丹参三倍体与二倍体叶片特征及抗旱性分析
2021-04-06刘婉秋庞俊秀曹艳楠王辉孙艳杰孙亚菲龙鸿
刘婉秋,庞俊秀,曹艳楠,王辉,孙艳杰,孙亚菲,龙鸿
丹参三倍体与二倍体叶片特征及抗旱性分析
刘婉秋,庞俊秀,曹艳楠,王辉,孙艳杰,孙亚菲,龙鸿通信作者
(天津农学院 园艺园林学院,天津 300392)
采用植物解剖学方法,对人工合成的丹参三倍体和丹参二倍体植株叶片解剖特征进行了观察,并比较分析了其抗旱性。结果显示,丹参三倍体植株成熟叶片表皮毛比丹参二倍体的长,叶片下表皮气孔长度、保卫细胞长度、保卫细胞厚度均大于丹参二倍体,气孔密度仅为二倍体的62.8%。此外,丹参三倍体叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度均明显增厚且栅海比比值更大,栅栏组织厚度可达到丹参二倍体的1.369倍。本研究表明,丹参三倍体比二倍体具有更强的光合作用能力和防止水分蒸腾的能力,抗旱性更强。本研究可为多倍体的抗旱适应性提供试验依据,为丹参三倍体在生产上的应用提供参考。
叶片解剖结构;抗旱性;三倍体;丹参
丹参(Bunge.)为唇形科鼠尾草属药用植物,是久负盛名的中药之一。中药丹参不仅具有祛除瘀肿、静心除躁、活血通络、保护肝脏等功效[1],还可有效治疗心血管疾病,广泛应用于临床冠心病、心肌梗塞和心绞痛等心血管疾病的预防和治疗,具有很好的疗效[2]。由于丹参大多以根或根茎入药,随着药用市场需求量的逐年递增,丹参已供不应求,致使国内外学者开始寻找新药源以缓解丹参根的药用市场压力。通过对丹参叶片所含物质的分析,发现其具有多种与根相似的成分,且其药理作用也与根基本相同[3],包括叶片在内的整株入药也可以在保证疗效的情况下满足更多市场需求。
多倍体植物是经过基因组染色体加倍形成的新物种,可人工合成或自然进化形成,就生物量而言,多倍体植株一般比二倍体植株高大、长势旺盛、生物量大[4]。对于以营养器官为主要利用目的的中草药,多倍体具有比二倍体更大的优势。除了器官巨大外,多倍体植物抗逆性往往较二倍体增强,例如小麦[5]、大豆[6]等植物的多倍体均具有比未加倍的物种有更强的抗逆性。丹参广泛分布于山东、山西、河南、四川、安徽、陕西等地,四川的丹参产量居全国首位[7]。目前,丹参多倍体还没有生产应用。本文以人工合成的丹参三倍体[8]为材料,探讨丹参三倍体叶片特征及其与抗旱性的可能关联,为丹参三倍体的抗逆性提供试验依据。
1 材料与方法
1.1 试验材料
丹参三倍体和二倍体材料由南开大学生命科学学院陈力教授馈赠,三倍体植物材料为组培苗,2018年4月经炼苗后移栽至天津农学院试验园,二倍体植物材料为正常实生苗,常规大田管理。待秋后10月移入实验室内,次年4月开花。选取开花前3月份相同生长期的丹参三倍体和二倍体材料,测量株高,在茎中段离地等高位置处,选取羽状复叶中间小叶测量叶长、叶宽,进行叶片表皮细胞观察与测定,经固定后做解剖观察。
1.2 试验方法
1.2.1 丹参形态学观察
经苗期生长成熟后至开花前,观察、测量、记录丹参二倍体和三倍体的形态学特征,包括:植株的叶长、叶宽、株高;利用Leica S8体式显微镜观察叶片表皮毛,分别测定30株植株。使用Microsoft Excel 2010版本对数据进行统计,利用SPSS软件(IBM SPSS Statistics 25)分析植株的叶长、叶宽、株高以及叶长/叶宽的测量数据。
1.2.2 丹参叶片表皮细胞观察与测定
用剪刀分别剪取丹参二倍体及三倍体完整健康的叶片,清水洗净后,采用直接撕取叶表皮的方法,先用刀片在叶表面轻划出一小方块,再用镊子分别撕取叶片的上、下表皮,将撕取的表皮放在滴有蒸馏水的载玻片上并使表皮展平,无重叠,加盖玻片后置于Leica DM 4000B显微镜观察,Leica DFC450 显微相机拍照。记录视野范围内气孔个数,随机选取10个气孔,Leica LAS软件测量植株的气孔长度、保卫细胞的长度及厚度,并对所测得的数据进行分析,计算气孔密度。
1.2.3 丹参叶片解剖结构的观察与测定
常规石蜡切片法制片,丹参二倍体及三倍体叶片采用FAA固定,梯度乙醇脱水,石蜡包埋后,Leica RM2235切片机切片,厚度8~10 µm,番红固绿复染,Leica DM 4000B显微镜观察,Leica DFC450 显微相机拍照。Leica LAS软件测量丹参的叶片厚度及叶片各组织层厚,进行数据分析。
2 结果与分析
2.1 丹参二倍体和三倍体植株形态
与二倍体植株相比,丹参三倍体在叶宽、叶长/叶宽和株高3个指标与二倍体差异显著,叶长差异不显著。三倍体植株更高,正常生长的三倍体植株株高最高可达45.26 cm,平均为37.04 cm,而二倍体植株株高最高仅为35.68 cm,平均为25.82 cm(表1)。与二倍体相比,三倍体叶片更为肥大,叶片较宽,叶长/叶宽值更小,即二倍体的叶片相比三倍体丹参更为细长(表1)。

表1 二、三倍体丹参植物形态学指标比较
注:叶长、叶宽、叶长/叶宽采用独立样本Mann- Whitney U检验,株高采用独立样本T-检验。单因素方差分析,同列数据后标注不同大写字母代表差异极显著(<0.01),不同小写字母代表差异显著(<0.05)。下同
2.2 丹参二倍体和三倍体叶片表皮细胞微形态
2.2.1表皮毛
丹参二倍体和三倍体植株成熟叶片上下表皮均覆盖浓密的表皮毛,观察结果显示,三倍体植株的叶片表皮毛长度为430.5 µm,二倍体植株的叶片表皮毛长度为250.6 µm,丹参三倍体叶片表皮毛比丹参二倍体的长(图1,表2),且表皮毛密度高(表2),表明丹参三倍体比二倍体防止水分蒸腾的能力更强,抗旱性也更强。
注:A. 丹参三倍体,B. 丹参二倍体。标尺=200 µm

表2 丹参二倍体及三倍体表皮毛长度和密度比较
2.2.2 气孔分布及数目
显微观察显示,丹参二倍体叶片上表皮可见少量气孔,而丹参三倍体叶片上表皮则未见明显气孔结构。丹参二倍体和三倍体叶片下表皮则可见气孔均匀分布于波浪状表皮细胞间,且两种倍性丹参叶片气孔保卫细胞形状无显著差异(图2)。
注:A. 丹参三倍体,B. 丹参二倍体。标尺=50 µm
相同视野范围内,丹参三倍体叶片下表皮气孔个数小于二倍体,且三倍体叶片的气孔密度较二倍体小(表3),仅为二倍体的62.8%。

表3 丹参二倍体及三倍体下表皮气孔密度比较
2.2.3 气孔长度及保卫细胞大小
丹参三倍体的气孔平均长度为14.949 μm,为二倍体的1.60倍;保卫细胞平均长度为26.982 μm,为二倍体的1.30倍;三倍体保卫细胞平均厚度为7.897 μm,为二倍体的1.18倍。三倍体叶片下表皮保卫细胞更大,其气孔长度、保卫细胞长度、保卫细胞厚度均大于丹参二倍体(表4)。

表4 丹参二倍体及三倍体下表皮气孔及保卫细胞比较 μm
2.3 丹参二倍体和三倍体叶肉的解剖结构
石蜡切片显示,丹参二倍体和三倍体叶片的解剖结构未见明显差异。其叶片横切面均由上表皮、栅栏组织、海绵组织、叶脉、下表皮组成(图3)。
注:A. 丹参三倍体,B. 丹参二倍体。标尺=100 µm
对丹参二倍体和三倍体叶片解剖组织进行测量,并计算各自栅海比,结果表明,丹参三倍体叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度及栅海比均大于二倍体,而下表皮厚度则小于二倍体,但表皮细胞厚度二者差异不显著。栅栏组织厚度所占叶片厚度的比例均为最高。三倍体的叶片厚度是二倍体的1.129倍,栅栏组织厚度是二倍体的1.369倍,海绵组织厚度是二倍体的1.239倍,上表皮厚度达到二倍体的1.128倍,而下表皮厚度则为二倍体的77.9%(表5)。丹参三倍体主叶脉直径、主脉导管直径、肋状突起高度均显著高于二倍体(表6,图4)。
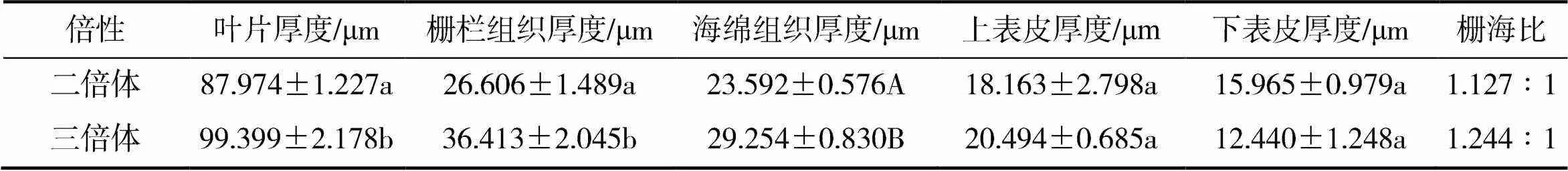
表5 丹参二倍体及三倍体叶片各组织层厚度比较

表6 丹参二倍体及三倍体叶脉比较 μm
注:A. 丹参三倍体,B. 丹参二倍体。标尺=100 µm
3 讨论与结论
植物在进化过程中,经历了漫长的自然选择,产生了广泛的适应性,其中多倍化就是一种适应性选择方式[9]。大多数多倍体具有更强的抗逆性。丹参多倍化后的适应性尤其是抗逆性研究具有品种推广的重要意义。
植物的抗旱性与植物体的生长状态有关。一般叶片小而厚,叶表面具有蜡质、叶片上下表面及茎上密被茸毛或鳞片都是植物的抗旱特征,是植物为了减少水分散失、提高水分利用效率的选择性适应[10]。研究表明,蒙农杂种冰草上、下表皮的表皮毛密度高、长度长,可以有效降低冰草在强光下的蒸腾作用,减少水分的蒸发[11]。叶片厚度是植物的抗旱特征之一,植物叶片越厚,保水能力越强,植株则越抗旱[12]。本研究中,丹参三倍体植株比二倍体植株高且叶片宽大,说明其生长能力较强;叶片表皮毛长度较长,说明防止蒸发、保水能力较强。丹参三倍体叶片下表皮的气孔长度、保卫细胞长度、保卫细胞厚度皆比丹参二倍体大,也说明其调节能力和保水能力更强。
从叶片解剖结构来看,叶片保水力与叶片的结构特点紧密相关[13-14]。徐扬等研究表明,川西高原地区4种苹果属植物叶片比较结果显示,4种植物叶片栅栏组织均由2~3层排列紧密的栅栏细胞组成,海绵组织细胞排列疏松,细胞间隙大,其中山荆子的栅栏组织厚度、栅海比和叶片结构紧密度最大,栅栏组织越发达,植株的抗旱性越强[15],抗旱性强的植物一般栅栏组织发达(2~3层),且栅栏组织/叶肉组织的比值大。贾梯研究证明,苹果叶片栅栏组织厚度的增加,对其叶片的光合速率以及其产量的提高具有重要作用[16]。此外,平欧杂种榛叶片的栅栏组织可有效减少叶片的水分蒸腾,提高植物的光合效能[17]。叶片的栅栏组织越厚,其表现出的抗旱保水的能力越强,而海绵组织越发达的植物,则表现出较差的抗旱性。栅海比也是评价叶片抗旱性的指标之一,栅海比越大,抗旱能力越强。本研究表明,相比丹参二倍体,丹参三倍体叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度均明显增厚,且栅海比比值更大,栅栏组织厚度可达到二倍体丹参的1.369倍。可见,丹参三倍体的光合作用能力更强且抗旱性优于丹参二倍体。
叶脉是植物叶片中的输导组织,具有水分和养分运输的功能,叶脉在植物叶片中的发达程度也与抗旱性相关。任媛媛等[18]、刘红茹等[19]研究表明,主脉中发达的维管束和强化的机械组织可使植株更加抗旱。蒙农杂种冰草主叶脉直径、主脉导管直径、肋状突起高度均显著高于中间冰草,分别为中间冰草的1.20、1.19、1.24倍,冰草叶片主叶脉直径、主脉导管直径、肋状突起高度等结构指标均与冰草的抗旱性存在密切关系。本研究中丹参三倍体主叶脉直径、主脉导管直径、肋状突起高度均显著高于丹参二倍体,说明丹参三倍体比二倍体具有更强的抗旱性。笔者认为,这些解剖结构上的变化可能与多倍体加倍后产生的器官巨大性相关,是多倍化之后适应环境能力增强的一种表现。
本研究初步得出以下结论:丹参三倍体植株成熟叶片表皮毛比丹参二倍体的长,叶片下表皮气孔长度、保卫细胞长度、保卫细胞厚度均大于丹参二倍体,气孔密度仅为二倍体的62.8%。丹参三倍体叶片厚度、栅栏组织厚度、海绵组织厚度、上表皮厚度均明显增厚且栅海比比值更大,栅栏组织厚度可达到丹参二倍体的1.369倍。丹参三倍体比二倍体具有更强的光合作用能力和防止水分蒸腾的能力,抗旱性更强,对环境的适应性更强。
[1] 袁媛,吴芹,石京山,等. 丹参及其主要成分保肝作用的研究进展[J]. 中国中药杂志,2015,40(4):588-593.
[2] 杨志霞,林谦,马利. 丹参对心血管疾病药理作用的文献研究[J]. 世界中西医结合杂志,2012,7(2):93-96.
[3] 薛治浦. 丹参叶抗氧化活性及相关酚酸类成分的研 究[D]. 郑州:河南科技大学,2011.
[4] ADAMS K L,WENDEL J F. Polyploidy and genome evolution in plants[J]. Curr Opin Plant Biol,2005,8:135-141.
[5] YANG C W,ZHAO L,ZHANG H K,et al. Evolution of physiological responses to salt stress in hexaploid wheat[J]. Proc Natl Acad Sci,2014,111(32):11882-11887.
[6] COATE J E,POWELL A F,OWENS T G,et al. Transgressive physiological and transcriptomic responses to light stress in allopolyploid(Leguminosae)[J]. Heredity,2013,110(2):160-170.
[7] 邓爱平,郭兰萍,詹志来,等. 丹参本草考证[J]. 中国中药杂志,2016,41(22):4274-4279.
[8] 李秀兰,陈力. 三倍体丹参的培育及其可持续利用研 究[J]. 中草药,2012,43(2):375-379.
[9] SCHIESSL S V,KATCHE E,IHIEN E,et al. The role of genomic structural variation in the genetic improvement of polyploidcrops[J]. Crop Journal,2018,7(2):127-140.
[10] SOUZA P U,SILVA LIMA L K,SOARES T L,et al. Biometric,physiological and anatomical responses ofspp. to controlled water deficit[J]. Scientia Horticulturae,2018,229:77-90.
[11] 李波,陈雪梅,李铁缘,等. 2种不同抗旱性冰草叶片解剖结构的比较[J]. 江苏农业科学,2015,43(9):247-249.
[12] 吴林,霍焰,聂小兰,等. 沙棘叶片组织结构观察及其与抗旱性关系的研究[J]. 吉林农业大学学报,2003,25(4):390-393.
[13] ESPERON-RODRIGUEZ M,CURRAN T J,CAMAC J S,et al. Correlation of drought traits and the predictability of osmotic potential at full leaf turgor in vegetation from New Zealand[J]. Austral Ecology,2018,43(4):397-408.
[14] PETROV P,PETROVA A,DIMITROV I,et al. Relationships between leaf morpho-anatomy,water status and cell membrane stability in leaves of wheat seedlings subjected to severe soil drought[J]. Journal of Agronomy and Crop Science,2018,204(3):219-227.
[15] 徐扬,陈小红,赵安玖. 川西高原4种苹果属植物叶片解剖结构与其抗旱性分析[J]. 西北植物学报,2015,35(11):2227-2234.
[16] 贾梯. 矮化砧苹果树叶片构造及光合作用的研究[J]. 北京农学院学报,1995,10(2):23-28.
[17] 李嘉诚,罗达,史彦江,等. 平欧杂种榛叶片解剖结构的抗旱性研究[J]. 西北植物学报,2019,39(3):462-471.
[18] 任媛媛,刘艳萍,王念,等. 9种屋顶绿化阔叶植物叶片解剖结构与抗旱性的关系[J]. 南京林业大学学报(自然科学版),2014,38(4):64-68.
[19] 刘红茹,冯永忠,王得祥,等. 延安5种木犀科园林植物叶片结构及抗旱性研究[J]. 西北农林科技大学学报(自然科学版),2013,41(2):75-81.
Leaf characteristics and drought resistance of triploid and diploid
Liu Wanqiu, Pang Junxiu, Cao Yannan, Wang Hui, Sun Yanjie, Sun Yafei, Long HongCorresponding Author
(College of Horticulture and Landscape, Tianjin Agricultural University, Tianjin 300392, China)
The plant anatomy method was used to observe the anatomical characteristicsof the leaves of artificialtriploid anddiploid plants, and their drought resistance was compared and analyzed. The results showed that the trichomes of mature leaves of triploid plants ofwas longer than that of diploid plants, and the stomatal length, guard cell length, and guard cell thickness of the abaxialtrichomes of the leaves were larger than those of the diploid ofwhile stomatal density was lower, 62.8% of the diploid. Moreover, the leaf thickness, the thickness of palisade tissue and spongy tissue, and the thickness of adaxial epidermis were significantly increased in triploid of, and the ratio of palisade tissue to spongy tissue was higher than that of diploid plants. The thickness of palisade tissue couldreach 1.369 times of that in diploid plants. These results indicate that triploid ofhas stronger photosynthesis ability and ability to prevent water transpiration than diploid, and has higher drought resistance. This study can provide an experimental basis for drought resistance adaptability of polyploids, and provide a reference for the application oftriploid in production.
leaf anatomical structure; drought resistance; triploid;
S567.53
A
1008-5394(2021)01-0001-05
10.19640/j.cnki.jtau.2021.01.001
2020-04-02
国家级大学生创新创业训练计划项目(201810061004)
刘婉秋(1997—),女,本科在读,林学专业。E-mail:1664883756@qq.com。
龙鸿(1964—),男,教授,博士,主要从事果树资源、果树生物技术研究。E-mail:longhong@tjau.edu.cn。
责任编辑:杨霞
