不同负荷游泳运动对大鼠海马食欲素A及其受体1表达的影响
2021-04-06张业廷魏翠兰袁琼嘉
张业廷,李 雪,付 燕,魏翠兰,袁琼嘉*
自主运动可以有效改善认知能力以及促进脑健康,强迫运动也可以起到类似的效果(Dishman,1997;Yuede et al.,2018)。运动可能会通过改变不同大脑脑区神经突触可塑性来影响学习和记忆能力,但具体机制有待进一步研究。海马是学习与记忆的重要脑区,其神经突触具有高度的结构可塑性和功能可塑性,在认知调控中起重要作用(Sun et al.,2018)。有研究表明,运动可以改善海马神经的突触可塑性,从而改善海马依赖性学习和记忆过程(Clark et al.,2008;Erickson et al.,2011;Hyodo et al.,2012)。但关于游泳运动对认知影响的相关研究较少,游泳运动对于大鼠而言属于典型的强迫运动形式。
虽然有研究表明,体育活动可以改善海马齿状回神经发生,但运动引起的海马神经可塑性的机制尚不清楚,尤其是强迫运动(Rhodes et al.,2003;Smith et al.,2005)。体育活动通过改变与海马突触可塑性相关分子的表达,包括脑源性神经营养因子(brain derived neurotrophic factor,BDNF)、神经细胞粘附分子(neural cell adhesion molecule,NCAM)等,可以引起神经生长,促进长时程增强效应(Long-term potentiation,LTP)过程和海马依赖性学习记忆(Erickson et al.,2011;Van Praag et al.,1999)。与自愿运动类似,强迫运动可以增强大鼠的空间学习和记忆形成以及海马BDNF表达(Khabour et al.,2013)。近年来,有研究表明,食欲素(Orexin)及其受体与海马的神经突触可塑性有关(Chieffi et al.,2017;Laperchia et al.,2017)。
食欲素是一种下丘脑神经肽,可与两种密切相关的G蛋白(OX1/2R)结合。食欲素有两种类型:食欲素A(Orexin A,OXA)和食欲素B(Orexin B,OXB),食欲素受体1(OX1R)是下丘脑外侧分泌的神经肽,OX1R与OXA高度相关,研究表明OX1R与学习记忆相关(Chieffi et al.,2017;Rashidy-Pour et al.,2015;Zhao et al.,2014)。虽然食欲素促进行动和认知过程的机制以及促进学习记忆的机制尚未被充分理解,甚至存在争议,但有研究表明,食欲素在调节高级大脑功能方面发挥着重要作用(Akbari et al.,2006,2007,2008;Jaeger et al.,2002;Li et al.,2017;Mavanji et al.,2017;Raoof et al.,2015;Selbach et al.,2010;Zhao et al.,2014)。如OXA被发现会导致年轻成年小鼠海马突触传递的长期增强(Li et al.,2017;Zhao et al.,2014)。研究表明,食欲素可以通过谷氨酸能、去甲肾上腺素能和胆碱能信号,在海马区引起突触可塑性(Chieffi et al.,2017;Selbach et al.,2004)。对于阿尔兹海默症转基因小鼠而言,其OXA表达低于野生型小鼠,而将OXA(1.5~3 nmol)注入其齿状回或蓝色黄斑核时,海马的LTP会显著增强(Jaeger et al.,2002)。对于正常大鼠而言,颅内注射OXA(0.14~0.56 nmol)能够显著增强大鼠的学习能力(Telegdy et al.,2002)。相反的是,有研究表明,正常大鼠颅内注射OXA(1~10 nmol)会抑制海马LTP,对大鼠在Morris水迷宫实验过程中表现的空间记忆能力造成损害(Aou et al.,2003)。大鼠海马神经元缺氧模型伴随着OXA表达的增加,而OXA可通过介导OX1R加重间歇性低氧血症引起的海马神经元损伤(Li et al.,2017)。上述研究中出现相互矛盾的结果,可能是由于研究的动物模型(正常或病理)、食欲素剂量/浓度差异所造成的。
目前关于食欲素及其受体在特定运动刺激下表达变化的研究较少,尤其缺乏强迫运动对其表达影响的研究。我们推测一定强度的游泳运动可以影响OXA和OX1R的表达,从而改变大鼠的空间学习和记忆。然而,不同运动负荷对认知的影响可能会有所不同(Dishman,1997;Yuede et al.,2018)。因此,本研究利用不同负荷游泳的运动方式对大鼠进行干预,观察游泳运动对大鼠空间学习和记忆能力的影响。同时测定OXA和OX1R的表达水平,探讨OXA和OX1R的表达水平与空间学习记忆能力的关系。
1 研究对象与方法
1.1 动物饲养
2月龄健康雄性SD大鼠30只,体重为300±20 g,购于成都达硕科技公司。分笼饲养于成都体育学院无特定病原体(SPF)动物实验中心。室内温度为22°C±2°C,室内湿度为45%~65%,采用12 h/12 h白天/黑夜循环环境喂养,自由进食与饮水。
1.2 动物分组及运动方案
大鼠适应性喂养第1周结束后,随机分为3组:对照组(C组)、中等负荷运动组(M组)、大负荷运动组(O组),每组n=10。C组大鼠自然喂养8周,M、O组大鼠进行为期8周的运动干预。游泳运动干预在第2周周一开始,游泳器具为透明玻璃缸(长150 cm,宽60 cm,深50 cm,水温34°C±2°C)。在游泳运动干预的第1天,M、O组大鼠游泳10 min,之后每天游泳时间递增5 min,直到第3周周五能够每天游泳60 min。然后,M组大鼠每天固定进行60 min的游泳运动干预,每周干预6天(周一至周六下午21:00—22:00),游泳运动干预至第9周。O组大鼠从第4周开始继续每天游泳时间递增10 min,至第4周周六每天游泳120 min,随后每天负重游泳,时间为120 min/天,从第5周开始,大鼠负自身体重游泳,从负自身体重1%至负自身体重5%,每周增加1%,直至第9周负自身体重5%,每周同样干预6天(周一至周六下午21:00—22:00)(崔笑梅 等,2017;袁琼嘉 等,2009;Elikov,2016)。
1.3 Morris水迷宫实验
Morris水迷宫实验(MWM)于第10周开始,用于评估大鼠空间学习与记忆能力,包括连续7天的定位航行实验和60 s空间探索实验(D’Hooge et al.,2001)。在定位航行实验中,每天将每只大鼠依次从4个象限中点轻轻放入水中,让其寻找水下2 cm处固定于第Ⅳ象限的黑色圆形平台。寻找水下平台的时间最长为120 s,若超时还未完成,则将大鼠引导至水下平台并在平台上保持15 s,同样的,在120 s内能够寻找并站在平台上的大鼠也需保持15 s后再捞起。大鼠从入水到找到平台的时间即为逃避潜伏期,结合大鼠游泳速度可以分析大鼠的空间学习能力。定位航行实验结束24 h后,拆除水下平台,进行空间探索实验。空间探索实验时,将大鼠从第Ⅱ象限轻轻入水,计时60 s大鼠在第Ⅳ象限中所用的时间及穿过原来水下平台所在位置的次数可以用来评价大鼠的空间记忆能力。
1.4 样本采集
MWM实验结束24 h后,断头处死大鼠,将脑组织迅速从颅骨中取出。各组取4例全脑进行免疫荧光检测。将各组剩余6例全脑迅速取出其海马组织,用于Western Blot和Real-time PCR实验。
1.5 Real-Time PCR实验
严格按照 RNAzol®RTRNA Isolation Reagent(GeneCopoeiaTM,USA)试剂盒说明提取海马组织mRNA。逆转录酶将mRNA样本反转录成互补的DNA(cDNA)。随后以cDNA为模板,进行Real-Time PCR实验,反应条件见表1。分析目的基因表达采用2-△△CT(Livak)方法。由Ta-KaRa宝生物工程有限公司设计所需引物,其序列见表2。

表2 PCR引物序列及扩增产物长度Table 2 PCR Primer Sequence andAmplification Product Length
1.6 免疫荧光实验
通过免疫荧光实验检测OXA蛋白表达量。将全脑组织进行冰冻切片,室温下放置8 min,随后在4°C环境下于4%多聚甲醛中固定10 min。用PBS清洗2次后,于4°C环境下孵育OXA一抗(1:50,R&D Systems USA)过夜。第 2天 PBS清洗后,用荧光抗体(1:100,R&D Systems USA)37°C环境下孵育1 h。采用DAPI进行核染色。利用OLYMPUS显微镜获得荧光图像,使用Image-Pro Plus 6.0软件分析荧光强度(AU/μm2)。
1.7 Western Blot实验
通过Western Blot实验检测海马组织中OX1R蛋白表达量。用湿转方法将蛋白质转至PVDF膜上。剪取目的蛋白条带,用去脂牛奶封闭1 h,之后将带有目的蛋白的PVDF膜用一抗在4°C冰箱中孵育过夜。第2天,用TBST液洗涤PVDF膜后用二抗室温孵育1 h;洗涤后进行化学发光、显影、定影,获得目的蛋白印迹图像。对胶片进行扫描,利用Gel-pro Plus软件对目的蛋白和内参蛋白作光密度(OD)值分析。
1.8 数据分析
所有实验数据由SPSS统计软件进行处理,以均值±标准差(M±SD)表示。各组间相关数据分析采用单因素方差分析法,若方差齐性,则用LSD法进行事后检验;若方差不齐,则将原始数据转换至齐性后再做统计处理。以P<0.05表示数据具有显著性差异。
2 结果
2.1 大鼠空间学习与记忆能力
在MWM定位航行实验过程中,各组大鼠的逃避潜伏期均逐渐减少(图1A),表明各组大鼠都能够有效的学习寻找水下平台的技能。每天各组大鼠之间的游泳速度无显著差异(P>0.05,图1B),表明逃避潜伏期未受到游泳速度的影响。M组大鼠在第3天平均逃避潜伏期显著低于C组及O组(P<0.05),而C组与O组大鼠在第5天平均逃避潜伏期最短,并且之后各组表现再没有显著提升,且C组与O组之间平均逃避潜伏期并无显著差异(P>0.05,图1A)。在MWM定位航行实验中,M组穿越原平台所在区域的次数显著高于C组及O组,O组与C组相比有减少的趋势,但并无显著差异(P<0.05,P<0.01,P>0.05,图1C)。
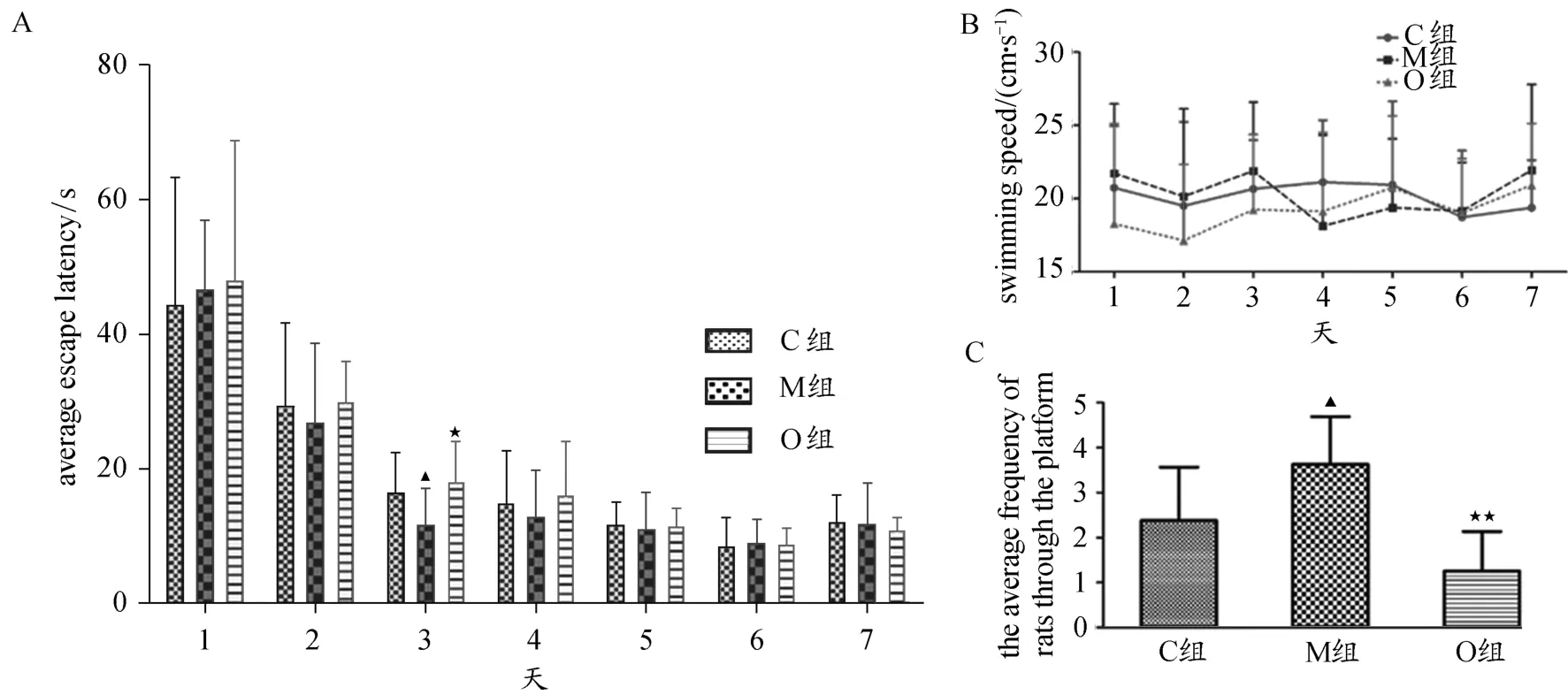
图1 Morris水迷宫实验大鼠空间学习与记忆能力Figure 1.Spatial Learning and Memory of Rats in Morris Water Maze Experiment
2.2 海马OXAmRNA及OX1R mRNA含量
所有检测基因均在200 bp以下,且无引物二聚体出现,结合它们的扩增曲线和溶解曲线,可以判断产物是目的基因。M组大鼠OXA mRNA表达量显著高于C组(P<0.05,图2A),但其OX1R mRNA表达量显著低于C组(P<0.05,图2B)。而C组与O组大鼠OXA mRNA及OX1R mRNA表达量无显著差异(P>0.05,图2)。由此看出,OXA mRNA与OX1R mRNA的表达量并不同步。
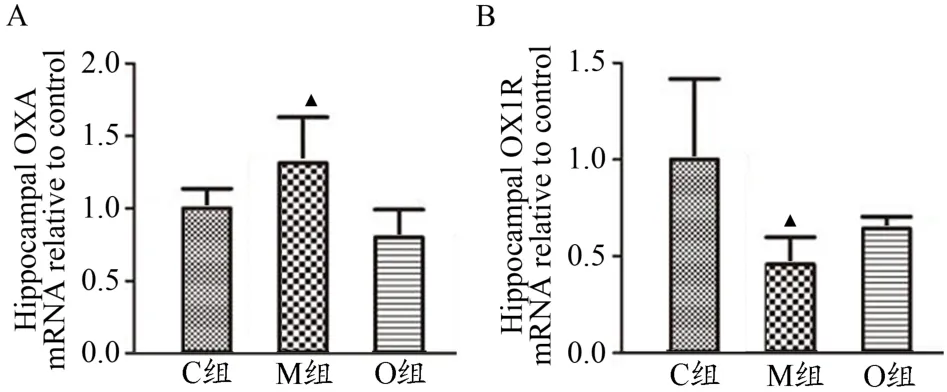
图2 海马OXAmRNA及OX1R mRNA表达情况Figure 2.Expression of OXAmRNAand OX1R mRNA in the Hippocampus
2.3 海马OXA蛋白及OX1R蛋白含量
激光共聚焦显微镜下观察到海马区OXA免疫阳性产物的分布。与C组相比,M组大鼠的免疫阳性产物荧光强度在海马DG、CA3、CA1 3个区域较低,说明M组大鼠OXA蛋白表达水平低于C组;O组大鼠免疫阳性产物荧光强度在海马DG、CA3、CA1 3个区域与C组无显著差异,说明O组大鼠OXA蛋白表达水平与C组无显著差异(P<0.05,P>0.05,图3A~I,图4A~C)。

图3 OXA免疫荧光检测结果Figure 3. OXA Immunofluorescence Assay

图4 蛋白表达水平Figure 4. Protein Expression Level
Western Blot分析显示,M组大鼠OX1R蛋白表达水平显著低于C组(P<0.05,图4D)。C组、O组大鼠比较无统计学差异(P>0.05,图4D)。因此,中等负荷运动训练后大鼠OXA和OX1R蛋白表达水平均明显下降,但大负荷运动训练对OXA和OX1R蛋白表达无显著影响。
3 讨论
适宜负荷的运动可以改善健康水平,但过高或过低的运动负荷可能会损害健康。这一点在心血管疾病患者中得到了最显著的证明,如果想要降低获得心血管疾病的风险,则不能低于体育活动负荷的阈值(Garber et al.,2011)。关于不同负荷运动对认知影响的研究还较少,有研究指出,较大的运动负荷可能会损害认知,但也是研究则表明,较大的运动负荷反而有助于改善认知(Baird et al.,2018;Moore et al.,2016)。在不同负荷游泳运动结束后,本研究对大鼠进行MWM实验,结果表明,中等负荷游泳运动可以改善大鼠的空间学习和记忆,大负荷游泳运动不能显著影响大鼠空间学习记忆能力。虽然如此,但在本研究中大负荷游泳运动表现出具有损害大鼠空间学习记忆能力的趋势,之所以未出现显著差异,可能与本研究的样本量较少及干预周期较短有关。同时,我们在进行游泳干预过程中发现,在大负荷游泳运动时,大鼠呛水的概率似乎会提高,这有可能会增加大鼠的应激性。在后续研究中我们将对游泳运动过程中除去运动以外的其他刺激,如呛水、水温等的应激作用展开进一步的研究,并进一步增加游泳运动干预的周期及大鼠样本量,以取得更加可靠的实验结果。
普遍认为,突触是神经元网络和信息传递的关键结构,神经元突触可塑性是神经生物学中学习记忆的重要影响因素(Edelmann et al.,2014)。LTP是突触可塑性的重要内容,被广泛认为是构成学习和记忆基础的主要分子机制之一。食欲素及其受体(OX1R和OX2R)广泛分布于大脑中,在调节学习和记忆方面发挥着重要作用。OXA可以调节多个大脑区域和行为系统,并通过作用于氨基丁酸能、谷氨酸能、肾上腺素能和胆碱能的海马神经元,特异性地参与学习和记忆的认知过程(Simth et al.,2005)。近年来,动物研究发现,OX1R和OX2R通过参与乙酰胆碱、组胺、去甲肾上腺素、血清素和多巴胺的调节,在调节睡眠觉醒和促进学习记忆方面发挥作用(Aou et al.,2003;Ni et al.,2014;Tsujino et al.,2009;Zhao et al.,2014)。OXA和OX1R在海马中的表达变化影响突触的可塑性,与调节学习记忆的过程密切相关(Aou et al.,2003;Selbach et al.,2010;Zhao et al.,2014)。OXA通过增强海马突触的可塑性和cAMP反应元件结合蛋白的磷酸化,促进了社会识别记忆(Lang et al.,2013)。另有研究表明,OXA可以促进学习和记忆的获得、巩固及恢复过程(Akbari et al.,2008)。但以上研究并未提及OXA具体的表达量与学习记忆能力之间的关系。
本研究表明,中等负荷游泳运动能够降低大鼠海马OXA和OX1R蛋白的表达水平,并且可以改善大鼠的空间学习与记忆能力。值得一提的是,M组大鼠OXA mRNA表达量显著高于C组,而OXA表达量显著低于C组。转录水平和蛋白表达水平出现了矛盾,可能是由于OXA蛋白的形成需要经过转录、翻译、翻译后修饰等过程,OXA mRNA属于转录环节,而翻译及翻译后修饰的过程存在很大的变数,机制复杂。本研究未能对其具体机制进行研究,在未来的研究中,我们将会尝试对这一现象进行分析。有研究指出,过度激活食欲素系统会导致神经元损伤,进而导致学习和记忆能力下降(Aou et al.,2003,Ni et al.,2014)。研究表明,原发性失眠症患者的记忆能力下降与OXA水平升高有关,高水平的OXA可引起失眠,并通过过度唤醒大脑皮层而损害记忆功能(Rayner,2003)。OXA可以双向调节海马CA1突触可塑性,在OXA表达水平较高时,提高OXA表达水平可以通过OX1R抑制LTP;在OXA表达水平较低时,提高OXA表达水平可以通过OX1R和OX2R诱导再增强LTP(Lu et al.,2016)。
通过以上研究可以看出,某些病症可能会导致OXA表达超过(如原发性失眠症等)或低于(如阿尔兹海默症等)正常的生理水平,从而可能会抑制海马LTP,损害学习与记忆能力;而OXA表达正常时,增高或降低OXA都有可能对学习和记忆能力起到促进作用。因此推测,OXA表达在正常的生理水平内时,可以双向调节海马CA1突触可塑性。OXA表达在正常的生理水平内时,当OXA表达水平超过某一阈值,降低OXA表达水平,有助于改善学习与记忆能力;当OXA表达水平低于某一阈值,提高OXA表达水平,有助于改善学习与记忆能力。目前还不能确定OXA表达的这一阈值。本研究中,所采用的2月龄健康雄性SD大鼠,OXA表达可能超出了这一阈值,因此,中等负荷游泳运动可能通过降低OXA表达水平,改善了大鼠的学习与记忆能力。另外,本研究显示,大负荷游泳运动没有显著改变OXA和OX1R蛋白的表达水平。
综上,中等负荷游泳运动可能通过降低OXA和OX1R表达水平,改变了大鼠海马的LTP,这也许是中等负荷游泳运动改善大鼠空间学习记忆能力的机制之一。
4 结论
中等负荷游泳运动能提高大鼠的空间学习记忆能力,这可能与中等负荷运动能够降低大鼠海马OXA和OX1R的表达水平有关。大负荷游泳运动不能显著影响大鼠海马OXA和OX1R的表达水平及其空间学习记忆能力。
