢霉属(Mortierella)和细基格孢属(Ulocladium)与土壤pH呈显著负相关关系(P<0.05);假雨盘菌属(Pseudombrophila)与土壤pH呈极显著负相关关系(P<0.01);假丝酵母属(Candida)和Kernia与土壤pH分别呈极显著正相关关系(P<0.01)和显著正相关关系(P<0.05)。黑粉菌属(Ustilago)与有机碳含量呈显著负相关关系(P<0.05);Fusicolla和Neonectria与土壤有机碳含量均呈显著负相关关系(P<0.05);Dactylonectria与土壤有机碳含量呈极显著正相关(P<0.01)。Pseudogymnoascus与土壤总氮含量呈显著正相关关系(P<0.05);Clonostachys与土壤总氮含量呈显著正相关(P<0.05);麦角菌属(Claviceps)与土壤总氮含量呈极显著正相关(P<0.01);细基格孢属与总氮含量呈极显著负相关关系(P<0.01);踝节菌属(Talaromyces)与土壤总氮含量呈显著正相关关系(P<0.05)。镰刀菌属和附球霉属(Epicoccum)与土壤铵态氮含量均呈显著负相关关系(P<0.05);Dactylonectria与土壤铵态氮含量呈显著正相关关系(P<0.05);附球霉属(Epicoccum)与土壤铵态氮含量呈显著负相关关系(P<0.05)。放射毛霉属(Actinomucor)与土壤硝态氮含量呈极显著正相关关系(P<0.01);Pseudogymnoascus与土壤硝态氮含量呈极显著负相关关系;Clonostachys与土壤硝态氮含量呈显著负相关关系(P<0.05);麦角菌属与土壤硝态氮含量呈极显著负相关关系(P<0.01);细基格孢属与土壤硝态氮含量极显著正相关关系(P<0.01);假丝酵母属与土壤硝态氮含量呈显著负相关关系(P<0.05);Boeremia与土壤硝态氮含量呈显著正相关关系(P<0.05);Kernia和油壶菌属(Olpidium)与土壤硝态氮含量均呈显著负相关关系(P<0.05)。Solicoccozyma与土壤速效磷呈显著负相关关系(P<0.05)。微生物属水平Spearman相关性分析结果显示,环境因子对真菌的影响比对细菌更为显著。
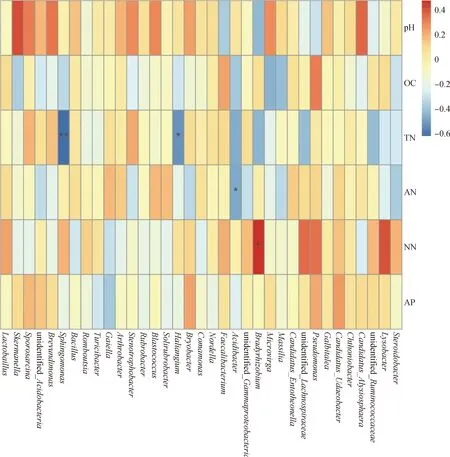
图3 土壤细菌属水平Spearman分析
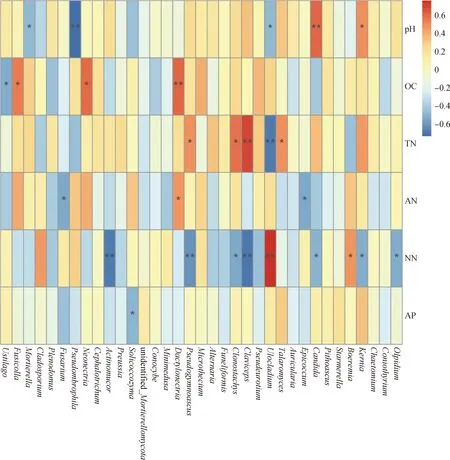
图4 土壤真菌属水平Spearman分析
3 讨论
3.1 不同种植方式和年限土壤微生物群落变化
土壤微生物是土壤有机质的主要分解者,也是土壤养分的重要源和汇,受土壤环境影响[16]。本研究检测出的细菌OTU数以及种类均高于真菌的,表明高寒地区多年生人工草地土壤微生物类群以细菌为主,这与杨希智[17]、韩玉竹[18]和胡雷[19]等有关高寒草地土壤微生物的研究结果一致。单播草地和混播草地的细菌OTU数均随生长年限增长而减少,短芒披碱草和混播群落土壤真菌OTU数呈现增加趋势,这与马欣[20]的结果一致,随着生长年限的增长,土壤中细菌生长环境变差,细菌生长受阻从而促进了真菌生长[12]。短芒披碱草单播草地的土壤真菌OUT数明显少于中华羊茅和混播草地的,这与牧草种类以及生长年限[12]有关。
Shannon指数可以用来估算样品中微生物的多样性[21],Chao1指数表示土壤微生物的丰富度[22],本研究不同种植方式土壤细菌多样性和丰富度均没有显著差异,表明在该地区无论是单播还是混播多年生禾草对土壤细菌群落没有显著影响。中华羊茅单播草地土壤细菌丰富度随生长年限的增长逐渐降低,这与周宝利等[12]关于土壤细菌变化的研究结果相似,与土壤肥力衰退有关。短芒披碱草单播草地和混播草地细菌多样性和丰富度均在年限上未发现显著变化,可能与本研究的年限较短有关。在检测到的63个细菌门中,变形菌门是土壤中丰度最高的细菌类群,这一结果与其他学者关于草地土壤细菌多样性的研究一致[23]。研究发现短芒披碱草单播草地短波单胞菌占比随年限减小,混播草地土壤绿弯菌门占比随年限显著增加,这与耿德洲等[24]和赵帆等[25]的研究结果一致,可能与土壤中速效磷含量变化相关。
真菌多样性和丰富度分析结果显示,混播草地显著高于短芒披碱草草地,这与植物群落多样性对土壤微生物的影响结果相同,植物群落多样性增加可以提高土壤微生物多样性[26-27]。中华羊茅单播草地真菌多样性和丰富度均显著高于短芒披碱草草地的,但生长年限对土壤真菌多样性和丰富度无显著影响,也可能是本研究研究的生长年限短,土壤真菌尚未发生显著变化。高雪峰等[23]和伍文宪等[8]分别在短花针茅(Stipabreviflora)荒漠草原和川西北高寒牧区不同人工草地土壤微生物群落研究中发现子囊菌门、担子菌门是优势真菌类群。本研究中,6个样地的土壤优势真菌门按照丰富度由高到低排列顺序依次为子囊菌门>担子菌门>被孢霉门。子囊菌门与担子菌门大多数为腐生菌,本研究的土壤pH均在8.5~9.0,适合腐生真菌的生长,这可能是子囊菌门与担子菌门为优势菌门的一个原因[28],本研究发现被孢霉门也是较优势的类群,这和耿德洲等[24]研究结果一致,被孢霉门真菌与土壤有机碳、全氮和铵态氮含量有关,是土壤养分含量丰富标志类群。本研究表明混播草地子囊菌门占比显著高于短芒披碱草单播草地,与真菌多样性和丰富度变化一致。纳小凡等[29]研究显示随着生长年限的增长,担子菌门的比例显著下降,本研究中华羊茅单播草地土壤担子菌门的占比也随年限减小,发现与土壤硝态氮和速效磷的含量有关。
3.2 土壤微生物群落与土壤理化性质
土壤微生物数量和多样性的大小也是表征土壤肥力状况的重要生物学指标,通常情况下,土壤养分与微生物生物量呈正相关[30]。薛凯等[31]关于青藏高原高寒草地土壤微生物群落及影响因子的研究发现,有机碳、铵态氮和速效磷与土壤微生物的多样性有显著的相关性,杨希智等[15]和齐文娟等[32]的研究也证实了总氮与速效磷对土壤微生物影响显著,本研究发现中华羊茅和短芒披碱草单播草地硝态氮和速效磷含量随着生长年限的增长呈降低趋势,所以单播草地土壤微生物种类数量和结构的变化主要由硝态氮和速效磷的变化引起。在已有的报道中,变形菌门被认为与碳利用有关,是世界上最为普遍的菌门[5],本研究发现变形菌门与土壤有机碳、铵态氮含量呈负相关。相关研究还发现酸杆菌门与土壤营养有较大相关性[33],本试验也证实了这一点。有研究表明人工草地建植初期,群落处于物种组成简单,生产力高且较稳定的阶段,加之施肥等管理措施,微生物群落结构较简单,绿弯菌门相对较少,随着生长年限的增长,绿弯菌门逐渐增加[18],本研究也发现这一现象,并发现绿弯菌门的丰富度和速效磷有关。土壤养分的变化影响到土壤真菌的变化,中华羊茅草地土壤担子菌门与硝态氮和速效磷含量变化一致。研究发现土壤化学性质对细菌菌属影响较小,对真菌菌属影响较大,镰刀菌属随生长年限的增长逐渐降低,但与土壤铵态氮含量的变化趋势不一致,这可能受土壤环境中其他因素影响,但是具体原因有待进一步的研究探讨。
4 结论
在所研究的高寒地区不同生长年限的单播和混播人工草地土壤中共识别出63个细菌门类、17个真菌门类,约有50%的真菌门未识别出来。已识别类群中细菌以变形菌门、酸杆菌门和放线菌门占优势,真菌以子囊菌门、担子菌门和被孢霉门占优势。短时间内,不同种植方式下土壤细菌多样性和丰富度无显著变化;中华羊茅草地和混播草地土壤真菌多样性和丰富度显著高于短芒披碱草单播草地的;同一种植方式混播草地不同生长年限细菌和真菌无显著变化,单播草地土壤细菌随年限的增长逐渐减少。高寒地区多年生人工草地土壤微生物多样性和丰富度显著受土壤硝态氮和速效磷含量变化的影响。
