栓皮栎伐桩萌苗生物量积累动态及其影响因素的研究
2021-04-06吴桐郭会云郑依腾马闯吉凯高贺毅
吴桐,郭会云,郑依腾,马闯,吉凯,高贺毅
栓皮栎伐桩萌苗生物量积累动态及其影响因素的研究
吴桐,郭会云,郑依腾,马闯通信作者,吉凯,高贺毅
(天津农学院 园艺园林学院,天津 300392)
伐桩萌苗的生物量是反映次生林地更新水平的重要标志,探明影响其积累的主要因素对森林经营和保护具有重要意义。以天津北部山区栓皮栎林为对象,研究其采伐后近5年的伐桩萌苗生长规律,构建生物量积累的异速生长方程,分析气候变化对其生长的影响。结果发现:该地区栓皮栎采伐后5年萌苗直径(1.61±0.44)cm,长度(143.02±9.81)cm,累计茎干干物质量为(75.49±10.51)g/株,前3年生物量积累较慢,随后快速增加;构建异速生长方程表明,直径与茎干重两组变量符合=a2+b+c模型,而长度与茎干重符合=a2+c模式,均达到显著性;Pearson相关分析表明,伐桩萌苗生长与降雨呈正相关,而与温度呈负相关,表明高温少雨是限制该地区栓皮栎伐桩萌苗生物量积累的因素。在今后的次生林经营中,要注意采用水肥管理、人工促萌、复壮等技术,促进次生林更新。
栓皮栎;伐桩萌苗;异速生长方程;Pearson相关分析
伐桩萌苗是指地上茎干经过采伐,或牲畜啃食、火灾、虫蛀等破坏后,依靠残留树桩上的不定芽和休眠芽萌发形成新的植株。由于其生长快、适应力强、有驻留生态位等特点,能迅速填补干扰所形成的林隙和空地,减少对种子的依赖,可加速林分的恢复[1]。以往有关伐桩萌苗的研究,主要集中在采伐次数、伐桩大小以及竞争强度的影响等方面[2-4],而有关伐桩萌苗干物质积累能力的研究较少。在次生林经营过程中,伐桩萌苗的干物质积累不仅反映了林地的生产能力,也是碳汇水平的重要体现[5]。因此,探究影响伐桩萌苗干物质积累的因素具有重要的理论意义。
栓皮栎()是壳斗科(Fagaceae)栎属()植物,是我国暖温带落叶阔叶林、亚热带常绿落叶林区地带性植被的主要建群种,在华北地区大量分布[6]。以往对栓皮栎的研究主要集中在种群生殖动态、年龄结构和生物学特性等方面[7-8]。余碧云等[9]研究指出栓皮栎萌苗地表部分生物量的配比为树干生物量占71.8%~88.4%,枝生物量占5.8%~19.6%,叶生物量占4.2%~8.6%。然而,有关栓皮栎伐桩萌苗生物量积累的动态研究还较少。本试验采用“空间代时间”的方法,对同一伐桩萌苗上不同生长年限的茎段进行测量,探究栓皮栎伐桩萌苗生长的年际规律,建立异速生长方程为伐桩萌苗更新过程生物量测算提供理论基础;探究影响伐桩萌苗生长的气候因素,为华北地区典型栎林保护和恢复提供指导。
1 材料与方法
1.1 研究区概况
试验地位于天津北部蓟州区八仙山国家自然保护区外围。北纬40°10′~40°14′和东经117°30′~117°36′,属暖温带半湿润大陆性季风气候。年平均气温10.1 ℃,年降水量970 mm,年积温4 153 ℃,无霜期105 d。土壤为山地森林褐色土,枯枝落叶层较厚,呈酸性。
1.2 样地设置
选择位于古强峪村自留山的栓皮栎林建立样地。2011年春,因村民建材需要部分林木被砍伐,形成栓皮栎次生林。先于2018年5月中旬确定栓皮栎伐桩萌苗试验样地,样方面积10 m×10 m,共建立6个试验样方,每个样方选择3株生长一致的伐桩萌苗并用红色油漆进行标记。
1.3 生长量与干物质量测定
2018年9月份在试验地进行生长量测定,根据主茎上芽鳞痕、茎条颜色、木质化程度可明显区分近5年的生长量,主茎上有明显的节可作为年际区分[10]。由前端往下,用游标卡尺测量同一主茎近5年节间的直径,前中后3段取平均值。用直尺测量节间长度,同一生长年限不同萌苗直径和长度取平均值(=18)[11]。
生长量测定完成后,使用手锯将萌苗采割带回实验室。用枝剪在节处将主茎剪切成段,不同年份的茎段和叶片分类标记。将标记好的试验材料在80 ℃的烘箱中烘干72 h脱水处理后,使用千分之一电子天平测量枝干重和叶片重。
1.4 气候数据获取
研究样点位于蓟州山区,缺乏基础的气象资料。查阅CRU TS3.23数据库,获得研究点2014—2018年逐年逐月的平均气温、同期的降水量值。CRU TS(Climatic Research Unit time-series)数据库是基于全球4 000个气象站数据,按高精分(0.5×0.5栅格化)获得的平均气象数据。
1.5 数据分析
采用Excel 2010计算每组数据的平均值和误差值,并绘制直径、长度、枝干重、叶片重生长折线图。用spss20.0比较不同生长年限伐桩萌苗直径与长度的差异,并用 Duncan 新复极差法对数据进行多重比较(α=0.05)。对栓皮栎伐桩萌苗的直径和枝干长度、直径和枝干重、长度和枝干重等参数取对数,分别进行二次多项式回归分析,回归分析结果绘制散点图。采用1stOpt15PRO软件拟合二次多项式回归方程,构建生长量指标间的异速生长方程[12]。不同生长量与气候因子的相关分析采用R软件(corplot Package)分析并作图。
2 结果与分析
2.1 伐桩萌苗逐年生长规律
由图1可以得出,该地区栓皮栎伐桩萌苗生长1年平均直径可达(0.48±0.13)cm,而生长5年平均直径可达(1.61±0.44)cm,萌苗直径的逐年增加量在0.22~0.35 cm,且不同生长年限的茎段直径差异显著。栓皮栎伐桩萌苗长度第1年增加最快,平均长度可达(34.34±11.72)cm,5年累计长度(143.02±9.81)cm,逐年长度增加量在24.03~34.34 cm。
由图1可以看出,栓皮栎伐桩萌苗茎干干重前3年积累较慢,逐年增加量为3.86~5.86 g/株,第4年开始干物质积累较快,为(22.98±15.73)g/株,栓皮栎伐桩萌苗5年累计茎干干物质量为(75.49±10.51)g/株,占总生物量的62%。不同生长年限的茎段上叶片干重在7.52~11.53 g/株,且无显著差异,叶片总干重为(46.62±4.95)g/株,占总生物量的38%。
2.2 伐桩萌苗生长量异速方程
植物异速生长类型可分为简单的异速生长和复杂的异速生长两种。简单异速生长符合方程= a2+c的数学形式,而复杂的异速生长关系则遵从=a2+b+c的数学形式。由图2可知,对栓皮栎伐桩萌苗直径、长度和茎干重分别取对数进行线性回归分析,发现lg(ɸ)- lg(M)线性相关较强,而lg(ɸ)- lg(D)之间线性关系较弱。
由表1可知,构建不同生长年限栓皮栎伐桩萌苗的直径、长度与茎干重的异速生长方程,发现直径&长度、直径&茎干重两组变量符合=a2+ b+c模型,而长度&茎干重符合=a2+c模式。其中,第3年、第5年的直径&长度非线性回归未达到显著水平(>0.05),其余年限均达到显著水平(>0.05)
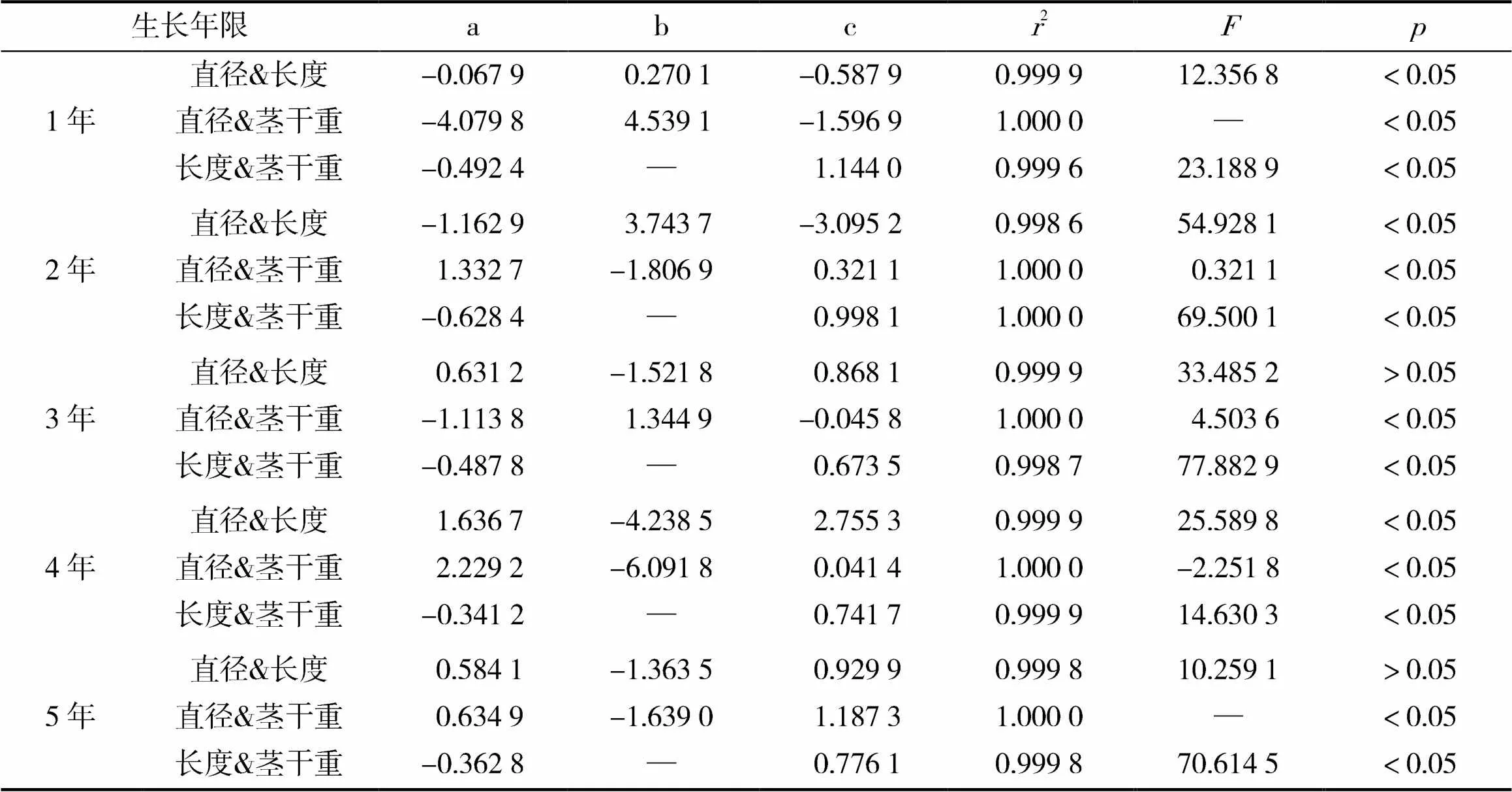
表1 栓皮栎伐桩萌苗直径、长度、茎干重异速生长模型参数
2.3 气候因子对伐桩萌苗生长的影响
天津市蓟州区八仙山地区气候因子年际变化情况如图3所示。
研究发现该地区年均降雨量为762.53 mm,其中2018年降雨量最大,为786.14 mm。不同月份降雨量差异较大,每年7月份降水量比较充足。从2014—2018年,年均温度变化处于增长趋势 (图3,虚线斜率=0.019,<0.05)。其中,2018年均温最高,为17.6 ℃,2014年最低,为13.7 ℃。
由图4所示,对年降雨量、年均温与栓皮栎伐桩萌苗生长量进行相关性分析,发现气候因子对萌苗长度影响较大,其次为萌苗直径和茎干重,而与叶片干重相关性较小(>0.05)。其中,降雨与萌苗长度、直径和茎干重呈显著正相关,而温度与其呈显著负相关。分析不同生长量间关系发现,萌苗的茎干重受其直径影响最大,其次为长度。
3 讨论
3.1 伐桩萌苗生长量的变化规律
栎类树种通常具有较强的萌蘖能力,其在采伐或茎干破坏后,能依靠伐桩萌苗快速恢复生长,从而维持林分稳定[13]。在本研究中,栓皮栎伐桩萌苗5年长度仅为140 cm,直径1.6 cm,茎干重可达75 g/株。而易青春等[3]研究发现,秦岭南坡不同生境栓皮栎伐桩萌苗单株干重在716~2 720 g/株,这可能与其所在区域为栓皮栎核心区,水热条件较好有关。此外,研究发现伐桩萌苗生长初期直径和长度增加较快,茎干物质积累较慢,而随着时间延长,直径和长度年增加量减小,干物质积累增速,说明萌苗的生长具有明显的阶段性。前期萌苗长度增加较快有利于其竞争上层的光照空间,以增加光合作用产物[14]。而随着茎干木质化程度的增加,干物质积累加快,则有利于抵御环境变化和病虫侵害[15]。一般认为,苗木的生长可划分速生期和木质化期,而后者表现为干物质量的增加[16]。
3.2 伐桩萌苗生长量的异速关系
异速关系理论指出生物量分配模式主要受植物本身大小调控,可采用幂函数的形式来表征,即异速生长方程[17]。该方程通过异速生长尺度参数来表征因变量和自变量之间的关系,并决定植物各器官生物量分配随植物大小的变化规律[18]。在以往对林木个体研究中,一般采用胸径作为自变量构建关于生物量异速生长方程,其结构简单,应用广泛[19]。本研究中,以栓皮栎伐桩萌苗为研究对象,其直径与茎干重所构建的异速生长方程均具有较高的2,说明采用直径作为估算生物量的参数同样对伐桩萌苗具有意义。然而,本研究发现,以萌苗长度作为自变量与茎干重所建立的简单异速生长方程,在不同的恢复年限均具有较高的2,说明伐桩萌苗的长度对生物量的反映更直接。CIENCIALA等[20]在对实生个体研究后认为,树高变量的加入能够改善赤松() 各组分的生物量预测效果。这为今后次生林生物量估算提供了更简洁的途径。
3.3 影响栓皮栎伐桩萌苗生长的因素分析
植物生物量的积累过程是植物自身与环境因素共同作用的结果,它既反映了植物对环境条件的适应及利用状态,也反映了植物本身的生存能力和生长发育规律[21]。在本研究中,萌苗长度与干物质呈较强正相关,说明早期萌苗生物量积累主要依靠长度的增加。庞志蕊等[22]认为,株高越高的个体,其综合排名较高,环境适应能力越强。此外,本研究发现降雨对萌苗直径、长度以及茎干重的增加呈显著的正相关,而温度与三者之间分别呈显著负相关。说明降雨充足、温度适宜有利于伐桩萌苗更新,而干旱和高温则不利于萌苗生长。王婧等[23]认为,栓皮栎在中国大陆分布区的多年平均气温为15.3 ℃,多年平均年降水量为411~2 000 mm。而秦巴山区被认为是栓皮栎的核心分布区,年降雨量在990 mm,年均温13 ℃[24]。本研究样地所在地区年均温15.65 ℃,年均降雨量762.53 mm,处于栓皮栎的边缘分布区,这可能是造成该地区栓皮栎伐桩萌苗生物量积累缓慢的原因之一。而随着华北地区暖干化趋势加剧,未来栓皮栎次生林更新将会受到更严重的影响。
4 结论
通过对天津北部山区栓皮栎次生林伐桩萌苗调查,发现5年萌苗直径(1.61±0.44)cm,长度(143.02±9.81)cm,累计茎干干物质量为(75.49±10.51)g/株。栓皮栎采伐后,前期茎干长度和直径增加较快,后期干物质积累较快,具有明显的阶段性。分别以直径和长度为自变量,以茎干重为因变量,构建异速生长模型,均具有较高的2。Pearson相关分析表明,降雨与伐桩萌苗生长呈正相关,而温度呈负相关。该地区栓皮栎伐桩萌苗生长缓慢,次生林恢复受到威胁,可能与降雨少、干旱加剧有关。今后,要注重栓皮栎次生林保护,注重水肥管理并配以人工促萌、复壮等技术,提升林地质量。
[1] BEAUDET M,MESSIER C. Beech regeneration of seed and root sucker origin: A comparison of morphology, growth, survival, and response to defoliation[J]. Forest Ecology and Management,2008,255(10):3659-3666.
[2] 马闯,张文辉,薛瑶芹,等. 邻体竞争和环境因子对栓皮栎伐桩萌苗表型特征的影响[J]. 西北农林科技大学学报(自然科学版),2011,39(10):71-80.
[3] 易青春,张文辉,唐德瑞. 栓皮栎伐桩萌苗保留量对其生长和生物量积累的影响[J]. 林业科学,2013,49(7):34-39.
[4] 易青春,张文辉,唐德瑞,等. 采伐次数对栓皮栎伐桩萌苗生长的影响[J]. 西北农林科技大学学报(自然科学版),2013,49(4):147-154.
[5] 霍常富,尤文忠,张慧东,等. 辽宁冰砬山不同林龄蒙古栎次生林生物量和生产力初探[J]. 辽宁林业科技,2011(4):4-6.
[6] 周建云,林军,何景峰,等. 栓皮栎研究进展与未来展望[J]. 西北林学院学报,2010,25(3):43-49.
[7] 马莉薇,张文辉,周建云,等. 秦岭北坡林窗大小对栓皮栎实生幼苗生长发育的影响[J]. 林业科学,2013,49(12):43-50.
[8] 冉然,张文辉,何景峰,等. 间伐强度对秦岭南坡栓皮栎天然林种群更新的影响[J]. 应用生态学报,2014,25(3):695-701.
[9] 余碧云,张文辉,胡晓静,等. 栓皮栎地上部分构型及生物量分配[J]. 应用生态学报,2015,26(8):2265-2272.
[10] 马莉薇,张文辉,薛瑶芹,等. 秦岭北坡不同生境栓皮栎实生苗生长及其影响因素[J]. 生态学报,2010,30(23):6512-6520.
[11] 陆珺,李蕊,郜伟,等. 天津市不同类型土壤彩叶植物生长特性比较[J]. 天津农学院学报,2020,27(2):35-37,43.
[12] 安慧,上官周平. 密度对刺槐幼苗生物量及异速生长模式的影响[J]. 林业科学,2008,44(3):154-158.
[13] 薛瑶芹,张文辉,马莉薇,等. 不同生境下栓皮栎伐桩萌苗的生长特征及在种群更新中的作用[J]. 林业科学,2012,48(7):23-29.
[14] WEINERJ. How competition for light and nutrients affects size variability in Ip[J]. Ecology,2008,67(5):1425- 1427.
[15] 李媛鑫,李效文,陈秋夏. 多效唑对降香黄檀容器苗抗寒性的影响研究[J/OL]. 西南林业大学学报(自然科学版):http://kns.cnki.net/kcms/detail/53.1218.S.20200316. 1149. 018.html.
[16] 沈国舫. 森林培育学[M]. 北京:中国林业出版社,2001.
[17] WEST G B,BROWN J H,ENQUIST B J. A general model for the structure and allometry of plant vascular systems[J]. Nature,1999,400(6745):664-667.
[18] 陈东升,孙晓梅,金英博,等. 林龄和竞争对日本落叶松各组分生物量异速关系的影响[J]. 生态学报,2020,40(3):843-853.
[19] 李巍. 兴安落叶松生物量分配格局和异速生长模型[D].哈尔滨:东北林业大学,2014.
[20] CIENCIALA E,ERNY M,TATARINOV F,et al. Biomass functions applicable to Scots pine[J]. Trees,2006,20(4):483-495.
[21] 张文辉,张存旭,李景侠,等. 川西泡沙参种群地上生物量生长发育的研究[J]. 西北植物学报,1999,19(5):44-53.
[22] 庞志蕊,周强,杨静慧,等. 不同类型盐碱地常见彩叶植物生长指标的相关性[J]. 天津农学院学报,2018,25(4):10-12.
[23] 王婧,王少波,康宏樟,等. 东亚地区栓皮栎的地理分布格局及其气候特征[J]. 上海交通大学学报(农业科学版),2009,27(3):235-241.
[24] 张文辉,卢志军,李景侠,等. 秦岭北坡栓皮栎种群动态的研究[J]. 应用生态学报,2003,14(9):1427- 1432.
Study on biomass accumulation dynamics of stump sprouting ofand its influence factors
Wu Tong, Guo Huiyun, Zheng Yiteng, Ma ChuangCorresponding Author, Ji Kai, Gao Heyi
(College of Horticulture and Landscape, Tianjin Agricultural University, Tianjin 300392, China)
The biomass of stump sprouting is a key indicator to reflect the regeneration of secondary forest. It is significant to find out the main factors affecting biomass accumulation for forest management and protection.forest in the northern mountainous area of Tianjin was selected as the research object. The growth dynamic of stump sprouting was studied in recent 5 years after cutting with fitting the allometric growth equation of biomass accumulation, and the influence of climate on the growth was analyzed. The results showed that the diameter and length of sprouting seedlings five years after cutting were (1.61±0.44)cm and(143.02±9.81)cm, respectively. The total stem dry matter weight was(75.49±10.51)g / plant. The biomass accumulated slowly in the first three years, and then increased speedly. The allometric growth equation showed that diameter and stem dry weight were in accordance with the model as=a2+b+c, while length and stem dry weight were in accordance with=a2+c model, both of which were significant. Pearson correlation analysis showed that the growth of stump sprouting seedlings was positively correlated with rainfall, whereas negatively correlated with temperature, indicating that high temperature and less rainfall were the factors limiting the biomass accumulation ofstump sprout in this area. In the future, the management of water and fertilizer should be strengthened, and the technology of artificial sprouting and rejuvenation should be adopted to promote the regeneration of secondary forest.
; stump sprout; allometric growth equation; Pearson correlation analysis
Q948.113
A
1008-5394(2021)01-0006-06
10.19640/j.cnki.jtau.2021.01.002
2020-08-13
国家青年科学基金项目(31600499);国家大学生创新创业训练计划资助项目(201910061112);河北省重点研发计划项目(20326810D)
吴桐(1999-),女,本科在读,主要从事森林培育研究。E-mail:aying1999@foxmail.com。
马闯(1984-),男,讲师,博士,主要从事近自然林经营。E-mail:machuang_tjau@126.com。
责任编辑:杨霞
