大花黄牡丹花粉萌发及贮存特性*
2021-04-02贾文庆王艳丽郭英姿王政齐庆闫三妮刘会超何松林
贾文庆王艳丽郭英姿王政齐庆闫三妮刘会超何松林
(1.河南科技学院 河南省园艺植物资源利用与种质创新工程研究中心 新乡 453003;2.河南农业大学风景园林与艺术学院 郑州 450002; 3.洛阳国家牡丹园 洛阳 471011)
大花黄牡丹(Paeonialudlowii)为芍药科(Paeoniaceae)芍药属(Paeonia)牡丹组(Sect.Moutan)肉质花盘亚组(Subsect.Delavayanae)落叶大灌木,为西藏特有植物,自然仅在林芝、米林、隆子县等有零星分布,生长势强,高达2~3.5 m,是牡丹最高大的种质,花鲜黄色,是金黄色牡丹品种来源的主要亲本,秋叶红色,观赏价值极高; 其根、花瓣入药,为名贵的藏药之一(汪松等,2004)。大花黄牡丹多数仅有1个心皮,结实率少,加之生境干扰严重和人为盗掘毁坏等原因,致使其野生种群数量大幅减少,处于濒危状态,现被列为国家二级保护植物(李嘉珏等,1998; 洪德元等,1999; 汪松等,2004; 杨小林等,2007)。芍药属野生种质资源中仅有黄牡丹(Paeoniadelavayivar.lutea)、大花黄牡丹具有黄色的基因,栽培牡丹除紫斑牡丹(Paeoniarockii)、杨山牡丹(Paeoniaostii)外,大多数种类植株低矮、生长纤弱,黄色品种少,严重制约了牡丹的应用推广。因此,研究分析大花黄牡丹花粉的贮存特性,对于开展芍药属植物的种间杂交育种工作,培育具有自主知识产权的株型高大、黄色系牡丹品种具有重要意义。
牡丹现已确认的9种野生种质及居群分布在华北、西北、西南、华中等地区,区域跨度大,花期存在较大差异,杂交需要贮存花粉,花粉低温或超低温贮存能够延长花粉的寿命,是随时随地为授粉杂交提供足量可育花粉的有效方法,检测花粉萌发率是远缘杂交前的基础性工作,花粉离体萌发技术是评价花粉萌发率以及花粉是否贮存成功的可靠手段(Akhondetal., 2000; Shekarietal.,2016; Sorkhehetal.,2018; Lietal.,2019)。近年来,越来越多的研究发现,花粉贮存期间的萌发率与活性氧、自由基的清除机制及膜质过氧化程度有关(谭健晖,2011; 刘艳萍等,2013; 贾文庆等,2015; Liuetal.,2020),氧化应激引起的细胞损伤甚至细胞凋亡可能是贮存花粉萌发率下降的主要原因之一(石印等,2015; 徐瑾等,2015; Renetal.,2020)。以往关于大花黄牡丹的研究主要集中在分类学、生态学(苏建荣等,2010; 杨翔等,2010)、遗传学(林启冰等,2004; 唐琴等,2012)、孢粉学(何丽霞等,2005; 曾秀丽等,2009)等方面,芍药属不同牡丹种质花粉萌发适宜的培养基差异较大(盖伟玲等,2010; 律春燕等,2010; 贾文庆等,2012; 施江等,2013),王士泉等(2012)报道了大花黄牡丹花粉的育性情况,但仅限于用孔雀绿-醋酸洋红测定花粉生活力,而关于培养基成分对花粉萌发,不同贮存温度对花粉寿命及细胞内生理指标的影响,尚未见报道。因此,本试验以大花黄牡丹隆子县居群花粉为试验材料,对其花粉萌发特性、低温下花粉的贮存特性进行研究,并测定贮存期间抗坏血酸(AsA)含量等生理指标的变化,旨在找出大花黄牡丹花粉短期、中期和长期贮存的适宜温度,并明确贮存期间不同温度下大花黄牡丹花粉萌发率降低的生理机制,为进一步开展大花黄牡丹的杂交育种工作提供参考。
1 材料与方法
1.1 试验材料
大花黄牡丹花粉采自西藏隆子县,2018年5月上中旬9:00—11:00,采集含苞待放的大花黄牡丹花朵,迅速放入冰盒,在实验室将花萼和花瓣剥去,去除花柄,然后倒置放入培养皿中,置于(22±1) ℃培养箱内散粉,24 h后取出花朵,左手用镊子夹住倒置的花朵,右手用镊子轻弹,收集花药开裂散出及掉落的花粉。将收集的花粉分成2份,一份直接用于花粉萌发的测定; 另一份30 ℃鼓风干燥箱干燥24 h后(含水量降至6.5%±0.5%),分装至2 mL带盖离心管中盖口贮存,备用。
1.2 花粉萌发试验及花粉扫描电镜观察
在前期牡丹花粉萌发研究(贾文庆等,2020)基础上,采用L9(34)正交试验设计,一个对照(表1),对硼酸、蔗糖、GA3、CaCl2进行4因素3水平的正交试验,每培养基(5 g·L-1琼脂,pH6.0)处理重复3次。培养基配制后,分装至35 mm一次性培养皿中,冷却后,将1.1收集的新鲜花粉放入1 mL蒸馏水中,充分搅拌制成悬浮液,然后用滴管将悬浮液移至培养基上,用涂布棒涂布均匀,然后放入盛有少量水的大号培养皿里,加盖。置于25 ℃下恒温培养18 h后,在光学显微镜下统计花粉萌发率,每样品观察200粒以上,以花粉管长度大于花粉粒直径作为花粉萌发标准。花粉饱满率观察方法如下:将1.1收集的花粉酒精梯度脱水后,将花粉均匀撒在粘有双面导电胶的圆形电极板上,喷金后在日立SU-1510扫描电镜下观察花粉并进行拍照,统计饱满的花粉比率。
1.3 不同贮存温度对花粉萌发和花粉寿命的影响试验
将1.1所得的大花黄牡丹干燥花粉分成5份,每份分装20~30个2 mL带盖离心管(将花粉装入放有1~3粒变色硅胶的离心管,每管1.2 g左右花粉,封口贮存),分别放在室温(地下室阴凉干燥处,18~25 ℃)、4 ℃、-20 ℃、-80 ℃、-196 ℃(放入液氮罐中,定期添加液氮)下贮存。贮存第24、40、72、120、184、264、365天后,分别从5种温度中取1~2只离心管,取少许花粉用2.1节最佳培养基检测萌发率(冷冻下花粉经35 ℃水浴快速化冻),确定5种温度下的花粉寿命(花粉寿命=花药散粉到萌发率降低到50%时所经历的天数),其余花粉用于保护酶活性等生理指标测定。
1.4 不同贮存温度对丙二醛(MDA)和抗坏血酸(AsA)含量的影响试验
MDA含量采用北京索莱宝科技有限公司提供的试剂盒测定:称取约0.100 g花粉,加入1 mL提取液进行冰浴匀浆; 8 000×g4 ℃离心 10 min,取上清,置冰上待测。混合液在 100 ℃水浴中保温60 min后(盖紧,防止水分散失),置于冰浴中冷却,10 000×g常温离心10 min。取上清至1 mL玻璃比色皿中,测定各样品在450 nm、532 nm和600 nm处的吸光度,换算出MDA含量。
AsA含量测定成分提取方法与MDA相同,反应体系参照李合生等(2000)的方法,采用分光光度计测定525 nm处的吸光度值,根据制作的标准曲线确定抗坏血酸的含量。
1.5 不同贮存温度对3种保护酶活性的影响试验
酶液的提取参照贾文庆等(2020)方法。超氧化物岐化酶(SOD)活性采用氮蓝四唑法测定,过氧化物酶(POD)活性采用愈创木酚法测定,过氧化氢酶(CAT)活性采用可见光分光光度法测定(李合生等,2000)。
1.6 数据分析
采用EXCEL2016处理数据和作图,并利用SPSS22.0进行方差分析、多重比较。
2 结果与分析
2.1 大花黄牡丹花粉萌发适宜培养基的筛选及花粉饱满率观察
不同正交组合处理培养基对大花黄牡丹花粉萌发率影响的研究结果(表1)表明,不同培养基条件下花粉萌发率存在显著差异(P<0.01),其中处理5萌发率最高,平均达92.10%,对照无任何添加物的培养基萌发率最低,平均仅为23.50%,这表明大花黄牡丹花粉虽然能够借助于自身积累的养分萌发,但萌发率较低。采用EXCEL2016 对花粉萌发率数据进行直观分析,蔗糖、硼酸、CaCl2和GA34因素的极差(R)分别为23.78、19.86、6.78、6.34,可以确定对花粉萌发的诱导效应表现为蔗糖>硼酸> CaCl2> GA3。根据分析结果,确定大花黄牡丹花粉萌发的最优组合为120 g·L-1蔗糖+45 mg·L-1硼酸+55 mg·L-1GA3+30 mg·L-1CaCl2,其萌发率显著高于其他组合。统计观察发现,大花黄牡丹隆子县居群单株开花3~8朵(图1A),秋叶猩红(图1B),每朵花的花粉量平均干质量在(180±32) mg,果实多为单心皮蓇葖果(图1C)。扫描电镜结果显示,畸形花粉有勺形、三角形、不规则形等,总量较少,仅占观察花粉总数的5.6%,花粉饱满率高达94.4%,饱满率与萌发率基本符合(图1D、E)。

表1 不同培养基对大花黄牡丹花粉萌发率的影响①Tab.1 Effects of different medium on pollen germination of Paeonia ludlowii

图1 大花黄牡丹植株状态及花粉饱满率Fig. 1 Plant status and pollen plumpness rate of Paeonia ludlowiiA.开花状态; B.示红叶; C. 示单心皮; D, E. 畸形花粉。A. Flowering state; B. Red leaf; C. A follicle with only one carpel; D, E. Deformed pollen.
2.2 不同贮存温度对花粉寿命的影响
花粉贮存是人工保存种质资源的重要手段。不同贮存温度和处理时间对花粉萌发率的影响差异较大(P<0.01)(图2),除-196 ℃外,其他4个贮存温度下,大花黄牡丹花粉萌发率均随时间延长而下降。室温下贮存花粉萌发率随时间延长迅速降低,说明越来越多的花粉快速丧失发芽力,贮存24天花粉萌发率47%,为贮存前新鲜花粉萌发率的50.87%; 贮存第72天时,萌发率降为0,花粉死亡,说明冷凉干燥下花粉贮存寿命为24天。4 ℃贮存72天花粉萌发率为49%,120天时萌发率降为15%,这说明4 ℃下花粉寿命大概为80天左右,第264天时花粉萌发率降为0。-20 ℃、-80 ℃和-196 ℃贮存下花粉萌发率下降趋势不同。-20 ℃下0~365天时,花粉萌发率呈快速下降趋势,贮存第120天,花粉萌发率55%,贮存第184 天,萌发率降为32%,说明-20 ℃下花粉寿命120~184天; -80 ℃下贮存,花粉萌发率缓慢下降,而-196 ℃贮存下萌发率曲线保持平缓,贮存第365天,-80 ℃花粉萌发率为62.10%,达到贮存前花粉萌发率的66.23%,-196 ℃贮存下花粉萌发率为92.1%,为贮存前萌发率的99.67%,这表明-80 ℃和-196 ℃花粉寿命均大于1年,-196 ℃更适合花粉的长期贮存,而-80 ℃适合隔年杂交的花粉贮存。综上所述,大花黄牡丹花粉贮存的最佳温度为-196 ℃。
2.3 不同贮存温度及时间对大花黄牡丹花粉MDA含量的影响
MDA含量是花粉细胞膜质过氧化的重要指标,大花黄牡丹花粉在不同贮存环境中MDA含量变化如图3所示。室温、4 ℃贮存,花粉MDA含量先升高而后快速降低,室温下首先达到峰值,4 ℃下随后出现高峰; 室温下MDA含量变化相对较大,贮存72天后,升高到24 μmol·g-1(FW),为贮存前的5.21倍; 4 ℃贮存184天,MDA含量峰值达20.5 μmol·g-1(FW); 随后,2个贮存温度下MDA含量迅速下降,推测可能是花粉细胞凋亡,MDA分解造成的。-20 ℃下MDA则呈现逐渐升高的趋势,贮存第365天时MDA含量达到贮存前的4.49倍,说明花粉细胞膜质过氧化严重; 而-80 ℃条件下,大花黄牡丹花粉MDA含量呈缓慢上升的趋势,贮存第365天MDA含量升高到7.8 μmol·g-1(FW),为贮存前的1.69倍; -196 ℃下,大花黄牡丹花粉MDA含量总体稳定,贮存365天后,MDA含量与贮存前无显著差异,这表明花粉细胞在-196 ℃贮存下并未造成膜质过氧化,花粉萌发率高。
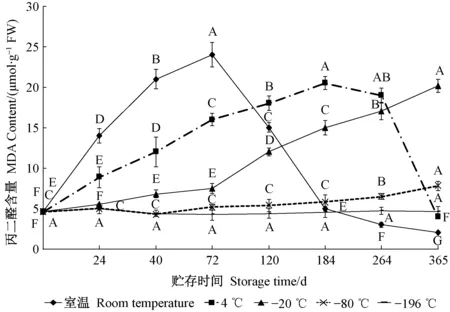
图3 不同贮存温度及时间对大花黄牡丹花粉丙二醛含量的影响Fig. 3 Effects of different storage temperature and time on MDA content in pollen
2.4 不同贮存温度及时间对大花黄牡丹花粉SOD活性的影响
不同贮存温度及处理时间对SOD活性有显著影响(P<0.01)(图4A)。室温下贮存,大花黄牡丹花粉SOD活性随时间延长总体呈迅速下降趋势,SOD活性的降低可能是花粉死亡较多,只有少部分花粉具有SOD活性造成的。4 ℃和-20 ℃下贮存,SOD活性升高,分别在第24天、第72天达到峰值,此后随着贮存时间的延长,SOD活性迅速下降,4 ℃下贮存第264天花粉萌发率降为0,SOD活性仅为3 U·g-1(FW),-20 ℃下贮存第365天,SOD活性仅为最高峰值的23.90%,这表明,此时SOD已不足以抵抗花粉膜质过氧化导致的伤害,花粉萌发率也出现了大幅下降。-80 ℃贮存条件下,SOD活性呈先缓慢上升随后下降再逐渐上升的趋势,但总体变化幅度比较平缓,贮存第365天,SOD活性达到贮存前的1.65倍,这表明大花黄牡丹花粉清除活性氧的能力强,花粉仍具有较高活性。而-196 ℃液氮贮存条件下,SOD活性基本保持稳定,贮存期间SOD活性无显著变化,花粉萌发率与新鲜花粉无显著差异,SOD活性124 U·g-1(FW),为贮存前的1.07倍,这表明大花黄牡丹花粉细胞内代谢稳定,没有受到伤害或伤害较小,因此花粉萌发率高。
2.5 不同贮存温度及时间对大花黄牡丹花粉POD活性的影响
大花黄牡丹花粉经过不同贮存温度和时间处理后,POD活性变化显著(P<0.01)(图4B)。花粉POD活性在室温、4 ℃、-20 ℃下,先升高而后降低。室温下第24天,POD活性升至59 U·g-1(FW),达到最高,为原来的2.32倍,之后随着时间的增长,大花黄牡丹花粉自身的保护能力迅速下降,POD合成受到抑制,至贮存第264天,降为0; 4 ℃贮存,大花黄牡丹花粉的POD酶活性最高峰值出现在第72天,之后随着时间增长,POD酶活性开始迅速下降,到第264天时,活性仅为贮存前活性的8%; -20 ℃,POD活性最高峰值出现在第120天,之后伴随着花粉萌发率的下降而迅速下降,贮存365天,POD酶活性仅为3.5 U·g-1(FW)。-80 ℃贮存,POD酶活性同样呈先升高后下降的趋势,最高峰出现在第184天,之后下降。-196 ℃贮存条件下,POD酶活性整体波动较小,趋势平缓,365天时活性是原来的1.04倍,这表明超低温下,花粉内部代谢处于平衡状态,自由基产生较少,低POD酶活性协同其他保护酶及其他成分等能够及时分解自由基,保证花粉细胞内部代谢稳定,因而花粉活力较高。
2.6 不同贮存温度及时间对大花黄牡丹花粉CAT活性的影响
不同贮存温度对CAT活性的影响显著(P<0.01)(图4C),室温、4 ℃、-20 ℃贮存条件下,大花黄牡丹花粉的CAT活性首先升高,分别在第24天、第40天、第120天达到峰值,之后随着贮存时间的延长,室温、4 ℃下花粉CAT活性迅速下降,分别在贮存第264天和第365天降为0; -20 ℃下,在第120天花粉CAT活性达到峰值后快速下降,贮存第365天,花粉CAT活性仅为24 mg·g-1(FW)。-80 ℃贮存温度下,随着贮存时间的延长,大花黄牡丹花粉的CAT活性总体上呈起伏不定的曲线,第365天最高峰值达91 mg·g-1(FW),为原来的1.25倍; -196 ℃贮存温度下,CAT活性呈现震荡下降再上升而后下降的趋势,但幅度极为平缓,与贮存前无显著差异。这表明随着时间的推移,花粉可以在低温下分解自由基,从而减少损害,保持较高的生存能力。

图4 不同贮存温度及时间对大花黄牡丹花粉酶活性的影响Fig. 4 Effects of different storage temperature and time on enzymatic activity in pollen
2.7 不同贮存温度及时间对大花黄牡丹花粉AsA含量的影响
AsA是细胞体内重要的抗氧化剂,由图5可以看出,大花黄牡丹花粉经过不同贮存温度和时间处理后,AsA含量变化显著不同(P<0.01)。室温、4 ℃、-20 ℃、-80 ℃贮存条件下,大花黄牡丹花粉AsA含量随着时间的延长,呈现先升高后迅速下降的趋势,室温、4 ℃、-20 ℃、-80 ℃的高峰值分别出现在第24、72、120、184天,之后除-80 ℃贮存温度外,伴随着花粉萌发率的下降,花粉AsA含量均迅速下降; 而-196 ℃贮存温度下,花粉AsA含量先出现震荡随后呈略为上升的趋势,365天时AsA含量为3.25 mg·g-1(FW),仅为原来含量的1.19倍,这表明此时花粉可以清除保护酶不能清除的超氧阴离子自由基、羟自由基等自由基,进而保护花粉减少低温伤害,保持较高的花粉萌发率。

图5 不同贮存温度及时间对大花黄牡丹花粉AsA含量的影响Fig. 5 Effects of different storage temperature and time on AsA content in pollen
2.8 大花黄牡丹花粉萌发率与生理指标的相关性
由相关性分析结果(表2)可知,大花黄牡丹花粉萌发率与MDA含量呈显著负相关关系,与AsA含量、SOD活性、POD活性、CAT活性呈显著正相关(P<0.01),SOD活性与花粉萌发率相关系数达0.816 5,这表明大花黄牡丹花粉在5个贮存温度下,SOD活性是影响花粉萌发、花粉寿命的最主要因子,其他主要影响因子依次是CAT活性、AsA含量和POD活性; 膜质过氧化是导致花粉凋亡的主要因素。表2表明,MDA含量与SOD呈显著负相关,与POD、CAT、AsA呈负相关,SOD、POD、CAT、AsA含量相互呈显著正相关,这说明大花黄牡丹花粉中3种保护酶和AsA含量协同作用较强,花粉SOD可以有效清除活性氧过多积累造成的花粉细胞膜质过氧化。
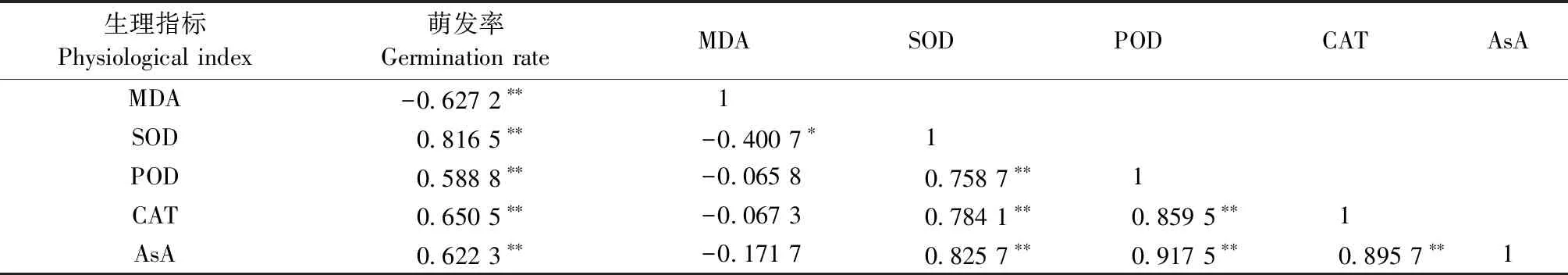
表2 花粉萌发率、MDA、AsA含量以及3种保护酶活性的相关分析①Tab.2 Correlation between pollen germination, protective enzymes activity, MDA, and AsA content
3 讨论
3.1 不同培养基及不同贮存方法对花粉萌发的影响
蔗糖、H3BO3、Ca2+和GA3是花粉萌发率离体检测最常用的添加物 (Feietal.,2003; Gokbayraketal.,2017; Flores-Renteriaetal.,2018)。其中,蔗糖不仅为花粉的萌发和花粉管的生长提供了必要的能量,而且在一定程度上维持了外界环境的渗透压,保证了花粉的萌发 (Hiroseetal.,2014); H3BO3可以增加花粉细胞对糖的摄取、转运和代谢,诱导Ca2+从外界进入细胞,建立花粉管顶端生长所需的Ca2+梯度; Ca2+在花粉管极性生长和生长方向的调节中具有重要的作用(Renetal.,2020); GA3对花粉萌发和花粉管生长有重要作用,低浓度GA3可显著提高杏(Prunusarmeniaca)花粉萌发率(Feietal.,2003; Gokbayraketal.,2017; Flores-Renteriaetal.,2018)。植物基因型不同,适宜的花粉检测培养基亦不同,芍药属植物花粉萌发适宜的蔗糖浓度为50~150 g·L-1,硼酸浓度30~100 mg·L-1(李秉玲等,2010; 律春燕等,2010; 贾文庆等,2012; 施江等,2013)。本试验结果也证实了这一点,蔗糖、H3BO3、Ca2+、GA3不同组合对大花黄牡丹花粉萌发有显著影响,萌发适宜的蔗糖浓度达120 g·L-1,高于矮牡丹(Paeoniajishanensis)的90 g·L-1(贾文庆等,2012),低于黄牡丹的150 g·L-1(律春燕等,2010),这表明不同牡丹野生种质花粉适宜的培养基不同。花粉萌发率与饱满率具有相关性(贾文庆等,2012; 王士泉等,2012),本试验发现,大花黄牡丹隆子县居群开花多、花粉量大,花粉畸形率较低,平均仅5.6%,结合萌发率(可育花粉率)90%以上来分析,二者具有关联,饱满率高可能是大花黄牡丹新鲜花粉萌发率较高的原因,这与成仿云等(1997)研究发现种子是大花黄牡丹唯一的繁殖途径是一致的,表明大花黄牡丹具有较强的有性生殖能力。
贮存后生物材料生存力的变化是用来决定贮存成功与否的重要指标,不同牡丹种质花期不同,自然情况下牡丹花粉寿命仅有3~15天,因此杂交需要贮存花粉,合适的贮存环境对于大花黄牡丹花粉延长寿命进而通过杂交选育优良新品种具有重要意义。研究表明,芍药属植物花粉寿命的长短受多方面因素的制约。一是受其基因型的控制,基因型不同,花粉寿命差异明显(李秉玲等,2010; 贾文庆等,2012; 石印等,2015); 二是花粉含水量是影响花粉寿命的主要因素之一,低含水量是延长花粉寿命的有效手段,含水量6%~13%的芍药属种质花粉贮存寿命长(施江等,2009; 李秉玲等,2010; 石印等,2015)。此外,贮存的环境极为重要,低温、干燥和低氧环境可有效延长花粉寿命,在应用较多的室温干燥、低温干燥、冷冻干燥贮存和超低温贮存中,4 ℃贮存芍药属种质花粉寿命为20~60天,-20 ℃贮存条件下花粉寿命为30~120天,而在-80 ℃、-196 ℃贮存环境中,矮牡丹、日本牡丹‘岛大臣’(Paeoniasuffruticosa‘Shimadaijin’ )、牡丹‘凤丹白’(Paeoniaostii‘Fengdan Bai’)等花粉寿命均超过1年,尤以-196 ℃液氮贮存条件下萌发率最高(施江等,2009; 李秉玲等,2010; 贾文庆等,2012; 石印等,2015)。本研究也证实了这一点,室温干燥下,大花黄牡丹花粉寿命仅有24天,而4 ℃、-20 ℃环境花粉寿命在3~4个月,分析可能是较高的温度下花粉生理代谢较强,造成大量花粉发芽力丧失; -80 ℃、-196 ℃贮存1年的花粉萌发率分别达新鲜花粉的66.23%、99.67%,这表明大花黄牡丹花粉在这2种温度下花粉寿命均超过1年。以上结果表明,低温特别是超低温是大花黄牡丹花粉长期贮存的最佳环境,可能是低温降低了花粉的生理代谢,进而延缓衰老延长了花粉寿命。综上所述,-80 ℃由于经济适用,适宜大花黄牡丹跨年杂交花粉的贮存,而-196 ℃适合大花黄牡丹花粉的长期贮存。
3.2 花粉萌发率与MDA、AsA含量及保护酶的关系
植物体内自由基、活性氧的产生与清除是平衡的,在正常生长条件下自由基、活性氧水平较低,低水平的自由基、活性氧不会造成损伤,且有一定的积极作用(Kanazawaetal.,2000)。在逆境或程序性死亡过程中,细胞内活性氧、自由基种类数量大幅度增多,活性氧产生与清除的平衡被打破,损害生物膜及功能,植物一般通过提高抗氧化酶活性、合成AsA等抗氧化物质启动防御系统,减少或避免对细胞功能的伤害(Liuetal.,2020; Renetal.,2020)。MDA是植物细胞膜系统受害、细胞凋亡的主要标志物,测定MDA可以推断贮存环境对花粉细胞是否造成伤害及伤害的程度(Dongetal.,2018; Zafraetal.,2018)。SOD在控制超氧阴离子的积累以防止膜脂质和蛋白质的氧化损伤中起着至关重要的作用,但SOD升高也增加了细胞中H2O2的浓度,而CAT、POD和其他抗氧化酶可以平衡H2O2的含量(Renetal.,2020),且POD能消除酚、胺、醛、苯等的毒性 (Liuetal.,2020)。此外,某些引起植物氧化损伤的剧毒活性氧成分,例如不能被SOD、POD、CAT等抗氧化酶清除的超氧阴离子和羟自由基,也是引起细胞氧化损伤的主要因素,因此植物只能依靠非酶类物质去除这些活性氧成分(Boseetal.,2014),AsA是非酶抗氧化剂系统中的关键成分,是最有效的抗氧化剂之一,AsA不仅与H2O2反应,而且与超氧阴离子自由基、羟自由基和脂质过氧化物反应,从而维持活性氧的平衡状态,保护细胞膜并参与其他抗氧化剂的再生(Renetal.,2020)。
室温、4 ℃贮存条件下,大花黄牡丹花粉萌发率、MDA含量、AsA含量、3种保护酶活性的变化规律相似,随着贮存时间的延长,花粉萌发率快速降低,MDA含量、AsA含量、3种保护酶活性基本呈升高然后迅速降低的趋势,这可能是因为贮存初期,在外界温度和水分的胁迫下,花粉代谢紊乱,自由基增加,MDA增加膜质过氧化加剧,为了维持代谢平衡,花粉细胞SOD、POD、CAT活性增加,合成AsA,协同花粉内的脯氨酸、维生素E等清除过多的自由基、活性氧,以维持细胞膜的稳定性。但随着贮存时间的延长,活性氧、自由基的过度积累导致细胞损伤严重,没有过多的保护酶、AsA来防止损伤,出现氧化应激,细胞受到损伤,花粉逐渐衰亡,因此花粉萌发率迅速下降。室温下,SOD活性逐渐下降,POD活性在第24天,CAT活性、AsA含量在第40天达到高峰值,MDA含量第72天达到峰值; 而在4 ℃条件下,3种保护酶、AsA含量在第24天、第72天达到峰值,MDA含量则第184天才达到峰值,较室温推迟112天,与SOD、CAT相比,POD、AsA含量的峰值推迟,在-20 ℃和-80 ℃时更明显。这表明,与室温相比,4 ℃下花粉活性氧积累较慢,膜系统伤害来得较迟,MDA含量达到峰值后下降可能是细胞凋亡后,MDA分解造成的; POD、AsA含量的峰值推迟可能是因为贮存时花粉细胞不仅产生SOD、CAT可以清除的活性氧,还可能产生其他有毒物质,如超氧阴离子自由基、羟自由基、胺或酚类物质(Fecht-Christoffersetal.,2006; Liuetal.,2020)。
-20 ℃贮存下,贮存初期(第0~40天)大花黄牡丹花粉萌发率下降缓慢,SOD、POD活性及AsA含量持续增加,而CAT活性则在下降后缓慢上升,MDA含量则缓慢上升,可能是在这一时期花粉通过协调3种类型的保护酶、AsA含量来维持自由基的产生和消除之间的平衡,从而维持花粉发芽力。随着贮存时间的延长,花粉萌发率明显下降,MDA含量持续上升,SOD活性在第72天出现高峰后迅速下降,而POD、CAT活性及AsA含量则在120天达到高峰随后下降,这表明第72天到120天,花粉中活性氧种类及其他有毒物质如酚类或胺类物质在增加,花粉通过不断增加POD、CAT活性及AsA含量消除这些有毒物质,从而减缓花粉活性的丧失(Fecht-Christoffersetal.,2006)。-20 ℃贮存第365天后,花粉萌发率仅为贮存前萌发率的12.9%,分析可能是因为在大多数花粉中,自由基、活性氧积累过多,膜质过氧化严重,花粉细胞损伤加剧,导致花粉无法萌发,只有一小部分花粉能够产生保护酶、AsA来维持活性(贾文庆等,2015)。谭健晖(2011)的研究表明,在-20 ℃贮存358天后,马尾松(Pinusmassoniana)的花粉仍然保持较高的萌发率(42.20%)和较强的抗氧化能力。这表明由于遗传因素的不同,不同植物花粉的萌发和贮存特性不同。
-80 ℃贮存的早期(第0~72天),大花黄牡丹花粉通过增加SOD活性、AsA含量来维持内部代谢平衡,花粉萌发率缓慢下降; 贮存第72~184天,MDA含量持续增加,SOD活性不断上升,CAT活性在下降后又上升,POD活性、AsA含量则持续上升,分别在第120、184天达到高峰,这表明此阶段活性氧持续增加,需要协调3种保护酶及AsA含量清除有毒物质维持花粉活性。贮存后期(184~365天),花粉中的活性氧逐渐积累,MDA含量逐渐增加显示膜质过氧化逐渐加重,导致花粉萌发率逐渐下降,为了消除细胞中过多的活性氧,细胞SOD活性升高,在此期间,细胞内的其他有毒物质可能逐渐减少,所以细胞只需要增加CAT活性来清除H2O2,因此,POD活性、AsA含量缓慢下降,而CAT活性逐渐增加。因此,在花粉贮存过程中,影响花粉萌发率的不仅是活性氧,还有其他有毒物质,具体还需要进一步的研究。综合来看,大花黄牡丹-80 ℃贮存期间,花粉3种保护酶、AsA含量、MDA含量总体变化幅度较-20 ℃、4 ℃、室温下小,这可能是大花黄牡丹花粉贮存1年后萌发率仍保持在较高水平的原因。
许多植物花粉在超低温贮存时出现“冷刺激”现象,日本牡丹、矮牡丹等芍药属植物花粉也有类似发现(李秉玲等,2010; 贾文庆等,2012),其发生原因可能与贮存后花粉细胞内Ca2+增加、存在差异蛋白有关(李秉玲等,2010)。本研究发现,-196 ℃下大花黄牡丹花粉贮存前期(第0~40天),花粉并非处于生理停滞状态,同-80 ℃一样,贮存第40天,花粉萌发率升高,出现“冷刺激”现象,较贮存前新鲜花粉分别提高1%和1.5%,伴随着花粉萌发率的提高,花粉MDA含量降低,而SOD、POD、CAT、AsA含量升高,这表明超低温下大花黄牡丹花粉细胞自由基、活性氧较少,膜质过氧化程度低,花粉细胞健康程度提高,推测这可能是大花黄牡丹花粉萌发率“冷刺激”现象发生的生理原因,具体有待进一步研究。贮存中后期(第40~365天),MDA含量处于较低的水平,一直保持稳定,而SOD、CAT、POD及AsA同样保持稳定,贮存过程中无显著差异,这表明-196 ℃下贮存1年的过程中,花粉细胞内SOD、CAT、POD及AsA协同作用,有效清除了多余的活性氧和其他有毒物质,处于代谢平衡状态,未造成严重的膜质过氧化,这是大花黄牡丹花粉贮存后萌发率仍较高的原因。
前人研究表明,马尾松、银杏(Ginkgobiloba)、大花红山茶(Camelliamagniflora)、芍药(Paeonialactiflora)等植物花粉的保护酶活性、MDA和AsA含量与萌发率具有相关性(谭健晖,2011; 刘艳萍等,2013; 贾文庆等,2015; 石印等,2015)。花粉萌发率与5个生理指标的相关分析表明,大花黄牡丹花粉在贮存过程中,SOD活性是影响花粉萌发、花粉寿命的最主要因子,其次的因子依次为CAT活性、AsA含量和POD活性; 膜质过氧化是导致花粉凋亡的主要因素。这些结果表明SOD在大花黄牡丹花粉贮存过程可以有效清除活性氧,防止或减轻膜质过氧化程度过高引起的细胞凋亡。本试验结果表明,室温下随着贮存时间的延长,大花黄牡丹花粉POD活性首先快速升高后降低,随着花粉萌发率的迅速降低,SOD活性逐渐降低,CAT则缓慢升高后降低,之后随着贮存时间的延长3种保护酶活性均迅速降低,说明POD在室温下较为敏感。4 ℃下,SOD和CAT在第24天同时出现峰值,但2种酶的抗氧化能力有限,胁迫的加重POD活性升高,AsA含量增加,在第72天达到峰值,之后迅速降低,与此同时,花粉萌发率迅速下降,说明花粉SOD和CAT在4 ℃下较为敏感。-20 ℃、-80 ℃下,花粉SOD活性则首先快速升高,说明SOD较为敏感。综上所述,POD在大花黄牡丹花粉室温贮存时为敏感的保护酶,SOD在-20 ℃和-80 ℃贮存时为敏感的保护酶,而SOD和CAT在4 ℃贮存时为敏感的保护酶,这与本试验的相关分析一致。
4 结论
大花黄牡丹自然环境下花粉畸形率低,花粉的萌发率与饱满率具有相关性; 适宜大花黄牡丹花粉萌发率检测的培养基为120 g·L-1蔗糖+45 mg·L-1硼酸+55 mg·L-1GA3+30 mg·L-1CaCl2; 花粉贮存温度直接影响大花黄牡丹的花粉萌发率,室温适合大花黄牡丹花粉1~24天的短期贮存,4 ℃、-20 ℃适合杂交时间间隔在80~120天大花黄牡丹花粉的中期贮存,-80 ℃适合花粉的跨年贮存,而-196 ℃适合大花黄牡丹种质花粉的长期贮存; 相关性分析结果显示,SOD活性是影响贮存期间大花黄牡丹花粉萌发、花粉寿命的最主要因子,其他因子依次为CAT活性、AsA含量和POD活性,膜质过氧化是导致花粉死亡的主要因素。花粉细胞内代谢处于动态平衡状态、细胞膜系统稳定是-196 ℃下大花黄牡丹花粉保持高萌发率的生理响应; 室温、4 ℃、-20 ℃、-80 ℃贮存期间,花粉保护酶活性、抗坏血酸含量显著升高后迅速降低,清除活性氧、自由基能力下降,活性氧、自由基积累过多,膜质过氧化程度加剧,细胞损伤严重是贮存期间花粉萌发率下降的主要原因。
