基于角尺度的随机体在森林稳定性维持中的作用*
2021-04-02惠刚盈赵中华张弓乔胡艳波
惠刚盈 赵中华 张弓乔 胡艳波
(中国林业科学研究院林业研究所 国家林业和草原局林木培育重点实验室 北京 100091)
森林群落稳定性维持机制是森林生态学研究的热点和核心问题。稳定性概念源于热力学理论,指对于外界干扰某个系统表现出的维持现状与恢复原状的潜力(Hurdetal.,1971),这里所说的群落稳定性是指群落在达到演替顶极后出现的能够进行自我更新并维持其结构和功能长期保持在一个较高水平、波动较小的现象。顶极群落是生物与环境在长期相互作用演变过程中相互适应和协调统一的产物,具有维持其结构和功能相对不变的稳定性。
一个多世纪以来,已有众多关于群落稳定性维持的理论先后被提出和验证。MacArthur(1955)提出了著名的多样性-稳定性假说,认为群落多样性和食物网复杂性会促进生态系统稳定性,群落多样性可为群落稳定性提供强有力的保障后盾,在一个群落中物种越多,其抵抗外界干扰的能力越强,恢复能力也越强。多样性-稳定性关系的实质是物种冗余所致,由此产生了稳定性的冗余(redundancy)理论(Lawtonetal.,1994; 党承林,1998),该理论来源于自动控制系统可靠性理论。在生态学中,Odum(1983)定义的冗余指超过1种以上的物种或成分具有执行相同特定功能的能力。Walker(1992)最初提出冗余种的概念是针对评价优先保护地区或优先保护物种的,认为冗余程度低的物种,一旦灭绝,对稳定性的影响十分明显,同时建议在确定冗余种时使用“功能群”概念,相同功能群的种可以在功能上相互替代,也就是冗余效应,当某一功能群仅剩单个物种时,此物种若从系统中损失将会导致系统功能遭受无法挽回的破坏,冗余种被认为是维持一定稳定性的“必要的”保险(Walker, 1995; Naeem,1998)。Janzen(1970)和Connell (1971)认为宿主专一性天敌在森林中具有随距宿主距离增加而降低的不均匀分布特点,导致靠近同种母体周围的更新体更易受天敌危害而具有较高的死亡率(距离制约性死亡)。这就是著名的Janzen-Connell假说:首先,母树周围的同种幼苗增补受到抑制,为其他物种定植提供空间,从而促进森林群落中物种共存; 其次,寄主高密度地点能够吸引天敌并有利于天敌的繁殖、传播,导致更新体具有较高的死亡率(密度制约性死亡)。在这一背景下,出现了另一个重要的生态学假说,即物种异群保护假说(herd immunity hypothesis),该假说认为异种邻体能够阻碍寄主专一的有害生物传播,目标个体周围异种邻体增加会提高其存活率。Ford(1975)、Weiner等(1984)证明了同种负密度制约和异群保护假说如何影响种群的自然发展,认为许多群落自然发展的过程是,期初林地被一些先锋树种侵占,根据同种负密度制约假说,先锋树种的优势木周围会排除同种个体,进而被其他树种代替,而这些先锋树种周围的其他树种常为小树。Pommerening等(2017)和Wang等(2018)指出大树个体周围常具有更高的混交度,并称之为混交度-大小假说(Pommereningetal.,2020),其本质是从相邻木关系角度诠释天然混交林特征,进一步印证了前人的假说。此外,物种间竞争可通过生态位分化来降低竞争紧张度而使更多物种共存。群落中的物种之所以能够长期稳定共存,是因为不同物种分别占据不同生态位,生态位互补性使不同树种共享地上和地下资源(Huietal.,2014; Belloetal.,2019),避免或延缓物种间的竞争排除作用,生态位相同或重叠程度较大的物种会因竞争有限的资源而发生竞争排除,不能稳定共存(牛克昌等,2009)。
上述假说的某些方面在试验中得到了证实,虽然许多生态过程仍然未知,但已暗示在稳定性研究中需要关注同种负密度制约、异群保护、生态位互补性以及物种多样性和冗余效应等方面。天然林是森林生态系统的主体,其结构基本上是物竞天择而形成的进化且健康的结构,具有较高的生物多样性、较复杂的群落结构、较丰富的生境特征和较好的生态系统稳定性,在保障农牧业生产、维持生物多样性、保护生态环境、减缓自然灾害、调节全球碳平衡和生物地球化学循环等方面发挥着极其重要和不可替代的作用(唐守正等,2000)。本研究试图精细解译天然林空间结构并挖掘其普适性规律来诠释森林稳定性维持机制,以期为改善我国人工林稳定性提供科学参考。
1 数据
本研究采用我国不同纬度带的11块天然林长期定位试验数据(表1)。样地详细情况参见文献Zhang等(2018)。
2 方法
本研究利用林分空间结构参数——角尺度进行林木格局解析(惠刚盈等,2003)。为描述相邻木围绕参照树的均匀性,惠刚盈等(1999)、Hui等(2002)提出了角尺度(Wi)概念。角尺度有5种取值,分别对应参照树相邻木的分布形式(图1)。根据参照树及其最近4株相邻木构成的结构单元中相邻木的分布形式定义为结构体,任意一个结构单元均由5 株林木即1株参照树与其4株最近相邻木组成。角尺度Wi=0或0.25的林木称为邻体均匀分布的参照树,简称均匀木,相应的结构单元称为均匀体; 角尺度Wi=0.75或1.00的林木称为邻体聚集或团状分布的参照树,简称聚集木,相应的结构单元称为聚集体; 角尺度Wi=0.50的林木称为邻体随机分布的参照树,简称随机木,相应的结构单元称为随机体(Zhangetal.,2018)。据此,任何一个森林群落最多由3种结构体组成,即均匀体、聚集体和随机体(惠刚盈,2020)。

表1 样地及其林分概况Tab.1 General situation of sample plots and stands
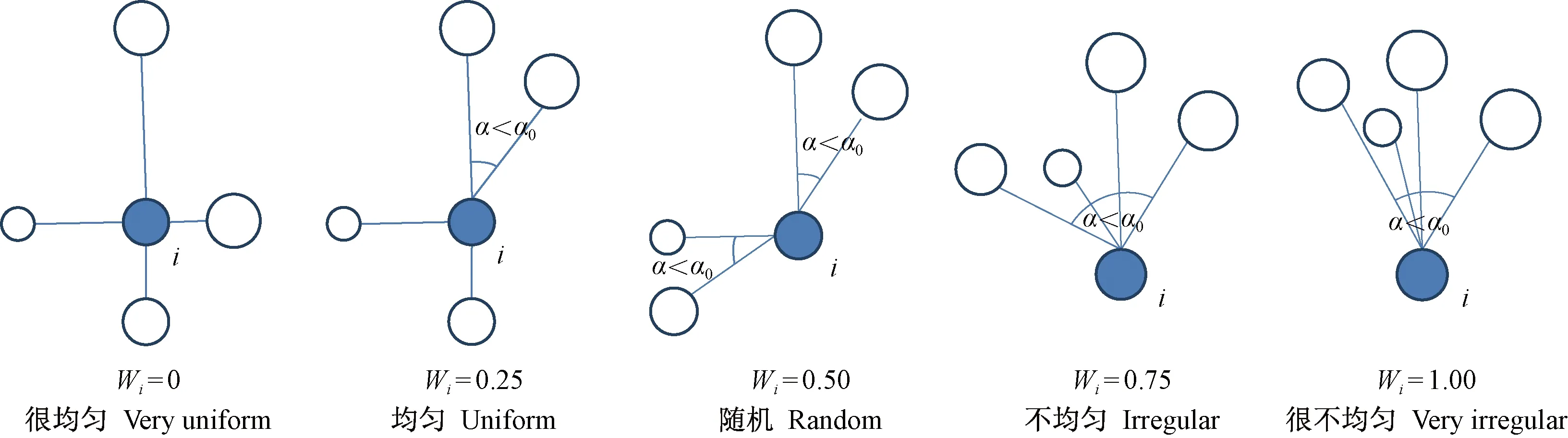
图1 角尺度的取值及其意义Fig. 1 The value and significance of uniform angle index
运用R软件编程,统计3种结构体的株数频率及其断面积占比。为消除边缘效应,每块样地均设置5 m缓冲区,处于缓冲区的相邻最近树仅作为潜在的邻体,不参与统计。
3 结果与分析
3.1 均匀木、随机木和聚集木的株数占比
统计不同角尺度取值的林木数量,获得林分角尺度分布,见图2。由图2可知,11块天然林样地林木角尺度分布呈现出一致规律:1) 林木角尺度基本为钟型分布(近似正态分布),峰值均出现在Wi=0.50,即随机木占多数; 11块样地中参照树邻体随机分布的林木均占半数以上,比例最高的甚至达62%左右(C4、C6和C9),最低的也可达55%以上(C7和C8); 2)Wi=0和Wi=1.00,即非常均匀和非常聚集的林木都很少,其中非常均匀分布情况最不常见,在所有试验样地中最高的也只有1%,有5块样地中的比例均为0; 非常聚集分布次之,占全部林木的10%以内; 3)Wi=0.25(均匀)和Wi=0.75(不均匀)的林木一般在10%~30%之间。
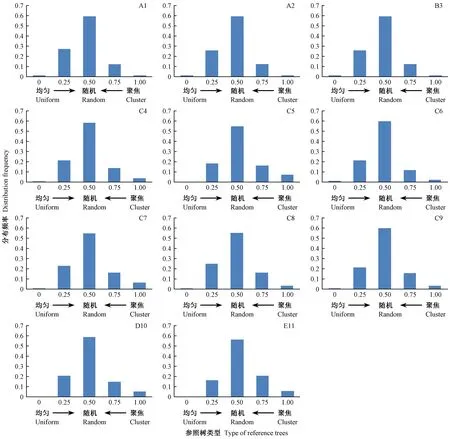
图2 天然林样地的角尺度分布Fig. 2 Uniform angle index distribution of natural forest sample plots
3.2 均匀木、随机木和聚集木的断面积占比
林木株数比例表达结构体分布的数量特征,而断面积占比在一定程度上可体现结构体分布的质量特征。分别统计均匀木、随机木和聚集木断面积占林分总断面积的比例,得到不同结构体的断面积分布见图3。由图3可知,林分中随机木断面积远高于均匀木和聚集木,在所有样地中均占林分总断面积的50%以上,最大可达62%左右(C6和C9); 而均匀木和聚集木断面积分别占林分总断面积的10%~30%以内,2类林木断面积总量基本小于林分总断面积的半数。
3.3 结构体形成机制分析
基于角尺度揭示的天然林中随机木、均匀木和聚集木的分布规律,既与天然林地域分布和森林类型无关,也与天然林树种组成和格局类型无关。天然林林分中主要群体为随机分布的林木个体(随机木),这一现象出现必有其内在的形成机制。
对于聚集体来说,相邻木拥挤在一起,使得中心木三面受光,可获得更多的营养空间、更大的树冠,甚至更高的生产力。相邻大树与小树之间的不平等将导致非对称竞争,尤其在相邻木相对拥挤时,竞争加剧不仅导致小径木生长放缓,而且相对弱势的相邻木可能在自然选择过程中死亡,此时聚集体瓦解,原结构体内剩余林木将与其他林木重新组合构成新的结构体,林木越聚集,越容易发生瓦解和重组(图4上)。而均匀体则相反,分布于均匀木四周的相邻木分别占据3~4个方位,从而使中心木至少受到来自3个方向的挤压或遮挡,造成中心木承受更大的竞争压力,如果均匀木处于相对劣势,则更容易在自然选择过程中被淘汰,此时均匀结构体瓦解,原结构体内剩余林木将与其他林木重新结合成为新的结构体,林木越拥挤,越容易发生瓦解和接踵而来的重组(图4下)。
不同于聚集体和均匀体,随机体中心木(随机木)具有两面受光的可能性,相较于均匀木,参照树竞争压力小,而随机木的邻体之间挤压程度较聚集体更小,因此随机木及其邻体相对来说承受的生存压力较小,这种结构体更不容易由于竞争缘故而导致弱势或不健康的林木频出,比其他2种结构体中的林木有更大的可能性稳定生长,从而在自然演替过程中获得更多的存活几率。
4 讨论
基于4株相邻木空间关系的林分空间结构参数——角尺度,将森林群落中的林木分为随机木、均匀木和聚集木,依次对应5株树组成的随机体、均匀体和聚集体(Zhangetal.,2018; 惠刚盈,2020),这一基于角尺度的森林结构解译拓宽了森林培育新视野。目前,已有许多基于角尺度进行林木分布格局的研究(Aguirreetal.,2003; 安慧君,2003; 惠刚盈等,2003; 夏富才,2007; 赵中华等,2009),其中,最有代表性的是惠刚盈等(2003)利用Winkelmass软件模拟产生了1 000个林木格局随机分布的林分,发现格局随机的林分角尺度分布为正态,其角尺度均值为0.496±0.007。随后,许多有关森林群落林木分布格局的研究均证实天然林林分平均角尺度接近0.50(胡艳波等,2003; 赵中华等,2009; 李纪亮,2008; 郑丽凤等,2009; 赵洋毅等,2012; 彭辉,2017)。近年来,随着研究不断深入,基于角尺度的林分空间结构分析不仅仅局限于用其均值进行格局判断,更重要的是格局形成机制的探索,愈加重视角尺度分布的研究。基于角尺度分布的天然林格局研究发现,处于随机分布(Wi=0.50)的林木(随机木)数量(株数或断面积)最多,占比在50%以上(Zhangetal.,2018; 陈科屹,2018; 万盼,2018),本研究印证了该结果。惠刚盈(2020)进一步从森林群落重要特征如树种组成(多度分布)、林分结构(直径分布、格局分布、混交度分布、密集度分布)和林木竞争状态(竞争分布)入手,对比分析了天然林整体与天然林群落中随机木的状态特征,结果发现二者具有高度相似性。可见,随机木是天然林的核心,在群落中起主体作用。在人工林空间结构与天然林对比研究方面,最有代表性的是曹小玉等(2020)对中亚热带典型林分空间结构的分析,其在立地条件基本一致的人工针叶纯林[10杉木(Cunninghamialanceolata)、10马尾松(Pinusmassoniana)、10柳杉(Cryptomeriafortunei)]、人工阔叶纯林[10木荷(Schimasuperba)]、人工阔叶混交林[7木荷+3闽楠(Phoebebournei)]、人工针阔混交林[7杉木+3木荷; 6马尾松+4木荷; 7柳杉+3马褂木(Liriodendronchinense)]和天然次生林[7青冈栎(Cyclobalanopsisglauca)+2苦槠(Castanopsissclerophylla)+1冬青(Ilexchinensis)]中共设置27块30 m×20 m样地,每种类型各重复3次,对林分平均角尺度的研究结果人工针叶纯林为0.35±0.025、人工阔叶纯林为0.28±0.03、人工阔叶混交林为0.40±0.02、人工针阔混交林为0.39±0.02、天然次生林为0.49±0.02,进一步证实人工林格局分布的均匀性和天然林格局分布的随机性。人工林分布格局的均匀性也表现在角尺度分布的左偏性上(袁士云,2010; 惠刚盈,2012; 岳永杰,2015)。人工林造林之初,如果严格按照相等株行距进行规则造林,根据角尺度定义可知,其分布格局必然如图5(惠刚盈,2020; 曹小玉等,2020; 岳永杰,2008; 冯宜明等,2018)中幼龄林阶段的分布格局,绝大多数林木角尺度为0,林分平均角尺度接近0; 随着林龄增加和林木生长,人工林郁闭,林木发生自疏,一些林木死亡,人工林林木角尺度出现0.25、0.50甚至是0.75的情况,林分角尺度分布逐渐趋向于倒“J”形。此后,随着人工林林龄进一步增加和林分自疏或多次经营干扰,林分中林木角尺度为0.25、0.50、0.75或1.00的林木越来越多,人工林角尺度分布也呈现出左偏状态,角尺度均值进一步增大(图5中龄林和成熟林阶段),虽在短期内还达不到像天然次生林一样的角尺度分布(图6)(Zhangetal.,2018; 陈科屹等,2018; 万盼,2018; 胡艳波等,2003; 赵中华等,2009; 李纪亮,2008; 郑丽凤等,2009; 赵洋毅等,2012; 彭辉,2017),但随着进展演替,其随机成分一定能达到像天然林一样的50%以上,只是时间问题。也就是说,人工林随着林分生长发育阶段演进,林分稳定性在增加的同时,角尺度分布将发生均匀木数量减少而随机木数量增多这样一个显著的变化过程(图5)。
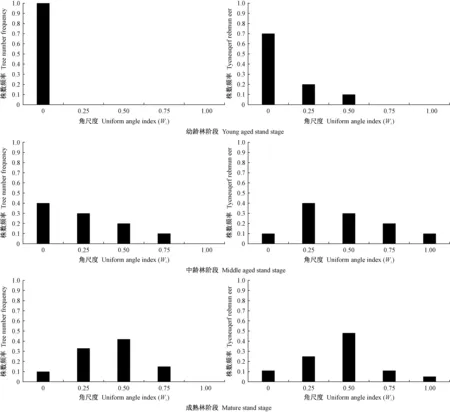
图5 人工林角尺度随时间的变化过程Fig. 5 Change process of uniform angle index of plantation with time
现有的森林稳定性维持机制主要关注组成森林群落的物种生态特性,对人工林培育的启示只能是营造树种生态位不同的多优树种混交林。本研究提出的基于空间结构的随机体-稳定性假说,对人工林培育的启示是随机化造林,考虑到生产可行性,提倡控位造林(惠刚盈,2020)。人工林林木分布格局的规则性使其本身失去了自然属性,而天然林林木分布格局的随机性则促使其更加稳定。现有人工林不稳定的根源除了树种组成单一外,主要是空间结构尤其是林木分布格局的均匀性所致,因此,提高现有人工林稳定性的关键是增加随机体,采用随机化经营(惠刚盈,2020)。
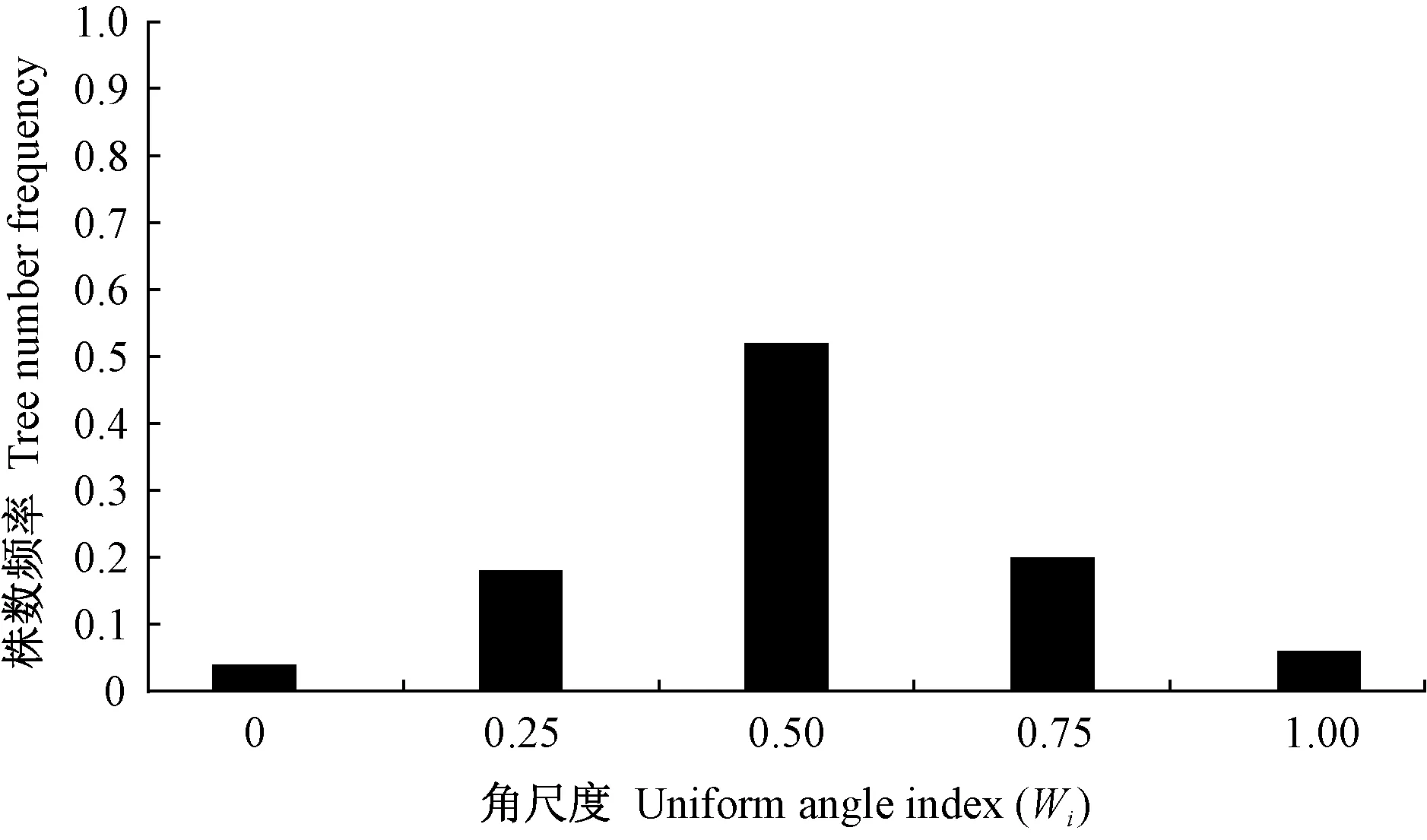
图6 天然林的角尺度分布Fig. 6 Uniform angle index distribution of natural forest
5 结论
基于4株相邻木空间关系的林分空间结构参数——角尺度,将森林群落中的林木分为均匀木(Wi<0.5)、随机木(Wi=0.5)和聚集木(Wi>0.5)。天然林中主要的林木群体为随机分布的林木个体(随机木),随机体作为可以长期存在且不易瓦解的结构体,可为林木提供长效、稳定的微环境,随机体在天然林中的主体构成是促使森林稳定性的关键。
本研究提出森林稳定性维持的“随机体-稳定性”假说,即林分中均匀体由于受四方的遮挡和挤压,使得位于中心的均匀木对环境资源(光)的利用受阻,加大中心木死亡的几率,导致结构体容易解体; 聚集体由于聚集木的邻体相互拥挤从而可增大邻体中个体死亡的几率,也容易导致其结构体解体。林分中随机体的架构形式使得随机木具有两面受光的邻体微环境,相较于均匀体,其中心木受相邻木的竞争压力更小,而相较于聚集体,随机木的邻体之间挤压程度则更小,随机木及其邻体(随机体)相对来说承受的生存压力较小,这种结构体比其他2种结构体中的林木有更大的可能性稳定生长,从而在自然演替过程中能够获得更大的存活几率。随机体被认为是林木在森林群落条件下最为普适的邻体配置形式,植物由于位置的固定性,只有通过适者生存的自然选择法则实现随机体的形成; 随机体数量越多,森林群落越稳定。
