中国杨树上的栅锈菌
2021-04-07梁英梅田呈明
姜 宁,梁英梅,田呈明
(北京林业大学,北京 100083)
锈菌是一类专性寄生真菌,引起多种植物的锈病。栅锈菌属(MelampsoraCastagne,1843)是人类不得不重视的病原菌,其拉丁学名含义为黑色坏死斑,中文名取自其冬孢子栅栏状的排列方式。在已知的百余种栅锈菌中,多数为害杨柳科(Salicaceae)寄主,导致明显的叶面失绿和枯死;少数寄生在亚麻科、夹竹桃科和大戟科等植物上[1-2]。
杨树(Populus)是我国防护林、用材林和城市绿化等人工林的主要造林树种之一,分布范围极广。叶锈病作为杨树上普遍而严重的叶部病害,常见于世界各杨树栽培区,在美洲国家感病杨树品种的人工林中,材积损失可高达65%;美国中部的杂交杨,由于锈病危害,减少材积31%~42%[3]。大量的栅锈菌寄生于杨树叶部,影响光合作用,可导致树势衰弱,诱发次期性病虫害的侵害而死亡,也对天然林的更新造成严重的影响[4-10],为此B.A.Thielges[3]将其列为杨树5大严重病害之首。
在Castagne建立栅锈菌属之后,Van Kraayenoord对分布于世界杨树上的32种栅锈菌进行系统整理,仅认可其中的11种[11]。尚衍重等[12]研究了已报道的杨树上34种栅锈菌,并根据寄主范围和夏、冬孢子形态特征的不同而承认其中的12种。袁毅[13]首次较为系统地研究了我国杨树上的栅锈菌并记载包括新记录种在内的7种栅锈菌。戴玉成[14]对杨树上的栅锈菌通过数量分类学方法,讨论了栅锈菌之间的进化关系,承认杨树栅锈菌有14种。Leveille依据寄主类型将杨树上的种都定为Melampsorapopulina,与锈菌的生物学特性有所结合,在一定程度上是一种进步,后来Thumen[1]和Harting[2]也曾根据寄主种类发表并订正了一些种,但并不符合实际。G.Bagyanarayana[15]根据夏孢子的大小、侧壁的加厚程度、冬孢子的大小及顶部是否加厚等特征,将杨树上的栅锈菌分为9个种,并根据转主寄主(锈孢子寄主)将M.populnea分为5个专化型。但这种划分未得到广泛认可[2]。目前仍然有很多学者包括我国的一些学者也使用该名称确定杨树上的一些种类,将青杨派、白杨派上的很多栅锈菌都鉴定为M.populnea,但我们研究后发现仅寄生于山杨上的M.laricis(同物异名)属于该种。迄今为止在栅锈菌属中,寄生在杨树上的栅锈菌有14个种和2个杂交种被广泛承认[2,16]。
目前国内外学者主要根据冬孢子形态及其在叶面的着生部位、夏孢子的形状、大小、壁的厚度、表面刺突的形状与分布、芽孔的数目、侧丝以及转主寄主植物的种类等作为分种的主要依据。然而由于栅锈菌的许多种在症状及病原形态等方面很相似,不同学者利用不同的分类标准在栅锈属的种类划分上得出了不同的结论。造成栅锈菌分种混乱的主要原因有:多数栅锈菌主要以夏孢子侵染杨柳科植物叶片或茎,有时不产生冬孢子,况且不同种之间的冬孢子形态差异并不明显;有些杨树栅锈菌不经过转主而完成其生活史;栅锈菌的寄生专化性不是很强,一个种可侵染不同派、不同种或品种的杨树,而同一种杨树又会被几种栅锈菌侵染,这样寄主杨树的种类在分类学上只能作为辅助特征;多数转主寄生种,人为强制接种可使非寄主植物发病。因此有必要建立较为完善的分类体系,明确杨树上的栅锈菌种类。本文系统地整理了中国报道的杨树栅锈菌类群,依据形态学特征和系统学分别进行了种类界定,最终形态学与系统学互为验证,确定中国杨树上的栅锈菌类群为8种。
1 中国杨树栅锈菌
1.1 冷杉-杨栅锈菌
Melampsoraabietis-populiImai,in Ito et Murayama,Trans.Sapporo Nat.Hist.Soc.17:164,1943
=Caeomaabietis-mayrianaeS.Imai,1942
性孢子器表皮下生,Ⅲ型。锈孢子器叶背生;锈孢子球形、近球形、倒卵形至椭球形(20~32.5×17.5~27)μm,壁厚2~3.5 μm,密生疣。夏孢子堆叶背散生,淡黄色,0.1~0.5 mm;夏孢子球形、椭圆形至长椭圆形,(17~28×9~24)μm,壁均匀加厚,0.8~2.7 μm,具刺,刺间距0.9~2.4 μm;侧丝头状,(30~65×11~30)μm,顶部加厚至7.5 μm(图1A、图1B)。冬孢子堆叶背单生或聚生,多边形,红褐色,0.3~1 mm;冬孢子菱形、棍棒状至圆柱状,(21~45×5~11)μm,壁厚1 μm。
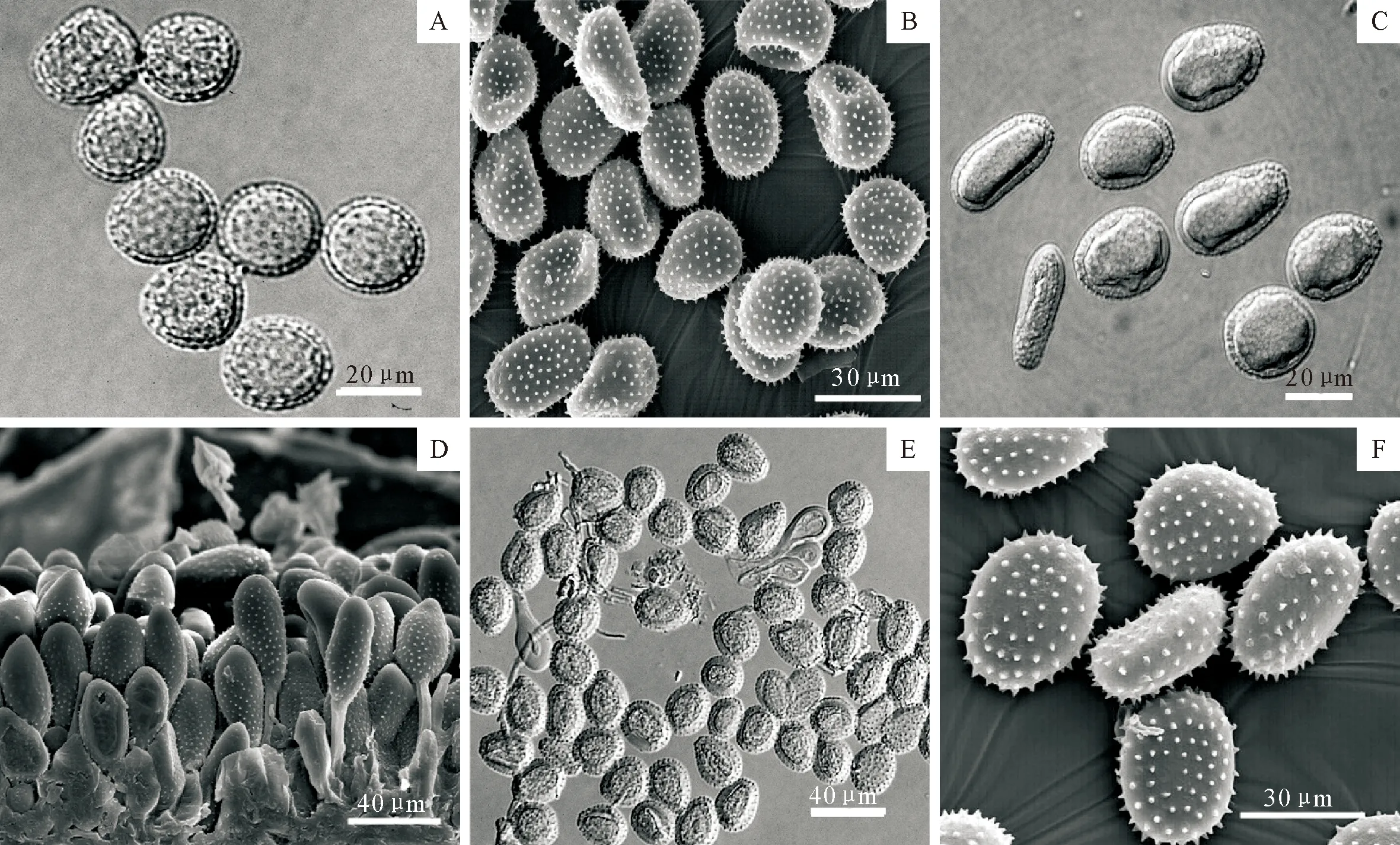
注:A,B.Melampsora abietis-populi冷杉-杨栅锈菌; C,D.M.allii-populina葱-杨栅锈菌; E,F.M.populnea杨栅锈菌。
寄主:0,Ⅰ寄生在日本冷杉Abiesfirma,日光冷杉A.homolepis,北海道冷杉A.sachalinensisvar.mayriana,东陵冷杉A.nephrolepis,富士山冷杉A.veitchii。
Ⅱ,Ⅲ寄生在香杨Populuskoreana,辽杨P.maximowiczii,黑杨P.nigra,钻天杨P.nigravar.italica,长序杨P.pseudoglauca,藏川杨P.szechuanicavar.tibetica,小叶杨P.simonii,欧美杨P.×euramericana,椅杨P.wilsonii,滇杨P.yunnanensis。
分布:中国、日本、俄罗斯。
讨论:该种的模式标本采自钻天杨上,同时用采自东陵冷杉的锈孢子接种钻天杨获得成功。随后Hiratsuka和Karube通过向香杨、辽杨、欧美杨等树种人工接种源自冷杉上的锈孢子,也证明了该种的转主寄生关系,并发现其夏孢子不能侵染白杨派树种[17-19]。此后由于该种仅在日本发现,故一度被认为是日本特有种。20世纪90年代在我国秦岭的椅杨上首次发现[20-22],此后在滇杨上有报道[18],但我国未发现锈孢子阶段寄主,也没有接种试验的报道。
Melampsoraabietis-populi和M.magnusiana、M.rostrupii以及M.populinea等种在夏孢子和冬孢子形态上极为相似,唯一不同的是该种具有极薄的夏孢子壁,而且刺间距也明显小于其他杨树锈菌[18-19]。
很多寄生在长序杨上的栅锈菌常常被错误地鉴定为Melampsorapopulnea(Per.) Karst,然而这些标本上的夏孢子具有很薄的壁,而且它们的系统发育学分析都显示与M.abietis-populi属于一个单进化枝。因此,夏孢子壁薄与否可以作为区分这个种和其它杨树栅锈菌的形态特征[18]。
1.2 葱-杨栅锈菌
Melampsoraallii-populinaKleb.Zeit.Pfl.-Krankh.,12:25,1902
=CaeomaalliorumLink,sp.Pl.2,7,1825
=Caeomaallii-ursiniWinter,Pilze Deutschl.I,p.255,1881
=Uredoallii-populinaArth.,Rèsult Scient.Congr.Intern.Bot.Vienne p.338,1905,1906
=Melampsorapopulneaf.sp.allii-populina(Kleb.) Bagyanarayana,Proce.1th IUFRO Fore.Trees Working Party Conf.P.47,1998
性孢子器叶表皮下生,圆饼形,直径60~100 μm。锈孢子器裸露,生于叶斑上,亮橙红色,直径0.8~1.2 mm;锈孢子球形、卵形,(17~25×14~18)μm,壁厚2 μm,密生疣。夏孢子堆叶两面生,圆形突出,橘黄色,约1 mm大小;夏孢子棍棒状至长椭圆形,稀球形,(20~40×10~24)μm,壁通常均匀加厚,偶见一端偏厚,但决不在赤道部加厚,大小1.3~6.8 μm,除顶端外均具刺,刺间距1.3~3.5 μm;侧丝通常头状,具细茎,(50~71×15~23)μm,壁厚2~3 μm(图1C、图1D)。冬孢子堆叶两面生,多数生于叶背,单生或群生,红褐色,0.5~1 mm;冬孢子不规则圆柱状,(35~60×7~13)μm,壁厚1~1.5 μm,顶端偶见增厚,但不超过2 μm。
寄主:0,Ⅰ寄生葱属Allium;Ⅱ,Ⅲ 寄生山杨P.davidiana,苦杨P.laurifolia,合作杨P.opera,密叶杨P.talassica。
分布:中国新疆和内蒙,以及欧洲南部和俄罗斯。
讨论:在已报道的中国栅锈菌中,Melampsoraallii-populina和M.larici-populina在夏孢子形态特征上很相似,但是该种的夏孢子壁较均匀且不加厚,而且冬孢子叶两面生是区别M.larici-populina最具识别度的特征。Bagyanarayana曾将葱-杨栅锈菌以及其它4种杨树栅锈菌一起,根据锈孢子寄主的不同作为M.populnea的专化型处理[15]。然而,M.allii-populina夏孢子较大、头部具光滑区以及冬孢子叶两面生等特征很容易与M.populnea复合种区别开来,同时分子系统学研究结果也支持这是一个独立的种[16]。野外接种试验证明,M.allii-populina能够以菌丝状态在杨树上越冬,因而在中国北方缺乏锈孢子寄主的地区亦然可以流行。
1.3 落叶松-杨栅锈菌
Melampsoralarici-populinaKleb.,Zeit.Pfl.-Krankh.,12:43,1902
=CaeomalaricisHart.Wichtige Krankh.Wäldbaume,p.93,1847
=Uredolarici-populinaArth.,Rèsult Scient.Congr.Intern.Bot.Vienne p.338,1905,1906
=Melampsorapopulina(Jacq.) Lev.,Ann.Sci.Nat.,p.375,1847
性孢子器叶背表皮下生,Ⅲ型,球形,褐色,直径100 μm,高70 μm。锈孢子器裸露,生于叶背黄色病斑上,橘黄色,表皮下生至突出状,直径0.5~1.5 mm;锈孢子球形、卵形,(17~30×14~27)μm,壁厚1~2 μm,密生疣,疣长2 μm。夏孢子堆叶背生,稀叶正面生,表皮下生,成熟后突出寄主表皮,粉状,橘黄色,直径0.3~1 mm;夏孢子长椭圆、矩圆形至棍棒状,(20~55×11~30)μm,壁厚度均匀但在赤道处强烈加厚,平均5.1 μm,最厚处达12.4 μm,孢子基部刺较大往上刺逐渐变小,绝大多数孢子头部光滑无刺,刺间距1~5 μm;侧丝棍棒状至头状,(50~90×15~25)μm,顶部加厚至25 μm(图2A,B)。冬孢子堆多叶正面生,稀叶背生,单生或聚生,多边形,红褐色;冬孢子圆柱状或菱形,两端圆,(20~45×7~12)μm,壁厚1~1.5 μm,顶部加厚2~3 μm。
寄主:0,Ⅰ阶段寄生欧洲落叶松Larixdecidua,落叶松L.gmelinii,L.gmeliniivar.japonica,美洲落叶松L.laricina,日本落叶松L.kaempferi,西部落叶松L.occidentalis,黄花落叶松L.olgensis,华北落叶松L.principis-ruppechtii,新疆落叶松L.sibirica。
Ⅱ,Ⅲ阶段寄生响叶杨(Populusadenopoda),中东杨(P.×berolinensis),北京杨(P.×beijingensis),青杨(P.cathayana,P.charkowiensis×P.caudina),美洲黑杨(P.deltoides),念珠杨(P.deltoidessubsp.monilifera)、棱枝杨(P.deltoidessubsp.wislizenii),密苏里杨(P.deltoidesvar.missouriensis),欧美杨(P.×euramericana),甘肃杨(P.×gansuensis),香杨(P.koreana),苦杨(P.laurifolia),大叶杨(P.lasiocarpa),马里兰杨(P.×canadensis‘Mariandica’),辽杨(P.maximoniicizii),黑杨(P.nigra),钻天杨(P.nigravar.italica),箭杆杨(P.nigravar.thevestina),合作杨(P.opera),少先队杨(P.×pioner),冬瓜杨(P.purdomii),长序杨(P.pseudoglauca),小青杨(P.pseudo-simonii),小青×黑杨(P.pseudo-simonii×P.deltoides),小叶杨(P.simonii),菱叶小叶杨(P.simoniivar.rhombifolia,P.simonii×P.nigravar.italica),川杨(P.szechuanica),大青杨(P.ussuriensis),滇杨(P.yunnanensis)以及黑杨派和青杨派的许多杂交种。
分布:中国、日本、印度、韩国、蒙古、新西兰、澳大利亚、美国、俄罗斯及欧洲。
讨论:该种是栅锈菌属分布最广泛的种,也是引起中国杨树栽培区叶锈病流行最严重的病原菌。Melampsoralarici-populina依据其冬孢子堆叶正面生,夏孢子壁在赤道部强烈加厚,顶部具光滑区等特征很容易在形态上与其他杨树栅锈菌区分。袁毅[13](1984)根据夏孢子顶部具有细刺而报道了一个中国新记录种M.occidentalis,但是电子显微镜观察发现M.occidentalis的孢子表面并不具有光滑区,且该标本的分子数据也显示为M.larici-populina,因此中国目前尚未发现M.occidentalis。尚衍重等[23]基于其叶两面生的冬孢子堆和1~3层的冬孢子等特征描述了采自辽宁欧美杨的一个新种—多生栅锈菌Melampsoramulta。然而,检查该种的等模式,其形态特征与M.larici-populina的形态相似,同时从其他大量的M.larici-populina的标本中也观察到2~3层冬孢子的现象,因此认为该种很可能是落叶松-杨栅锈菌的同种。
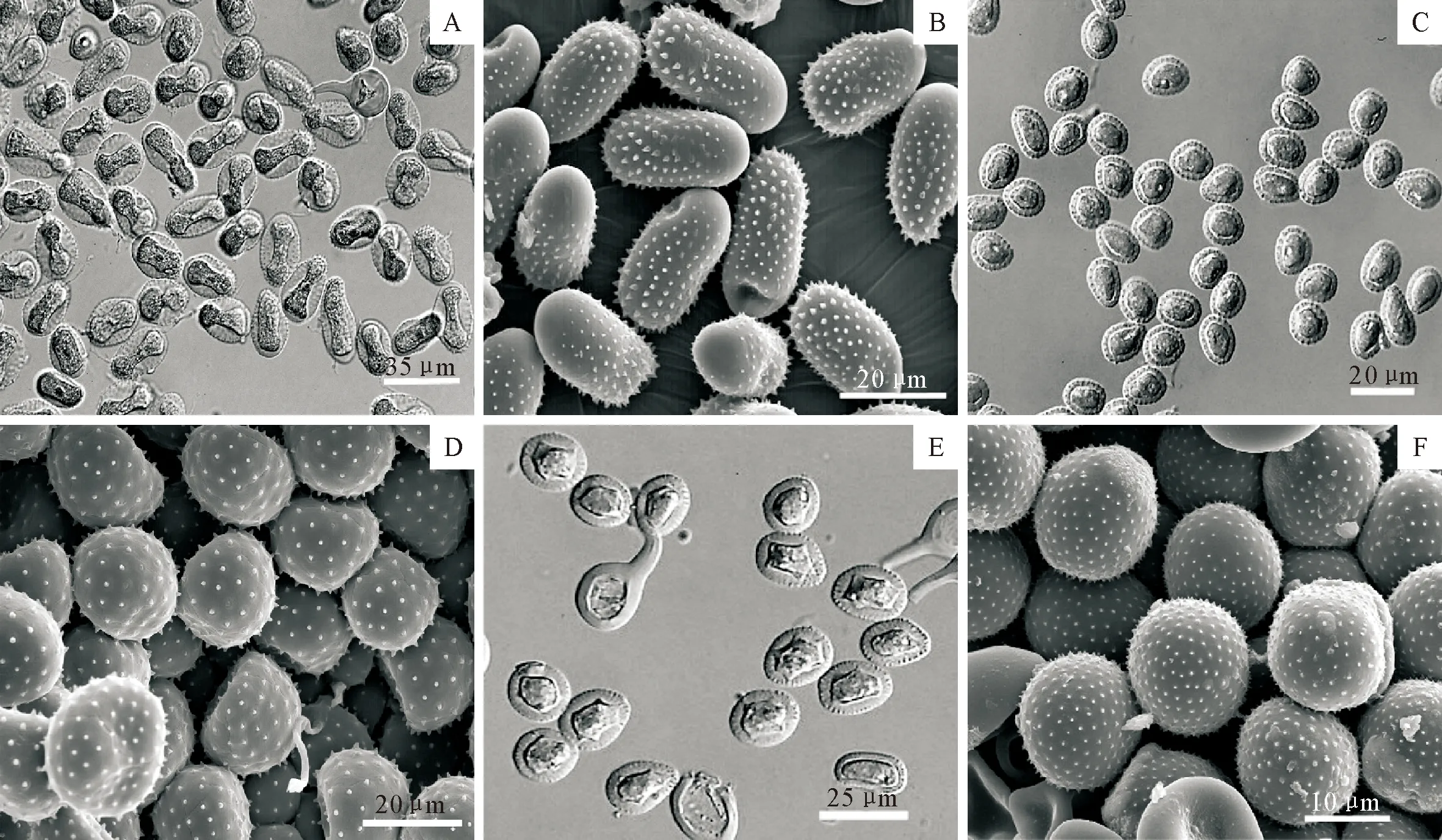
注:A,B.M.larici-populina落叶松-杨栅锈菌; C,D.M.magnusiana马格栅锈菌; E,F.M.pruinosae粉被栅锈菌。
1.4 马格栅锈菌
MelampsoramagnusianaWagner,Zeit.Pfl.-Krankh.,7:340,1897
=M.populneaf.sp.magnusianaBagyanarayana,Proce.IUFRO Rust Forest Tree Working Party Conf.,48,1998
=M.populneaf.sp.rostrupiiBagyanarayana,Proce.IUFRO Rust Forest Tree Working Party Conf.,50,1998
性孢子器叶两面生,Ⅱ型。锈孢子器叶背生;锈孢子球形、椭球形,(14~23×12~20)μm,壁厚1~1.5 μm,密生疣。夏孢子堆多数叶背、叶柄、叶芽生,稀叶正面生,表皮下生,成熟后突出寄主表皮,粉状,黄色,直径0.3~1 mm;夏孢子球形至广椭圆形,(13~33×12~26)μm,壁均匀加厚,1~5 μm,具刺,刺间距1.0~4.5 μm;侧丝棍棒状至头状,(44~95×12~24)μm(图2C,D)。冬孢子堆叶背生,圆形至多边形,红褐色;冬孢子柱状,两端圆,(37~53×7.5~12)μm,顶部加厚1~2 μm。
寄主:0,Ⅰ阶段寄生白屈菜(Chelidoniummajus),紫堇属(Corydalis);Ⅱ,Ⅲ阶段寄生银白杨(P.alba),新疆杨(P.albavar.pyramidalis),河北杨(P.hopeiensis),毛白杨(P.tomentosa),圆叶杨(P.rotundifolia),欧洲山杨(P.tremula),日本山杨(P.sieboldii)等白杨派树种。
分布:中国、美国、俄罗斯及欧洲东部。
讨论:我国的毛白杨上已经报道了3个种,即Melampsoramagnusiana[24]、M.rostrupii[25]和Uredotholopsora[26]。前2个种在形态上并没有不同,而且它们在杨树上的症状、致病性差异不大,所不同的仅仅是锈孢子寄主不同,但缺乏接种试验。一般认为M.magnusiana的锈孢子寄主是紫堇属和白屈菜属,而M.rostrupii的锈孢子寄主是山靛属Mercurialis。Miura[27]报道了采自辽宁草河口紫堇上的一种裸孢锈Caeomafumariae,但并没有确定M.magnusiana和Caeomafumariae的关系;戴芳澜[28]据此锈孢子寄主紫堇上的锈菌将其记载为M.magnusiana,但这个记载是可疑的。因此,M.magnusiana和M.rostupii的转主寄主在中国是不清楚的,而Hiratsuka通过日本山杨上的夏孢子(M.magnusiana)接种Chelidoniummajus,确定了其寄生关系[29-30]。
M.magnusiana和M.rostupii常常寄生同一个寄主杨树,所不同的仅仅是锈孢子寄主植物种类。Boerema和Verhoeven(1951)将这2个种作为M.populnea种下的小种(专化型),并认为M.rostupii是M.aecidioides的裸名[31],此后很多人都在同时使用这几个种名。尚衍重(1984)提出M.magnusiana和M.rostrupii可作为M.aecidioides的同种异名。基于同样的考虑,Bagyanarayana将杨树上的5个种作为M.populnea种下专化型处理,其中包括2个新的专化型:M.populneaf.sp.magnusiana和M.populneaf.sp.rostupii,同时指出M.populnea的夏孢子超过30 μm[15]。然而,寄生在白杨上的M.magnusiana或者M.rostupii的夏孢子均<30 μm[18]。人工接种试验也证明中国白杨上的M.magnusiana或M.rostupii是同一个种,所不同的仅仅是致病性有差异[32]。另外,对收集自欧洲、中国、日本等地区被鉴定为M.rostupii和M.magnusiana的标本的分子系统学分析结果也显示这2个种之间的遗传特性并没有差异[18],而且M.rostupii本身就是个裸名,因此在中国被鉴定为M.rostupii、M.aecidioides以及一些M.populnea的种类都应为M.magnusiana。
1.5 美洲栅锈菌
MelampsoramedusaeThüm.,Bull.Torrey bot.Club 6:216,1878
夏孢子堆主要叶背生,圆形,金黄色或橘黄色;夏孢子倒卵形至卵形,(20.5~31.5×14.0~20.5)μm,壁略加厚,赤道部具光滑区,1.7~3.6 μm,具刺;侧丝球形,(25~44×12~18)μm,壁厚4~5.5 μm。冬孢子堆主要叶背生,散生,初时淡褐色,后深红褐色至黑色;冬孢子角形至圆柱状,(31~44×10~15)μm。
寄主:0,I阶段未见。在原产地北美,寄生于美洲落叶松(Larixlaricina)和花旗松(Pseudotsugamenziesii);Ⅱ,Ⅲ阶段寄生小叶杨(P.simonii),川杨(P.szechuanica),滇杨(P.yunnanensis)。
分布:原产北美,后传播至中国、日本、俄罗斯、澳大利亚、南非、法国、印度、西班牙、葡萄牙和阿根廷。
讨论:该菌作为我国进境植物检疫对象中的危险性植物病原菌2019年首次在中国发现[33],应引起足够重视。夏孢子赤道部具有光滑区是该种区别于其它栅锈菌的主要特征。
1.6 怒江栅锈菌
MelampsoranujiangensisY.M.Liang,C.M.Tian & Kakish.,Mycoscience 47(4):199,2006
0,Ⅰ阶段未知。
夏孢子堆叶背生,多散生、少聚生,圆形至椭圆形,橘黄色或淡黄色,直径0.2~0.8 mm;夏孢子球形至卵形,(17~27×18~23)μm,壁均匀加厚,1~2 μm,具刺,刺间距1~2 μm;侧丝头状,(33~56×12~15)μm,壁厚2~6 μm。冬孢子堆叶背表皮下生,散生,黄色至红褐色,0.4~0.8 mm;冬孢子菱形、棍棒状至圆柱状,(23~44×5~13)μm,壁厚1 μm。
寄主:滇杨(P.yunnanensis);0,Ⅰ阶段未知。
分布:中国。
讨论:怒江栅锈菌与马格栅锈菌的夏孢子有部分相似,但该种的夏孢子更圆,且壁薄、刺间距更短[34],可明显与M.abietis-populi、M.magnusiana区分。
1.7 杨栅锈菌
Melampsorapopulnea(Pers.) P.Karst.,Bidr.Känn.Finl.Nat.Folk 31:53 (1878)
=Caeomamercurialis(H.Mart.) Link,in Willdenow,Sp.pl.,Edn 4 6(2):35 (1825)
=MelampsoralaricisHartig,Allg.Forest-Jagdzeit.,61:326,1885
=Melampsoraaecidioides(DC.) J.Schröt.,in Cohn,Krypt.-Fl.Schlesien (Breslau) 3.1(17-24):362 (1887)
=MelampsorapinitorquaRostr.,Farl.Snylt.Danm.Skove (Copenhagen):10 (1889)
=Melampsorapopulina(Jacq.) Lév.,Annls Sci.Nat.,Bot.,sér.3 8:375 (1847)
=MelampsorarostrupiiWagner,Oest.Bot.Zeit.,46:274,1896 (nomen nudum)
=MelampsoratremulaeTul.,Annls Sci.Nat.,Bot.,sér.4 2:95 (1854)
=M.larici-tremulaeKleb.,Forestl.Nat.Zeits,468,1897
=M.tremulaef.laricisHartig,Lehrb.Baumkrankh.Ed.2,p.14,1889
=M.populneaf.sp.laricis(Hart.).Boer.& Verh.,Neth.J.Pl.Path.,78 (1):82-201,1972
=UredolaricisArth.Result.Sci.Congr.Internat.Bot.Vienne (1905),p 338,1906
性孢子器叶两面生。锈孢子器通常叶背生;锈孢子球形、倒卵形,(14~7×12~16)μm,壁厚1 μm,密生细疣。夏孢子堆叶背表皮下生,分散,成熟后突出寄主表皮,粉状,淡黄色或土黄色,直径0.2~0.4 mm;夏孢子倒卵形至椭圆形,(14~33×12~26)μm,壁略在侧面略微加厚,1.1~5.0 μm(平均2.8 mm),具刺,赤道处偶见光滑区,刺间距1.1~4.4 μm;侧丝头状至纺锤状,(34~50×10~17)μm(图1 E,F)。冬孢子堆叶背生,黑褐色,圆形至多边形;冬孢子棱柱状,两端圆,(28~59×8~13)μm,壁厚1~2 μm,顶端不增厚。
寄主:0,Ⅰ阶段寄生黄花落叶松(L.olgensis),落叶松(L.gmelini),日本落叶松(L.kaempferi),华北落叶松(L.principis-ruppechtii),新疆落叶松(L.sibirica)。
Ⅱ,Ⅲ阶段寄生山杨(P.davidiana),青杨(P.cathayana),响叶杨(P.adenopada),日本山杨(P.sieboldii),毛白杨(P.tomentosa),毛山杨(P.tremulavar.villosa),山白杨(P.davidiana×P.alba),山灰杨(P.davidiana×P.canescens,P.davidiana×P.davidiana,P.davidiana×P.davidiana×P.tremula,P.erecta,P.alba×P.alba,P.alba×P.davidiana,P.alba×P.sieboldii,P.alba×P.tremula,P.sieboldii×P.davidiana)。
分布:中国、日本、美国、俄罗斯、欧洲和非洲。
讨论:M.populnea常常被作为复合种处理,涵盖白杨派及青杨派上的种类,有的学者将寄生在白杨派的栅锈菌根据锈孢子寄主作为M.populnea的专化型[35-36]处理。一些作者将M.laricis作为M.populnea的异名来处理[37-38],但是他们却同时接受M.magnusiana、M.rostrupii作为一个独立的分类单元,于是常常将一些相同的标本鉴定为M.populnea或M.magnusiana,造成不必要的混乱。
这个种与寄生白杨派的其他已经报道的种类,如M.magnusiana等在夏孢子和冬孢子形状及大小上很相似,但是该种的夏孢子壁在赤道部位略有加厚,而且部分夏孢子有时候在赤道部具有小的光滑区。另外,其它白杨栅锈菌的夏孢子堆常产生于叶柄、嫩芽上,并且集合成鲜黄色的堆。
袁毅根据寄生在P.davidiana和P.cathayana的栅锈菌的少数夏孢子在赤道部具有光滑区而将它们鉴定为M.abietis-canadensis,并认为少数夏孢子赤道部的光滑区是一个重要的鉴别特征[13]。Hiratsuka和Karube(1969)描述日本的M.laricis(=M.populnea)具有加厚的壁和光滑区[17]。作者观察了采自世界不同地区山杨上的夏孢子特征,基本上都具有加厚的壁和部分光滑区,而且分子特征显示它们和其他M.populnea归于同一个进化枝[18]。因此,M.abietis-canadensis(Farl.) Ludwig作为中国新记录种是一个错误的鉴定,应该为M.populnea。
M.populnea通过转主寄生落叶松来完成生活史,Hiratsuka证实了日本山杨上的栅锈菌和日本落叶松的寄生关系[29-30],尚衍重等[12]也通过接种试验证实了M.laricis(M.populnea)的转主寄主为黄花落叶松、落叶松、日本落叶松、华北落叶松和新疆落叶松等,同时也发现该菌的锈孢子在自然和人接种时都不能侵染黑杨派和青杨派以及它们的杂交品种。
1.8 粉被栅锈菌
MelampsorapruinosaeTranz.,Tranzxchel et Serebriannikov,Mycotheca Rossica,No.265,1912
0,Ⅰ阶段未知。
夏孢子堆叶两面生,散生或聚生,橘黄色;夏孢子球形至椭圆形,(19~32×14~25)μm,壁厚度均匀,2.4~5.7μm,具刺,刺间距0.7~1.4 μm(图2E、图2F)。冬孢子堆叶两面生,单生,红褐色;冬孢子圆柱状,(37.3~60.2×9.4~13.1)μm。
寄主:胡杨P.euphratica,灰胡杨P.pruinosa。
分布:中国、俄罗斯。
讨论:粉被栅锈菌形态上与马格栅锈菌相近,但夏孢子的壁更厚,刺间距及刺更小。在国内,仅分布在新疆、甘肃、宁夏及内蒙古。自然状况下,该种仅侵染胡杨及灰胡杨,但曾对毛白杨人工接种成功[13]。有学者将几个寄生于毛白杨和钻天杨(我们对此寄主缺乏鉴定)上的栅锈菌鉴定为M.pruinosae[22],但观察发现这些标本具有大的刺和长的刺间距以及冬孢子叶面生等特征应该是M.magnusiana。此外,尚衍重曾在合作杨上偶尔发现粉被栅锈菌和落叶松杨栅锈菌的混生现象。
2 杨树栅锈菌的系统学研究
随着系统学在真菌分类中的广泛应用,越来越多的类群的分类学改变颇多。作为难以人工培养的真菌,锈菌在DNA的提取难度大于丝状真菌,且纯度低于丝状真菌。但是,应用系统学鉴定真菌的优越性明显,本文利用转录组内间隔区对中国杨树栅锈菌做了系统学分析(表1)。结果表明,8种栅锈菌在系统树图中明显形成独立的分枝,除粉被栅锈菌支持率为67%外,其余7种支持率均>85%。此外,依据发育关系,这7种可分为3支系,分别是Melampsoraabietis-populi,M.nujiangensis支系;M.medusae,M.populnea支系;M.allii-populina,M.pruinosae,M.magnusiana,M.larici-populina支系。因此,在鉴定杨树栅锈菌时,可以通过转录组內间隔区的差异,将样本快速鉴定至某一物种。

表1 系统发育学所用标本及基因信息

图3 基于ITS序列杨树栅锈菌的系统发育树
3 中国杨树栅锈菌的分种检索表

