雪松年内径向生长节律研究
2021-04-08吕庚鑫徐军亮何存存皇开源李艺杰张玉茹魏龙鑫
吕庚鑫,徐军亮,何存存,皇开源,李艺杰,张玉茹,魏龙鑫
(河南科技大学 园艺与植物保护学院,河南 洛阳 471023)
年内径向生长节律[1](intra-annual radial growth)是树木生长特性研究的主要内容,是一年内树木生长发育过程中受环境因子和自身生物学特性共同作用形成的规律。树干径向木质部生长的起止时间以及木质部细胞分化的持续时间决定了树木的年内生长状况,是理解树木生长复杂年动态的关键[2]。因此,通过研究树木径向生长动态监测等工作,不仅可以了解树木生长的动态变化,揭示树木的生长活动规律,还可以为微树芯研究提供理论基础[3]。
微树芯技术研究前景广泛。郭霞丽等[4]研究明确了微树芯技术可以监测形成层细胞的动态变化,并且可以研究树木的初级生长和次级生长动态变化过程。S.Rossi等[5]认为通过微树芯技术对树木茎干采样,对树木的伤害小,样芯质量高。因此该技术近年来得到大多数国内外学者的认可[4]。国内利用该技术监测树木形成层以及木质部的发育过程的研究尚处于起步阶段[6]。现阶段微树芯技术被认为是监测树木径向生长动态比较可靠、有效的方法[7]。目前关于树木年内径向生长的研究还存在着以下不足:1)研究区域不够广泛。大多国外研究集中在高山林线区域[8],在国内中部地区采取此方法研究树木径向生长节律的研究还相对较少,主要在北方干旱半干旱地区利用径向生长测量仪的方法研究过树木径向生长监测[9]。2)使用方法较为落后,误差较大。目前国内学者采用树木径向生长测量仪(dendrometer)连续性监测树木的径向生长的动态变化,并模拟其季节生长量[10-12]。然而,树木径向生长测量仪直接测量的数据有一定的限制性,生长季初期的水分影响会覆盖径向生长信息,例如在生长季初期或生长季期间突然大规模降水,木质部细胞会吸水膨胀,茎干会明显增粗,干旱缺水时期细胞收缩,茎干增量会出现负值,不能准确直观的获取树木生长过程的重要生理时期节点,如形成层活动启动及停止时间,木质部细胞生长速率出现的准确时期等[10,13-14]。3)以往研究都趋向于树木连续多年、多树种径向生长监测[15-16],而忽视了年内径向生长的动态变化与气候因子内在关系。准确探究树木生长节律对气候变化的响应机制,需要在不同地区开展不同类似专题研究进行比较[10]。针叶树被认为是适于进行树轮气候学分析的树种[17]。相对来说,利用微树芯技术对暖温带地区针叶树进行年内径向生长动态变化监测的研究在国内较少,在整个生长季进行各阶段细胞动态变化的研究涉及更为缺乏,因此在温带地区开展针叶树种形成层活动规律及木质部细胞分化的研究显得尤为必要。
本研究拟以暖温带常绿针叶树雪松(Cedrusdeodara)为研究对象[18],采用微型生长锥进行周期性取样,后期对雪松样芯进行石蜡切片制作[19],获取形成层和木质部细胞活动的起止时间及生长速率等生长指标数据,利用Compertz生长模型,进行年内细胞增量曲线拟合[20],研究雪松年内径向生长的变化规律,并进一步结合雪松生长年内气象数据,探讨气候因子对树木年内径向生长的影响。
1 材料与方法
1.1 研究区概况
河南科技大学校园位于河南省中部偏西(34°40′06″-34°41′02″N,112°26′02″-112°27′06″E),海拔142 m,地处暖温带南缘向北亚热带过渡地带,属于半干旱半湿润区[21]。年平均气温14.8℃,历年平均年降水量647 mm,试验地为褐土,地势平坦[22]。
1.2 样品采集
采样时间为2018年3月至2018年12月,本试验对3棵雪松进行为期近9个月的连续监测,7~15 d为一个采样周期。选取位于河南科技大学农科实验楼前用于行道树的雪松,选取其中3棵树干通直、无病虫害、无机械损伤、年龄、树高、胸径基本一致的雪松(表1)。沿树皮向髓方向,用微型生长锥[5](Trephor;采样头长度15 mm,采样头长度直径1.9 mm),在胸径处钻取树芯,每次每棵树钻取3~4个样芯。采样后封存于装有50%乙醇和甘油(丙三醇)混合溶液=1∶1的小瓶中,进行固定和软化,盖紧瓶塞,瓶内抽气真空,立即带回实验室避光,低温保存。共计采样30次,共采集微树芯约300个,完成室内石蜡切片180片。

表1 样树雪松的基本特征
1.3 样品处理
将处理好的微树芯放入全自动脱水机(型号Leica ASP200S)中,依次用浓度为50%、70%、90%、100%的乙醇梯度脱水,二甲苯和石蜡等药品使细胞透明和浸蜡。用自动石蜡包埋机(型号Leica EG 1150H)进行包埋,采用轮转切片机(Leica RM 2235),在烘片机上65℃烘干15 min,之后用二甲苯进行脱蜡,用70%、95%和100%的乙醇溶液复水,用1%番红水溶液和0.5%固绿95%乙醇溶液进行细胞染色,最后用中性树胶封片。用Leica显微镜(Leica DM 2500)分别在5倍镜和10倍镜下进行全片拍照。
1.4 数据处理
用Image J软件测量年内径向生长的长度,量3条求其平均值。观察并记录形成层细胞数量,扩大加厚期细胞数量和成熟期细胞数量。将记录到的3棵树的细胞数量和长度数据求平均,可以得到雪松累积径向生长的年均值序列。为了评估雪松茎干径向生长的动态,使用 Origin 2018中包括的非线性回归程序,对每个位点的木质部细胞总数采用Gompertz函数建立生长模型[23]。此外,通过对Gompertz函数的一阶求导可知生长季节的细胞生长速率。
Gompertz函数模拟公式如下:
Y=Ae-e(-k(x-xc))
(1)
式中,Y:径向生长木质部细胞累积量;k:内秉生长率;A:上渐近线;x:时间(年序日DOY);xc:最大增长速率出现的时间[15]。
1.5 气象数据采集与处理
气候资料来自于中国气象科学数据共享网(http://data.cma.cn),记录时间间隔为1 h,包括空气温度(TA)、降水量(P),气象数据用Excel记录保存并处理,采用origin 2018软件完成图表制作。
2 结果与分析
2.1 年内形成层细胞活动过程
结果显示,雪松在年内生长阶段,形成层细胞活动时间为3月中旬到11月中旬,持续时间为239 d。4-10月活动较为活跃,即第97天到第305天。在整个生长季期间其形成层活动呈现明显的双峰分布(图1)。形成层细胞在70 d开始活动,4月初期(97 d)至5月,形成层细胞持续增长,5月份出现形成层活动双峰的第1个峰值(7~8个)。7月中旬至9月中旬,细胞数量有所减少。形成层活动双峰的第2个峰值出现在9月底至10月中旬(第272~294天)。之后形成层数量逐渐较少,到11月中旬恢复初始水平。这与肖生春等[24]的研究结论相似。

注:年序日是指day of the year以DOY表示。
2.2 年内木质部扩大加厚期细胞活动过程
根据结果显示,在生长季期间,扩大加厚期细胞活跃时间为4月初到11月初,即第97天到第305天,扩大加厚期细胞生长持续时间为239 d,活动过程呈明显的双峰分布(图3)。扩大加厚期细胞在83 d开始出现,3月中旬(第83天)至5月上旬,细胞数量持续增长,5月中下旬出现活动双峰的第1个峰值(17~18个)。6月至7月,细胞数量略微减少。扩大加厚期活动的第2个峰值出现在8月(第208~243天),之后扩大加厚期细胞数量逐渐减少,到11月中旬减少到0个。

注:CZ表示形成层细胞。

图3 扩大期和加厚期细胞数量与年序日的关系
2.3 年内木质部成熟期细胞活动过程
成熟期细胞因细胞壁厚,故染色较深,呈现深紫色(图4)。成熟期细胞的活动时间为4月中下旬到11月底(即第104~324天),活动持续时间约为220 d,成熟期细胞数量的年生长动态变化呈现“S”型曲线(图5、图6)。成熟期的结束时间即为生长季的结束[25]。完全停止生长时,监测到年内平均累积成熟细胞个数为178.3个。

注:EN表示扩大期细胞,WT表示加厚期细胞,M代表成熟期细胞,比例尺长度为50 μm。

图5 成熟期细胞数量与年序日的关系

图6 Gompertz函数模拟雪松年内径向生长变化
2.4 雪松径向生长过程模拟
利用Gompertz生长模型对雪松茎干径向变化进行拟合,拟合后表现为“S”型曲线(图6)。径向累积生长量上限值为5 276.7 μm,累积径向细胞增量上限值178.3个。Gompertz模型能较好体现雪松的年内径向累积生长动态变化[11],拟合度达到了98%。在3月中旬(即第83天)雪松径向生长开始,5-6月,雪松进入快速生长时期(大约60 d),随后7-9月径向生长速率变慢,直至11月停止。对Gompertz拟合方程进行求导,获得生长季雪松径向生长速率的拟合曲线(图7),表现为单峰型曲线,峰值出现的时间为一年中的第162天(即Gompertz拟合方程拐点),雪松茎干径向日均生长最大速率为36.7 μm/d,径向细胞增量最大速率为1.15个/d。
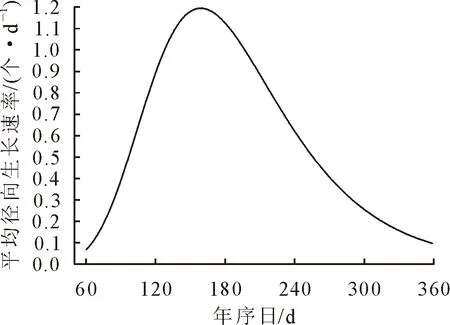
图7 Gompertz函数模拟的雪松年内径向生长速率
2.5 洛阳气象要素特征
由图8可知,洛阳地区的2018年的日均气温呈近似正态分布,在6月中旬至8月底期间温度达到最高,日均气温在25~30℃变化。洛阳地区的降水量主要集中在6月中旬至7月底(第166~208天)和9月中下旬(第260~272天)2个时间段,第1阶段降雨持续时间42 d,最大日均降雨量高达50 mm,占全年降雨量的54%。第2阶段持续降雨持续时间12 d,最大日均降雨量高达43.5 mm。3月初(第60天)和4月中下旬(第110±5天)均有少量降雨。

图8 洛阳气象站逐日平均气温和降水量(2018年)
3 结论与讨论
3.1 结论
利用微树芯技术对洛阳地区的雪松年内径向生长过程进行了周期性监测,得出了雪松的年内径向生长规律。并结合当地气象资料,探讨了主要环境因子温度和降水量对年内径向生长的影响,得出如下结论。
研究结果表明,雪松茎干形成层的细胞和扩大加厚期的细胞数量在生长季期间呈现明显的双峰动态分布,双峰出现时间分别在5月和8-9月,生长季期间形成层和扩大加厚期细胞数量在6~8个和12~18个变化。形成层细胞和扩大加厚期细胞的数量变化是感受环境变化最敏感的指标。
通过Gompertz函数模拟雪松生长过程可知,雪松木质部生长活动主要发生在3-11月,木质化平均持续时间为251 d,生长过程呈现“S”型曲线,生长过程完全符合“慢-快-慢”的生长规律[26]。最大生长速率出现时间为5月底至6月初,平均最大生长速率为1.15个/d (36.7 μm/d),持续时间为15~18 d。年内平均累积生长细胞个数为178.3个(年内平均累积径向生长长度为5 276.7 μm)。
在生长季期间,不同生长阶段,洛阳地区雪松的径向生长受到温度和降水的共同影响,且降水影响更加明显,这种影响的滞后效应期约为1个月。早期(4-7月)温度和降水对径向生长有促进作用,夏季高温时(6-8月),高温抑制径向生长,降水促进径向生长。
3.2 讨论
3.2.1 雪松年内径向生长细胞动态变化 雪松形成层细胞活动呈现双峰分布,细胞活动时期个数6~8个。峰值时间为5月和9月,峰值细胞个数为7~8个,活动时间持续时间239 d。木质部扩大加厚期细胞活动呈现双峰分布,在5月下旬和8月期间有最大值,分别为18.3个和15.6个。木质部成熟细胞活动呈现“S”型曲线[24],活动持续时间为251 d。通过Gompertz函数模拟雪松生长过程,发现在第162天,径向生长细胞个数和长度均出现最大值,分别为1.15个/d,36.7 μm/d。年内平均累计生长细胞个数达到178.3个,年内平均累计生长量达到5 276.7 μm。华北地区的华北落叶松(Larixprincipis-rupprechtii)和白桦(Betulaplatyphylla)最大年生长量分别为0.41 cm和0.44 cm,与本研究结论基本一致[27]。本研究发现暖温带地区雪松的最大径向生长速率出现的时间在6月11日,比梁尔源等[28]的研究结论提前10 d。最大径向生长速率出现在年内日均最高气温之前[29]。
3.2.2 温度对年内径向生长的影响 由年内径向生长细胞数量动态变化和年内日均气温进行分析可知(图1、图3、图8),雪松茎干径向生长经历了4个不同的阶段,主要体现在早期低温抑制,中期温度促进,后期高温抑制,末期低温抑制。与董满宇等[11]的研究高山林线树种白杄(Piceameyeri)茎干径向生长结果(3个阶段)稍有不同,主要考虑是地域之间的差异,高海拔地区的年均气温较低,在夏季也未能达到影响形成层活动的高温临界点。崔克明等[30]研究阔叶树杜仲也有类似结论,并认为第3阶段是形成层的生理休眠期。温度在生长季(3-11月)不同的时期对茎干径向生长的影响不同。这与田义超等[31]的研究结果类似。洛阳地区雪松在早期(3月)温度占主导因素[6],土壤低温限制树木根部对土壤养分和水分的吸收,进而影响径向生长进程[32]。4-6月适宜的温度促进茎干径向快速生长[11],7-8月夏季高温导致形成层和扩大加厚期细胞数量有所下降,呈负相关,径向生长速率减慢[33]。说明洛阳的夏季持续高温一定程度上影响了雪松的年内径向生长:1)高温加速土壤水分蒸发,土壤含水量下降,造成水分胁迫,树木调节自身生长代谢水平,通过缓慢甚至停止生长以度过高温干旱的逆境。2)夏季高温会使土壤温度升高,高温抑制雪松根系生长甚至造成根系老化,根毛减少,吸水面积减小,吸水能力下降,树体整体水分供应不足,形成层细胞分裂受阻,数量有所下降,进而影响径向生长进程[8]。3)夏季的持续高温使土壤升温,从而限制了树木的水分利用速率,树木在高温条件下通过蒸腾水分自身降温,因此树木的生长受到抑制。高温影响树木径向生长具有滞后效应,本研究发现高温对雪松的滞后期为1个月。这与张赟等[34]的研究结论基本一致。
3.2.3 降水量对年内径向生长的影响 由年内径向生长细胞数量动态变化和年内日均降水量进行分析可知(图1、图3、图8),洛阳地区的雪松年内径向生长与降水量呈现明显的正相关,4月中下旬的短暂降水极大促进了径向生长的进程,降水提高了土壤含水量,促进根系快速吸水,促使形成层和扩大加厚期细胞数量出现了第1次峰值,表现为茎干径向增长加快,这与A.Turcotte等[35]的观测结果一致。当6月中旬降雨量增加时,平均气温虽然变化不大,但是降水缓解了高温脱水带来的干旱胁迫,充沛的雨水促进细胞膨胀增大和分裂,因此对径向生长具有明显的促进作用[33,36-37]。同时也说明雪松的径向生长受到了水分的限制[38]这与B.Franco等[39]和曹受金等[40]的研究结论一致。韩艳刚等[41]研究认为6月份的大幅度降水对针叶树的径向生长有显著促进作用,与本研究结论基本一致。
