细胞衰老及相关疾患的circRNA机制及功能
2021-04-07余素素张文娟
余素素,张文娟
(暨南大学 基础医学与公共卫生学院,公共卫生与预防医学系,广东 广州 510632)
人口老龄化是一个正在持续扩大加深全球化的普遍趋势.衰老及其相关疾患,如阿尔茨海默病、慢性获得性进行性智能障碍综合征(俗称“痴呆”)、动脉粥样硬化和多种肿瘤癌症等,是全人类亟待解决的健康相关问题[1].积极应对人口老龄化及其相关疾病问题,也是当前“健康中国2030”规划的工作重点之一.衰老是指机体随年龄增长,呈现出各细胞、组织和系统发生退行性变化的种种征象.细胞是机体衰老的原始结构.长寿基因的发现[2]、表观遗传学以及表观转录组学[3]等研究领域的迅猛发展,使得衰老及相关疾患研究进入新领域.
1976年首次在植物类病毒和副流感病毒颗粒中发现circRNA[4],这种特殊结构的RNA曾被误认为是错误的基因转录副产物[5],但随着高通量测序技术和生物信息学的发展,circRNA已然成为研究新热点.circRNA是一类特殊的长链非编码RNA(lncRNA),cirRNA主要来源于细胞母基因的非编码区域,通过反向剪接反应,以共价键连接形成不具有5′末端“帽子”结构和3′末端poly(A)尾巴的RNA分子结构,这一特殊的环状结构使得circRNA不易受到RNaseR外切酶影响,从而有效稳定保持其功能[6];该结构中常富含有多种microRNA(miRNA)结合位点,具备有吸附miRNA的“海绵体功能”,通过与相应RNA结合蛋白(RNA-binding proteins, RBPs)及核糖核蛋白复合体(ribonucleoprotein complex, RNP)结合,起拮抗竞争作用,抑制调节基因表达[7],如图1所示.
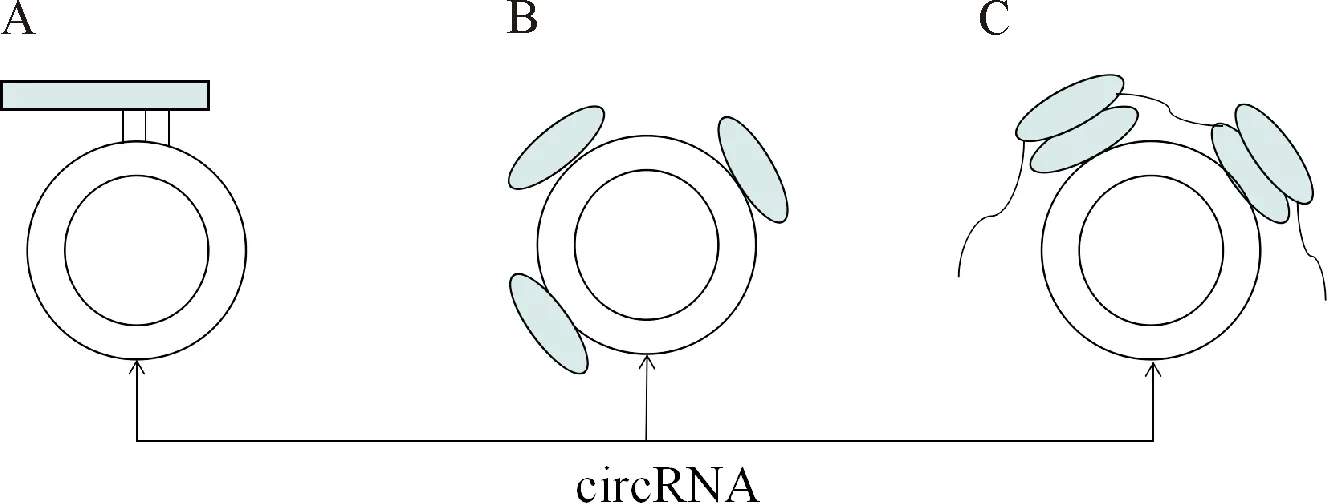
A:miRNA“海绵体”吸附功能;B:结合RNA结合蛋白和核糖核蛋白复合体;C:翻译蛋白质A:The absorption function of ‘miRNA’ sponges;B:binding with RNA binding protein & ribonucleoprotein complex; C:The translation of proteins图1 circRNA功能示意图Fig.1 The function of cirRNA
目前已证实circRNA参与调控细胞基因的转录、翻译和表达,影响细胞周期.circRNA可通过经典剪切、反向剪切和选择性剪切等模式调控基因的表达,有效扩展基因转录的多样性[8].circRNA还可与细胞周期蛋白依赖性激酶2(cyclin-dependent kinases 2,CDK2)和周期蛋白依赖性激酶抑制剂1(cyclin-dependent kinase inhibitor 1,CKI或p21)结合,可通过哺乳动物叉头转录因子(forkhead transcription factor,FOXO)等多种方式来抑制细胞增殖[9].circRNA在机体内呈特异性的丰富表达,具有多种生物学功能,在血液中具有更长的半衰期,比起同源亚型RNA更具稳定性和保守性,是组织和体液检测可靠的生物标志物,可作为多种疾病(心血管疾病、神经退行性疾病和肿瘤等)临床筛检、诊断和预后的特异性生物标志物[10].circRNA在真核细胞中广泛表达,在基因分子水平上发挥重要的调控作用,以多种形式参与调控细胞周期,参与调控器官系统及机体的衰老进程.
1 circRNA与神经细胞衰老
果蝇体内的circRNA随年龄增长而逐渐累积,并在其中枢神经系统老化过程中,高度特异性表达,且具有发育和组织特异性.与同源线性亚型相比,环状RNA明显增多[11],circRNA在人与果蝇的大脑细胞中均特异性表达,表明circRNA可作为神经系统衰老的早期生物标志物,参与调控神经系统的衰老,并适用于多个物种.circRNA在衰老细胞与亚细胞间的表达模式尚待进一步研究,但明确的是,circRNA富集于神经元细胞间的亚单位结构(轴突、树突和突触),而神经系统的衰老通常与海马体和前额叶皮层中特定的、相对微妙的突触改变有关[12].树鼩神经细胞中83.1%的circRNA与人类具有同源性,经相互作用调控网络发现,circRNA_007362可通过吸附24个miRNA调控UBE4B的表达,参与诱导神经元细胞的凋亡和神经系统的衰老[13].神经退行性疾病,如阿尔茨海默病和帕金森症等,与神经细胞衰老密切相关.Aβ1-42寡聚体诱导阿尔茨海默病小鼠研究中,神经细胞的mmu_circRNA_017963参与其自噬体组装、胞吐作用和凋亡过程,进而诱导神经细胞衰老[14].散发的阿尔茨海默病可能与ciRS-7异常表达有关,其作为一种内源性的miRNA海绵,吸附并抑制正常miRNA-7的功能,使调控miRNA-7-circRNA表达的系统失效,诱导神经系统衰老,疾病产生[15].尽管circRNA分子在多个物种的神经系统中均有高表达,具有上调神经元分化和高度丰富突触的功能,但其具体的作用途径和机制尚不清楚.特异性circRNA表达与神经系统有害结局之间的关联作用,不同circRNA表达之间的联合,及特定circRNA功能改变对人类神经系统衰老及相关疾患进程的影响,将使神经系统特异性circRNA机制研究更具有潜在的临床应用价值.
2 circRNA与肌肉、纤维细胞衰老
随年龄增长,心肌细胞和骨骼肌肉细胞衰老加速,细胞内基因表达与肌肉细胞年龄相关性综合征的发生发展密切相关[16].miR-206、miR-29和miR-431等non-coding RNA(ncRNA)参与骨骼肌系统的生理、病理过程[17],而特定circRNA基因序列的特殊结合位点与细胞中相应的因子结合,参与调控肌细胞的生长发育,或诱导肌细胞发生病理性改变[18],如少肌症等.转录因子circ-Foxo3产生的circRNA在老年患者和老年小鼠的心脏样本中均高表达,其变化趋势与细胞衰老的其他标志物一致.DNA结合蛋白抑制剂-1(inhibitors of DNA Binding-1,ID-1),转录因子E2F-1 (transcription factor, E2F-1),局部粘着斑激酶(focal adhesion kinase,FAK)和低氧诱导因子-1α(hypoxia inducible factor 1,HIF1α)与circ-Foxo3相互作用后滞留在细胞质中,使心肌细胞失去抗衰老和抗应激功能,加速心肌细胞的衰老进程.circ-Foxo3异位表达同样可诱导小鼠肌肉细胞衰老.在多种小鼠成纤维细胞中,过氧化氢可诱导circ-Foxo3过表达并促进细胞衰老,相反,沉默circ-Foxo3可抑制细胞衰老.特异性表达的circRNA可作为肌细胞衰老及其相关疾患的标志物,也对外界因素引起的衰老及病变进行应答,例如,阿霉素可诱导circ-Foxo3过表达,促进心肌细胞早衰病变,而沉默circ-Foxo3可缓解阿霉素的作用并抑制肌细胞衰老[9].circRNA-miRNA-mRNA交互网络发现circRNA014844、circRNA011308和circRNA018401可通过吸附miR-493-5-p调节衰老过程促凝因子FGB的表达,诱导心肌细胞衰老,产生病理改变[16].
circPVT1由PVT1外显子环化产生.在电离辐射诱导衰老成纤维细胞中,circPVT1表达丰度降低,结合衰老相关的β-半乳糖苷酶活性的升高,CDKN1A/P21和TP53的丰度增加及细胞增殖减少,进一步诱导结缔组织衰老进程.circPVT1选择性地富集let-7(该基因减少表达促使抑癌基因表达加强),而降低其表达丰度可选择性地下调let-7下游基因,IGF2BP1、KRAS和HMGA2等抗衰老的增殖蛋白的表达,最终促进衰老发生[19].circRNA_100797作为miR-23a-5p的海绵,两者在中波紫外线诱导的早衰人成纤维细胞中呈负相关表达,高表达的circRNA_100797具有光保护作用,延缓细胞衰老[20].circRNA的特异性表达通过多种形式影响细胞的衰老进程,特异性表达的circRNA可作为肌肉、纤维细胞衰老及其相关疾患的生物标志物.不仅外界环境因素可调控特定circRNA的表达,诱导细胞的衰老过程;而且通过调控circRNA的表达丰度也可调节外界刺激诱导的衰老进程.通过上调或沉默某些特定circRNA实现临床调控机体的肌、纤维细胞的衰老及相关疾病,使circRNA的机制研究更具有实际意义.
3 circRNA与外周血细胞衰老
circRNA在人类外周血循环中有特异性表达[21].circRNA特异性表达可调控ATP代谢过程以及突触信号传递,形成相关基因互作网络的反馈调节系统,主要富集于与吞噬、昼夜节律调节、癌症途径和高尔基体相关的细胞囊泡中.InCHIANTI人群是居住在意大利托斯卡纳基安蒂地区的代表性老年人口,即对研究对象的年龄、健康情况、父母双方的死亡年龄、寿命评分限制并随访的人群.随访队列第三、四代65岁以上的该人群中,其外周血细胞中circITGAX、circPLEKHM1、circDEF6、circATP6V0A1和circASAP等分子表达丰度低于30~36岁的年轻组,而circ-Foxo3、circFNDC3B、circAFF1、circCDYL和circXPO7则在86~95岁老年组中外周血细胞中表达丰度增高[22].免疫相关因子可通过外周血循环到达机体全身发挥作用,随着年龄的增长,免疫器官逐渐老化、免疫细胞及免疫因子的机制水平逐渐降低,机体免疫功能下降,许多免疫衰老相关疾病,如感染性疾病、肿瘤的发生等,导致群体死亡率增高[23].circ_0132266具有 miR-337-3p“海绵体”吸附功能,通过靶基因circ_0132266-miR-337-3p-PML轴,其上升性表达可显著促进外周血单核细胞的活性[24].通过基因转导持续表达的CD28可延缓复制性衰老的过程,增强端粒酶活性、增加整体增殖潜能、促炎细胞因子分泌减少.circRNA_100783参与T细胞衰老的CD28及CD8(+)相关蛋白磷酸化的过程,进而促进衰老发生.因此,circRNA_100783可作为免疫细胞衰老和免疫系统衰老过程新的生物标志物[25],另外,发现在系统性红斑狼疮患者体内HSA_CIRCpedia_7637表达下降,推测其可能通过竞争性结合hsa-miR-638来参与免疫系统疾病的发生[26].目前尚未阐明不同来源的同型circRNA产生不同作用的内在原因和调控机制,circRNA有望成为外周血细胞以及相关免疫疾病诊断的生物标志物和相关药物治疗靶点.

表1 circRNA与机体细胞衰老的功能关联Table 1 The functionality correlation between circRNA and somatic cell senescence
4 circRNA与肿瘤细胞衰老
除了细胞衰老之外,衰老反应亦会使细胞突变累积,相互作用,增加患癌风险,有些突变更是人类肿瘤恶性转化发生的必要条件,肿瘤的发生发展可能是哺乳动物机体系统逃逸衰老的标志[27].circRNA的异常表达可调控肿瘤癌症的发生发展,hsa_circ_0055176在口腔鳞癌低表达可增强口腔鳞状细胞癌细胞的迁移和侵袭能力[28].体内、外研究实验发现染色体易位编码的融合蛋白和融合circRNA(fusion circ RNAs, f-circ RNA)与其他致癌物质均能提高细胞增殖率,促进细胞转化和肿瘤的发生,胞内的f-circM9表达可使癌细胞逃逸药物诱导的凋亡[29].肿瘤细胞中circRNA特异性表达与肿瘤细胞衰老进程存在密切相关.如表2所示,肿瘤的发生发展常与细胞中某些特定circRNA的特异性表达相互作用,可以启动肿瘤细胞的衰老发生与否.circRNA可作为肿瘤的辅助临床诊断标志物,而能否通过调控circRNA的表达控制肿瘤细胞的周期,仍需进一步的研究.

表2 circRNA与肿瘤细胞衰老的关联Table 2 The correlation between circRNA tumor cell senescence
5 展望
circRNA的生物产生、结构和功能的系统认识,其特异性表达与机体生理病理过程联系的探究逐步加深.衰老相关的circRNA变化特征不仅反映细胞渐行性退化,也反映出细胞内分子适应衰老的反应.circRNA的特异性表达与细胞衰老及其相关疾病密切相关,外源环境因素,如药物、射线、氧化物等刺激也可影响细胞内相关通路,进而调控细胞衰老进程[14, 20, 28].因此,特定时间和环境刺激作用下,引起的细胞内关键分子特定circRNA的表达上调或沉默,可作为衰老及相关疾患的早期生物学标志,在临床诊断中,circRNA可与常规癌症标志物癌胚抗原(carcinoembryonic antigen,CEA)、CA 125、CA 153、前列腺特异抗原(prostate specific antigen,PSA)和甲胎蛋白(alpha fetoprotein,AFP)结合应用.另外,通过其特异性表达也能从分子水平上尽早发现疾病,在未出现临床症状时预测衰老相关疾病的发生,判断疾病的发生阶段,某些circRNA还可作为疾病预后评价标志物[40].为其预防、筛检、诊断和治疗提供新的思路,也为响应“健康中国2030”规划纲要及健康中国的全周期生命健康行动提供科学理论基础.目前,对circRNA与衰老及相关疾患的实验研究仍较少,相关综述研究认为较其他RNA转录物,circRNA与衰老间有更强的相关性[41],在不同物种衰老过程中均存在circRNA特异性表达,因此,circRNA可能是衰老和年龄相关疾患的潜在的致病因素,而高稳定性可能是其随增龄高表达现象[42].
总之,细胞衰老是一个多因素影响的过程,circRNA通过其特殊结构与调控RNA和蛋白质结合,发挥相应生物学作用,可影响衰老的进程.随着相关研究的进行,对circRNA产生和基因调控机制的总结,阐述其作为新的疾病诊断和预后标志物的可能,为衰老及相关疾患的复杂调控网络增添新的研究思路[10, 41-42].随着高通量分子测序技术的发展为相关疾病标志检测提供技术可能,本研究阐述了在不同组织细胞中,特异性circRNA表达在衰老及相关疾患发生发展中的内在调控机制,多种circRNA及其他因子的协同作用,内源性和外源性的同型circRNA表达的效应差异,这些将为衰老及其相关疾患的病因诊断和分子机制提供新的研究方向,但circRNA作为衰老及相关疾患的标志物临床应用的可靠性和灵敏性仍需要进一步的深入研究.
