盐生植物盐爪爪液泡膜钠氢反向运输载体基因(KfNHX1)遗传转化拟南芥的耐盐性鉴定
2021-04-01银芳柳毛晓菲曾幼玲
银芳柳,毛晓菲,曾幼玲
(新疆大学生命科学与技术学院/新疆生物资源基因工程重点实验室,乌鲁木齐 830046)
0 引 言
【研究意义】盐胁迫是影响作物产量的主要因素之一。为了应对盐胁迫,植物已进化出多种生理和分子机制,如渗透调节、抗氧化防御以及离子转运和区隔化机制等[1]。其中离子稳态对于维持植物细胞的正常生理代谢非常重要[2]。研究离子稳态的关键基因对于植物耐盐性有重要意义。【前人研究进展】Na+/H+反向转运蛋白(NHX1和SOS1)是一类在离子稳态中起重要作用的膜蛋白,属于单价阳离子/H+反向转运体CPA1家族成员[2]。SOS1位于质膜上,在质外体中排出Na+[3];而NHX1位于液泡膜中,将Na+泵入液泡中来降低胞质中的Na+浓度[4]。通过调节胞间离子的跨膜转运来保持适当的离子浓度,从而避免Na+在胞质中的毒害。植物液泡膜Na+/H+反向运输载体基因(NHX1)已经从多种植物中克隆,如拟南芥(Arabidopsisthaliana)[5]、盐地碱蓬(Suaedasalsa)[6]、甜菜(Betavulgaris)[7]、海马齿(Sesuviumportulacastrum)[8],其过表达都能够增强转基因拟南芥的盐胁迫耐受性。【本研究切入点】藜科盐生灌木盐爪爪在西北盐碱沙漠地区广布,其茎叶肉质化,具有极强的耐盐能力[9]。研究是基于已克隆的盐爪爪KfNHX1基因[10],通过遗传转化拟南芥对其耐盐性进行分析。【拟解决的关键问题】研究农杆菌介导的转盐爪爪KfNHX1基因拟南芥的耐盐性,通过基因组PCR和RT-PCR鉴定转基因株系,在盐胁迫下对转盐爪爪KfNHX1拟南芥的萌发率、根长、离子含量及表型分析,为盐爪爪KfNHX1基因的耐盐分子育种奠定基础。
1 材料与方法
1.1 材 料
野生型拟南芥(Col)和转基因拟南芥种子在1/2 MS(Murashige and Skoog medium)培养基上生长1周,移栽至花土(腐殖土∶蛭石∶珍珠岩= 3∶1∶1)中。培养条件:温度为22℃,相对湿度为40%~60%,光照周期为16 h光照/8 h黑暗。
BamH I、SalI等限制性核酸内切酶、ExTaq、DNA polymerase、dNTP Mixture、DNA Marker、Oligo (dT)18Primer、Reverse Transcriptase M-MLV (RNase H) 和Ribonuclease Inhibitor购自大连宝生物技术有限公司;植物基因组提取试剂盒、植物总RNA提取试剂盒、质粒小提试剂盒购自北京天根生化科技有限公司;引物由上海生工生物工程股份有限公司合成;其余常用试剂为国产分析纯试剂。
1.2 方 法
1.2.1 植物表达载体的构建和农杆菌介导的遗传转化
将KfNHX1(AY825250)的开放阅读框(ORF)克隆到pCAMBIA1301质粒的35S启动子和PolyA信号之间(BamH I、SalI)。将成功构建的质粒转化农杆菌EHA105。采用花序浸染法浸染拟南芥,收获种子。35 mg/L潮霉素筛选转基因种子,筛选T2代符合3∶1的转基因株系,T3代纯合株系用于后续实验。
1.2.2 转KfNHX1基因拟南芥的PCR及RT-PCR鉴定
根据试剂盒说明书分别提取植物样本的基因组和总RNA。通过KfNHX1全长编码序列ORF的引物P1(5'-TCAGGATCCATGTGGTCACAGTTAAGC-3')和P2(5'-GGAGTCGACCTATGTTCTGTCTAGCAAATTGT-3')扩增拟南芥基因组中的KfNHX1;使用KfNHX1部分编码序列引物P3(5'-ATAATCAGTTTACAAGGTCAGGGC-3')和P4(5'-TAATAGTGGACGGTGTGAGTAGGT-3')进行RT-PCR扩增拟南芥cDNA中的KfNHX1。
1.2.3 NaCl和ABA胁迫处理下转基因拟南芥耐性
野生型和T3代转基因拟南芥的纯合种子进行表面灭菌并在4℃ 放置2 d,于植物光照培养箱中生长,培养条件同上。
1.2.3.1 种子萌发率
将野生型和转基因的拟南芥种子播种在含有0、50、75、100、125、150 mM NaCl和1 μM ABA的1/2 MS培养基中,当种子出现子叶时记录为萌发,每天的萌发率为发芽种子数与种子总数之比。每处理共120粒种子用于测定萌发率。
1.2.3.2 苗期根长
将野生型和转基因拟南芥幼苗(5日龄)转移至含有0、120 mM NaCl和10、15 μM ABA的1/2 MS培养基上,3个生物学重复,于植物光照培养室中竖直培养10 d后拍照,采用ImageJ软件测定根长。
1.2.3.3 成苗的胁迫表型及Na+和K+含量
用逐渐增加到200 mM NaCl浇灌生长5周的转基因拟南芥,盐胁迫处理至第15 d拍照。在胁迫第7 d时取拟南芥叶片,置于60℃的烘箱中烘干至恒重。用HNO3处理样品,煮沸至酸液挥发尽,用超纯水定容。使用原子吸收分光光度计(Z-8000,Hitachi,Tokyo)测定Na+和K+含量,每个处理3个生物学重复。
1.3 数据处理
数据表示为平均值±SE(n=3)并且进行单因素方差分析(ANOVA)。进行多重比较以确定组间和组内之间的显著差异。显著性水平为P< 0.05(*);P< 0.01(**);P< 0.001(***)。
2 结果与分析
2.1 转KfNHX1基因拟南芥的遗传转化和鉴定
研究表明,将KfNHX1编码序列构建至具有潮霉素抗性标记的植物双元表达载体pCAMBIA1301中。根癌农杆菌介导花序浸染法转化拟南芥。使用潮霉素抗性(HPT)筛选T2代具有3∶1的转基因株系,进而获得T3代纯合株系。对转基因纯合株系进行分子检测。通过基因组PCR和RT-PCR检测,均在转基因株系中扩增出相应的目的条带,KfNHX1已整合到拟南芥的基因组中,并在RNA水平上转录表达。图1,图2

图 1 pCAMBIA1301-CaMV35S-KfNHX1的构建图谱Fig.1 Physical map of pCAMBIA1301-CaMV35S-KfNHX1 construct
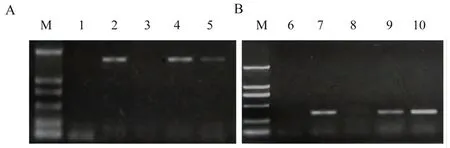
注:A、B:分别为转KfNHX1基因拟南芥中的基因组PCR和RT-PCR分析;M:DL2000 Marker;1、6:阴性对照;2、7:阳性对照;3、8:野生型;4、5、9、10:转基因株系KN6和KN14
2.2 盐和ABA胁迫条件下转KfNHX1基因拟南芥的种子萌发
研究表明,野生型与T3代纯合转基因拟南芥的种子分别在不同浓度(0、50、75、100、125、150 mM)的NaCl以及外源施加ABA(1 μM ABA)处理6 d后测定萌发率。50 mM NaCl和对照条件下野生型和转基因株系种子萌发率没有显著差异,而在75至125 mM NaCl和1 μM ABA处理后,野生型种子萌发率仅约40%,而KN6株系超过80%。相较于野生型,转基因株系在种子萌发率方面表现对盐和ABA的抗性。图3,图4
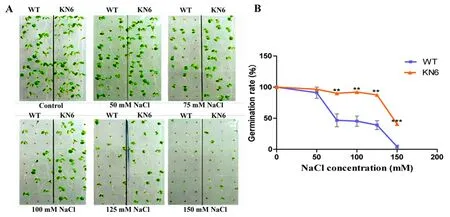
注:A、B:分别为在0、50、75、100、125、150 mM NaCl胁迫下WT和KN6的萌发表型和萌发率
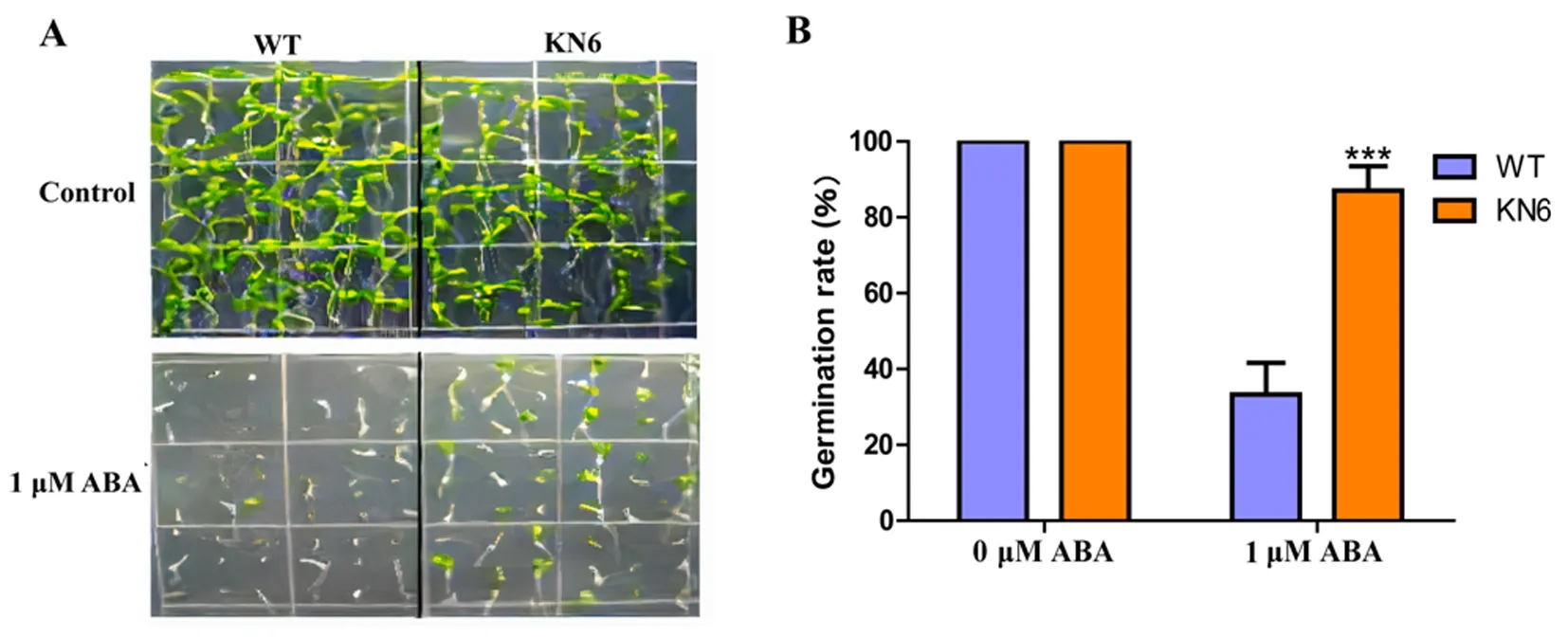
注:A、B:分别为ABA处理下WT和KN6的萌发表型和萌发率
2.3 盐胁迫和ABA处理下转KfNHX1基因拟南芥的抗性
研究表明,转基因植株的生长明显优于野生型,而不处理的对照拟南芥的生长无差异,KfNHX1基因能增强拟南芥苗期的耐盐性。同样,在添加10 μM ABA胁迫激素的培养基上处理10 d,转基因株系生长良好,叶片较大,而野生型的叶片黄化且生长受到严重抑制。转化KfNHX1基因的拟南芥能够降低拟南芥苗期对ABA敏感性。图5
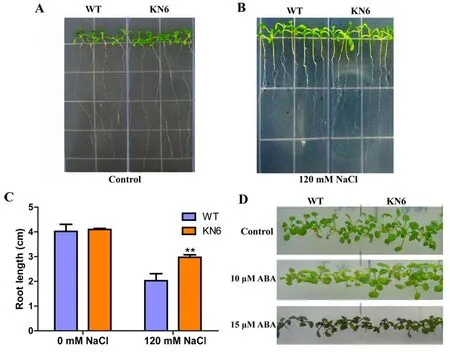
注:A、B:分别为正常生长和120 mM NaCl胁迫下的根长表型;C:根长分析;D:10 μM 和15 μM ABA胁迫下的生长表型
研究表明,成苗的拟南芥,在正常条件下,野生型和转基因拟南芥的生长良好且无差异,200 mM NaCl胁迫15 d时,野生型植株大多生长抑制、枯萎甚至死亡,转基因拟南芥普遍长势较好。利用原子吸收分光光度计测定盐胁迫下野生型和转基因拟南芥叶中的Na+和K+含量的结果显示转基因植株中Na+和K+含量都高于野生型。KfNHX1基因可能增强了将Na+区隔化至液泡的能力,提高了转基因拟南芥的耐盐性。图6

注:200 mM NaCl处理下的拟南芥的生长表型(A)、拟南芥叶中的Na+含量(B)和K+含量(C)
3 讨 论
高盐造成植物水分亏缺、离子毒害,是世界农作物减产的主要原因[11]。其中液泡膜Na+/H+反向运输载体能将胞质中的Na+区隔化至液泡中,维持胞质内较低的Na+水平,来减轻Na+对植物造成伤害[12]。
在拟南芥、番茄、油菜和棉花中过量表达AtNHX1,在200 mM Nacl处理下均能提高转基因的耐盐性。其中转基因棉花产量增加和产生更多棉纤维的现象[13-16]。将互花米草SaNHX2基因过表达至拟南芥,在200 mM NaCl胁迫条件下,与野生型相比,转基因拟南芥的叶绿素含量更高、根更长,和胁迫应答基因的表达量增加明显[17]。过量表达枣树PdNHX6基因至拟南芥,转基因株系的Na+和K+含量均高于野生型,PdNHX6可能增强了拟南芥将Na+区隔化至液泡的能力,从而提高了转基因拟南芥的耐盐性[18]。
盐爪爪是生长于盐碱荒漠区多年生积盐盐生植物,叶肉质、多汁[9]。利用农杆菌介导法将KfNHX1基因转化至拟南芥进行耐盐功能分析。盐胁迫下,转基因拟南芥种子的萌发率、根长和成苗的生长表型明显优于野生型,Na+和K+含量也高于野生型,表明盐爪爪KfNHX1能够增强拟南芥的耐盐性。这与文献中报道在拟南芥中GhNHX1[19]、ZmNHX1[20]、SsNHX1[6]和MsNHX1[21]的过表达通过提高积累Na+来增强其耐受性的结果一致。
ABA是一种重要的植物胁迫激素,参与种子休眠、萌发和幼苗生长等发育过程及植物的非生物胁迫的信号转导过程[22-24]。过表达水稻OsPP108的拟南芥植株在种子萌发、根生长和幼苗生长过程中对ABA高度不敏感,对高盐和甘露醇胁迫耐受性强[25]。在外源ABA处理下,转大麦HbMBF1a拟南芥都具有较高的子叶绿化率、幼苗根长和逆境响应基因的表达量,增强了转基因拟南芥的ABA不敏感性,是ABA反应的正调控因子[26]。也开展了转KfNHX1基因拟南芥对ABA的胁迫耐受性,通过萌发率和幼苗的生长表型来看,转基因拟南芥降低了对ABA的敏感性,与一些文献的研究结果一致[27-28]。
4 结 论
以农杆菌介导花序浸染法将KfNHX1基因转入拟南芥中并获得了T3代纯合株系。盐胁迫下转基因株系的拟南芥种子的萌发率和根长明显高于野生型。结合200 mM NaCl胁迫处理15 d下的拟南芥成苗表型,相较野生型叶片萎黄和死亡,转基因植株的生长状况较好,且积累了较高的Na+和K+。异源表达KfNHX1能够提高转基因拟南芥的耐盐性。
