摘花和打顶措施对毛竹林下多花黄精块茎生物量积累特征的影响
2021-04-07杨清平陈双林郭子武
杨清平,陈双林,郭子武*,郑 进
(1.中国林业科学研究院亚热带林业研究所,浙江 杭州 311400;2.江山市林业技术推广站,浙江 江山 324100)
我国毛竹(Phyllostachysedulis)林面积超过400万hm2 [1],分布于20多个省(市、自治区),丰富的毛竹林资源、多样的立地生境与气候条件为毛竹复合经营提供了环境条件和生物基础,而复合经营也为目前毛竹林效益下滑、生物多样性降低和生态功能退化等问题提供了有效解决途径[2-3]。林药复合经营是近年来兴起的竹林复合经营新模式,已经开展了诸如毛竹-多花黄精(Polygonatumcyrtonema)、金线莲(Herbaanoectochili)、白芨(Bletillastriata)、白术(Atractylodesmacrocephala)、决明(Cassiaobtusifolia)等复合经营技术的研究与探索[4-8]。其中,毛竹与药用植物复合经营多通过林地土壤管理、林下植被控制与林分结构调控等措施来改善林地生境条件[6,8-12],达到增产增效的目的,而通过生长促进措施,促进药用植物良好生长与效益增加的研究较少[13-14],毛竹林下药用植物效益体现不显著,一定程度上限制了竹药复合经营新技术的大规模推广应用。
多花黄精为百合科(Liliacea)黄精属(Polygonatum)多年生草本植物,分布区域广,适应性较强,是毛竹林下常见植物之一。多花黄精块茎是中国传统大宗药材,已成为多种增强免疫类药品、保健品的重要原料,同时还可加工成蜜饯。鉴于其良好的市场开发应用前景和在毛竹林下良好的适应性,一些毛竹主产区开展了毛竹林下多花黄精复合经营技术研究与推广应用,探明了毛竹林林分结构、坡位、坡度等对多花黄精生长和活性成分含量的影响[5-6,9,13-14],然而摘花、打顶等措施对多花黄精块茎生长与生物量分配的影响鲜见报道。本研究以毛竹林下人工种植3年的多花黄精为研究对象,通过摘花、打顶、摘花并打顶处理后多花黄精1年生、多年生块茎与根系生物量及其异速生长关系的变化规律,分析摘花、打顶措施对多花黄精块茎生物积累与分配的影响,阐明摘花、打顶处理的增产增效作用,以期为毛竹-多花黄精复合经营增产增效技术集成与创新提供理论指导。
1 材料与方法
1.1 试验区概况
试验区地处浙江省江山市保安乡(118°22′37″~118°48′48″E,28°15′26″~28°53′27″N),年平均气温17.1 ℃,年降水量1 650~2 200 mm,年平均日照时长2 063.3 h,年平均无霜期253 d左右。试验区土壤为黄红壤,土层厚度60 cm以上。区域内毛竹资源丰富,面积约2 300hm2,多为纯林,以材用林经营为主,经营相对粗放,实行季节性伐竹、留笋养竹,没有化学除草剂使用历史,未采取过林地垦复和施肥等措施。为增加毛竹林经济效益,2011年4月在试验区选择水肥条件好、土壤疏松的区块状毛竹林进行多花黄精复合种植,种苗栽植初植密度3万株/hm2左右,成活率95%以上,种植面积20.8 hm2,长势良好。
1.2 试验处理与调查方法
2013年4月初(盛花期),选择坡度15°以下半阴坡的毛竹-多花黄精复合经营试验林3块,每块面积不小于1.2 hm2,在每块试验林中布设20 m×20 m样地3个,调查毛竹林结构和多花黄精地上生长情况。试验毛竹林林分结构为:立竹密度(2 700±165) 株/hm2,立竹胸径(9.22±1.01) cm,年龄结构1度、2度、3度竹株数比为3.74∶4.17∶1.09。多花黄精植株高(65.8±4.78)cm,地径(4.19±0.32) mm。每块样地中随机选取生长正常的多花黄精样株各90株,分别实施打顶(DD,去除冠层高度的1/3)处理、摘花(ZH,摘去处理植株所有的花)及摘花打顶处理(ZD),每处理各30株,另选30株为对照。于2013年10月多花黄精地上部分枯萎后,将所有样株连根完整挖出,去除泥土等杂物,装于冰盒中带回实验室,冲洗干净泥土后,用天平(0.01 g)称量多年生整个块茎鲜质量(g)、1年生块茎鲜质量(g)和根系鲜质量(g),并计算地下部分总生物量及各构件生物量分配比例。
1.3 数据处理与分析
试验数据在SPSS 19.0统计软件中进行单因素方差分析(One-Way ANOVA)。多花黄精地下构件生物量异速生长用方程Y=a×xb来描述,经对数转换方程形式为lgY=a+b(lgx),其中x与Y代表各处理地下总生物量和构件(1年生块茎、多年生块茎、根)生物量,方程斜率b为异速生长指数。采用标准化主轴回归分析(standardized major axis,SMA)计算方程斜率b,并比较斜率之间及其与1.0的差异性。若斜率间无显著差异,则给出共同斜率,并采用Wald检验各处理沿共同主轴位移差异的显著性,并计算位移量。SMA分析采用R语言的Smatr模块完成[15-16]。
2 结果与分析

不同小写字母示处理间差异显著(P<0.05),不同大写字母示器官间差异显著(P<0.05)。下同。Different lowercase letters indicated significant differences among the treatments at 0.05 level,and different capital letters indicated significant differences among the organs at 0.05 level.The same below.图1 不同处理多花黄精地下器官生物量积累Fig.1 Biomass of belowground organ of Polygonatum cyrtonema under different treatments
2.1 摘花和打顶措施对毛竹林下多花黄精地下构件生物量积累的影响
不同经营措施下,毛竹林下多花黄精地下各器官生物量差异明显(图1),多年生块茎生物量显著高于1年生块茎和根系生物量。摘花和打顶措施对多花黄精块茎生物量积累影响显著,多年生块茎生物量较对照显著增加,但摘花、打顶及摘花打顶处理间并无显著差异。摘花、打顶及摘花打顶后,多花黄精1年生块茎、根及块茎总生物量显著增加;摘花处理多花黄精1年生块茎、根及总生物量虽高于打顶处理,但二者并无显著差异,且均显著低于摘花打顶处理,即摘花打顶措施对多花黄精地下生物量积累作用显著,呈现明显的加性效应(图 1)。
2.2 摘花和打顶措施对毛竹林下多花黄精地下构件生物量分配的影响
多花黄精地下构件生物量分配比例差异明显,其中以多年生块茎生物量分配比例最高,超过50%,其次是1年生块茎生物量比例,根系生物量比例最低,不足20%。摘花和打顶措施对毛竹林下多花黄精地下构件生物量分配会产生明显的影响。与对照相比,摘花、打顶和摘花打顶处理后多花黄精多年生块茎生物量比例均显著下降,摘花处理略低于打顶处理,二者并无显著差异,且二者均显著高于摘花打顶处理;摘花、打顶和摘花打顶处理会使多花黄精1年生块茎生物量分配比例显著提高,摘花、打顶处理间并无显著差异,且均显著低于摘花打顶处理;各处理的根系生物量分配比例并无显著差异(图 2)。
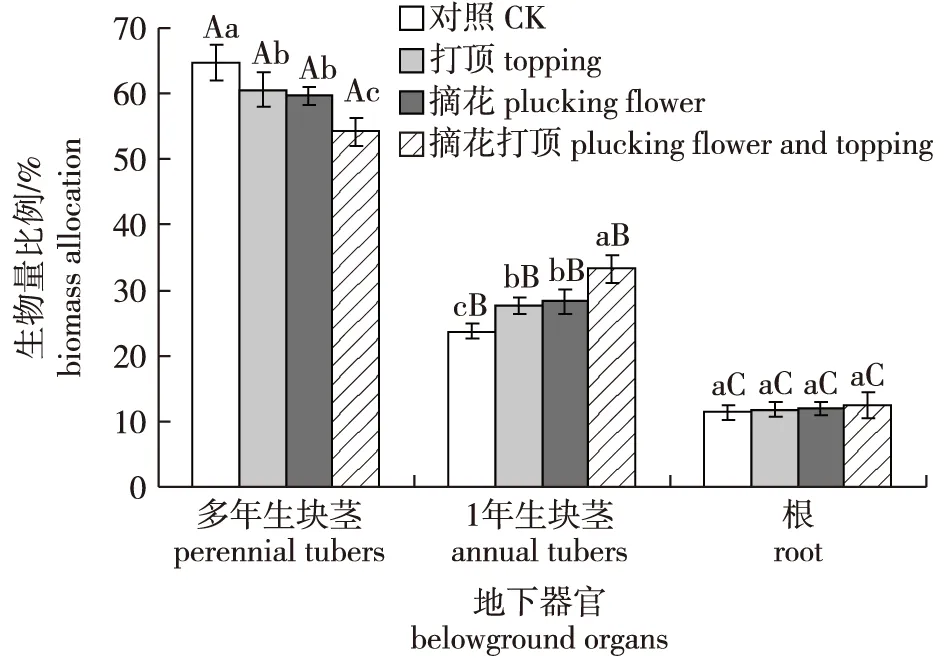
图2 不同处理多花黄精地下器官生物量分配Fig.2 The biomass allocation of belowground organ of Polygonatum cyrtonema under different treatments
2.3 摘花和打顶措施对毛竹林下多花黄精地下构件生物量异速生长关系的影响
多花黄精地下构件多年生块茎、1年生块茎、根与总生物量间均呈异速生长关系(b≠1)(图3,数据均经对数转换)。摘花、打顶、摘花打顶处理的多花黄精多年生块茎生物量与总生物量间异速生长指数总体呈升高趋势,但各处理异速生长指数并无显著差异,且存在共同斜率(0.875),均沿共同斜率出现了显著的正向移动(移动量分别为0.012 0、0.025 1和0.050 3,Wald检验,P<0.05)。摘花、打顶、摘花打顶处理后多花黄精1年生块茎、根生物量与总生物量异速生长指数总体上均明显增加,1年生块茎生物量与总生物量间异速生长指数各处理间差异显著,以摘花打顶处理异速生长指数最大(0.871);摘花、打顶根与总生物量异速生长指数仅略高于对照,三者间存在共同斜率(0.185),且以摘花打顶处理最高(图3b、3c)。这说明摘花和打顶措施明显促进了1年生块茎和根的生长,且以摘花打顶处理增效最佳。

图中数据均经对数转换。The data in the figues is logarithm with base 10.图3 不同处理多花黄精地下构件异速生长关系Fig.3 The allometric growth of belowground module biomass for Polygonatum cyrtonema
3 讨 论
农业生产上往往通过去除作物部分器官生物量及其比例的方法来调控作物生长极性与营养生长和生殖生长进程,从而达到增产增效的目的[17-20]。合理打顶能打破植物的顶端优势,促进植物更好地利用光热资源并获取更多养分,最大限度地提高作物产量和改善品质;而摘除部分花果则打破了营养生长与生殖生长的平衡,改变了营养物质分配方向与模式,从而在一定程度上促进营养的储存与产量的提高[21-23]。本研究表明,摘花和打顶措施对多花黄精块茎生物量积累与分配产生显著影响。打顶、摘花及摘花打顶后多花黄精块茎生物量较对照明显增加,虽然打顶、摘花处理间多花黄精多年生、1年生块茎及块茎总生物量并无显著差异,但打顶摘花处理的1年生块茎及块茎总生物量显著高于打顶、摘花处理,这与1年生块茎生物量分配比例显著增加的研究结果一致,说明摘花和打顶措施显著促进多花黄精块茎的生长,并且以打顶摘花处理块茎的增效最明显,体现出处理措施的加性效应。这可能是因为合理打顶、摘花及打顶摘花处理的生物量去除及伤口诱导作用,引起了多花黄精强烈的补偿性生长[23-25],拉动了多花黄精光合碳同化,促进了多花黄精1年生块茎生物量的积累。但打顶不仅破除了多花黄精顶端优势,也明显减少了植株叶片数量,会导致光合碳同化能力短时间内受抑,其增产增效作用受到了部分限制[20,23]。而摘去部分多花黄精的花,显著降低花果对碳同化物的拉力,块茎对碳同化物的拉力骤增,势必促进碳同化物源源不断地流向块茎[26-28]。另一方面,地下块茎和种子均是多花黄精重要的繁殖体,而摘除部分花果后,多花黄精种子数量骤减,迫于生存与种群延续的压力,多花黄精势必会动员大量光合碳同化产物向地下块茎运输[29],以提高块茎的生长量,以期增加繁殖体数量,达到维持种群繁衍与稳定的目的,这也可能是摘花增产效果强于打顶的原因之一。
植物器官的生物量积累规律及其异速增长模式虽主要由其生物学特性和遗传特征决定,但环境条件、经营措施和人工干扰也会对其产生重要的影响[20,30-32]。打顶、摘花及摘花打顶等措施,疏除了多花黄精植株部分生物量,势必导致其构件剩余部分相对生长关系、生物量积累与分配格局的改变。本研究结果显示,打顶、摘花处理后,多花黄精多年生块茎生物量与总生物量间异速生长指数仅有升高,各处理间存共同斜率,但却沿共同主轴产生了正向移动[25]。这可能是由于多年生块茎的生物量增加往往体现在干物质的积累与含水率的下降上,其膨大生长较缓慢,导致增效作用较小;而由于相对位置的优势,1年生块茎往往处于优先生长的地位,加之其对碳同化物的截流作用,从而在一定程度上导致运输到多年生块茎碳同化物量的递减,这也可能是多年生块茎生长能力与活性下降的原因之一。与多年生块茎相比,摘花和打顶措施使1年生块茎异速增长指数显著增加,各处理增幅均超过12%,且以摘花打顶处理增速最大(67.31%),这说明1年生块茎由于着生的位置优势和强烈的生长活性,生物量增速最快,对摘花和打顶措施最为敏感。
毛竹林下药用植物复合经营的关键技术是适生和经济价值高的药(食)用植物筛选、生境确立与优化及增产增效措施的研发。本研究表明:打顶、摘花及摘花打顶措施显著增加了多花黄精块茎和根系生物量,改变了生物量积累与分配模式和相对生长关系,且各器官对打顶、摘花及摘花打顶处理的敏感性不同,以1年生块茎对摘花和打顶措施最为敏感,因而呈现很好的增产作用;摘花和打顶措施增产效果也存在差异,其中以摘花打顶处理的块茎增产效果最好,可作为毛竹林下多花黄精复合经营的增产增效推荐技术。
本试验仅研究了打顶、摘花及摘花打顶处理1年对多花黄精块茎生长的影响,而多花黄精一般是种植后3年采收块茎,因此仍需开展连续处理对多黄精块茎增长与生物量分配影响的研究。同时,摘花和打顶措施虽然增加了多花黄精块茎生物量,但是否也会影响块茎有效成分的含量,从而呈现一定的药用价值增效作用,也是下一步研究重点。
