不同放牧方式对典型草原草场植被变化的影响
——以中蒙边境两侧对应嘎查为例
2021-03-31其力木格高苏日固嘎赛西雅拉图乌兰图雅
其力木格, 高苏日固嘎, 赛西雅拉图, 乌兰图雅, 罗 培
(内蒙古师范大学 地理科学学院,内蒙古 呼和浩特 010022)
蒙古高原处于欧亚大陆中部,气候以典型的大陆性干旱半干旱气候为主,它既是个气候变化的敏感区又是生态系统的脆弱区[1]。蒙古高原典型草原主要分布在中国内蒙古东北部地区和蒙古国东部地区。游牧制度在蒙古高原传承几千年,至今也是蒙古国主要的生产方式,而中国内蒙古自治区近30年来,以定居放牧为主,草地被围栏分割,游牧逐渐消失。近半个世纪以来,蒙古高原草原均出现了不同程度的退化现象[2]。由于不合理的利用方式和气候变化导致草地生产力下降,草畜矛盾日益尖锐,植被多样性和稳定性下降[1-3]严重威胁着草地畜牧业的可持续发展。因此,草地科学管理和持续利用是一个备受人们重视的研究方向[4]。
放牧是草地利用的主要方式之一,其实质是一种生态干扰[5]。放牧作为草地的利用方式,对植物生长既有抑制作用,又有促进作用[6]。不同放牧措施对草地植物群落的影响存在差异[7]。禁牧是恢复退化草地生态系统的有效途径,但长期禁牧不利于维持较高的群落多样性[7-9]。放牧干扰过于频繁剧烈,植物多样性降低; 相反放牧干扰影响较少时群落由优势种统治,提高物种和群落多样性,促进植被生长[7-9]。许中旗等[10]的研究表明,典型草原的物种多样性变化与“中度干扰假说”一致,即物种多样性指数和丰富度指数都是在禁牧时间中等的情况下达到最大,而在放牧和禁牧时间较长的情况下相对较低。李学斌等[11]的研究表明,在中国北方干旱半干旱草地,围封一段时间后,适当的割草或放牧有利于加速退化草地恢复。周建琴等[12]的研究表明,连续放牧区由于长期的放牧压力,羊群啃食和践踏,草地趋于干旱化,各物种竞争关系发生变化,抑制优势物种对草地资源的竞争,旱生植物逐渐增多。
迄今为止,很少有对蒙古高原的跨境地区进行比较研究,有关遥感数据结合地面数据多尺度的研究更少。因此本文研究不同放牧方式下的草场植被群落特征变化,分析哪种放牧更有利于草地资源可持续利用和畜牧业健康发展,为牧民合理放牧、维护草场生态和促进人与草原和谐共处提供理论支撑。
1 研究区概况
额尔敦敖包嘎查、阿拉坦陶高图嘎查、阿日宝拉格嘎查是我国阿巴嘎旗原那仁宝拉格苏木(2006-2013年)的三个嘎查(图1中的研究区1)。那仁宝拉格苏木地处113°28′~114°06′E,44°10′~44°47′N。东邻青格勒宝拉格苏木,西与苏尼特左旗接壤,南接宝格达乌拉苏木,北与蒙古国接壤。地形属于高原丘陵区,北高南低,温带大陆性半干旱气候,年降雨量为223 mm左右,无霜期为100~110 d。主要植物有羊草(Leymuschinensis)、克氏针茅(Stipakrylovii)、寸苔草(Carexduriuscula)、灰绿黎(Chenopodiumglaucum)、猪毛菜(Salsolacollina)等。其放牧方式主要从游牧性放牧转化为连续性放牧。
陶森郭勒巴格是蒙古国纳兰苏木的一个嘎查(图1中的研究区2)。纳兰苏木位于蒙古国苏赫巴托省南部,距我国新浩特212 km; 南部与我国阿巴嘎旗接壤,东与达仁刚嘎苏木、西与翁根苏木、北与阿斯哈图苏木接壤。地理坐标为43°04~44°26′N,113°27′~114°11′E。气候同上,年降水量178.5 mm左右; 地形南高北低,波状高原缓丘陵,局部花岗岩出露; 土壤以淡栗钙土和栗钙土为主。主要植物有羊草(Leymuschinensis)、克氏针茅(Stipakrylovii)、冰草(Agropyroncristatum)等。其放牧方式主要以游牧为主。

图1 研究区域及现场取样实验设计Fig.1 Study area and design of the field sampling experiment注: 该图基于自然资源部标准地图服务网站下载的审图号为GS(2016)1600号的标准地图制作,底图无修改。
2 数据与研究方法
2.1 数据
2.1.1 遥感数据 遥感影像是http://glovis.usgs.gov/网站下载的LandsatTM、ETM数据图像,分辨率为30 m。分别选取1987年6月27日、1999年7月19日、2003年8月10日、2010年6月26日、2016年7月28日的影像,通过一系列图像处理,即几何校正、大气校正、辐射校正、最后用像元二分法提取植被覆盖度。
2.1.2 调研数据 研究组于2016年7月下旬在研究区进行野外调研。主要采样方法为: 在两国国境线两侧的对应嘎查区域中蒙跨取3条调研样带(图1)。在两国境内共取相隔3 km间隔的21个样组,每一样组由三个相隔300 m的样方组成。所有植物样方所测数据经归总无误后汇总并保存。每个样方中分别测量以下指标: 地上生物量、平均高度、平均密度、平均盖度。放牧方式与畜群结构则是通过对两国境内的对应苏木牧民进行访谈获得。
2.2 研究方法
2.2.1 植被群落特征测定方法 植被测定及其计算方式如下:
重要值=(相对高度+相对密度+相对盖度)/3,
(1)
相对高度=(某一植物种群的平均高度/各植物种总平均高度之和)×100%,
(2)
相对密度=(某一植物种群的个体数/全部植物种群的个体数之和)×100%,
(3)
相对盖度=(某一植物种群的平均盖度/各植物种群的平均分盖度之和)×100%。
(4)
2.2.2 像元二分法 像元二分法[14]是一种简便实用的线性模型,它假设一个像元的地表信息由纯植被和纯土壤(非植被)两部分信息构成,所得到的光谱信息是由两种纯组分的面积比例加权的线性组合。具体可以表示为:
S=If vc×Svge+(1-If vc)×Ssoil,
(5)
对(6)式进行变换,得到植被覆盖度公式:
If vc=(S-Ssoil)/(Svge-Ssoil),
(6)
其中:Ifvc表示像元中纯植被所占的面积比例,即植被覆盖度;S为像元的光谱信息;Sveg和Ssoil分别表示像元中的纯植被像元信息和纯土壤像元信息。
2.2.3 群落物种总体稳定性M.Gordon指数 采用M.Gordon贡献定律法,将研究区所采植物样方内所有植物种类按频度大小依次排序,并转化为相对频度逐步累加。然后按植物种类排序的顺序将植物种类的总和取倒数,并依次累加。两者一一对应并以植物种类倒数百分比累计值为x轴,累计相对频度为y轴绘图,建立模糊散点平滑曲线二次函数y=ax2+bx+c,再与一次函数y=x+100相交,得交点坐标(x,y) 。最后计算交点与稳定性坐标点 (20,80) 的距离,距离越近则表示群落物种总体稳定性越高[15]。
3 结果分析
3.1 不同放牧方式下地上生物量比较
在三种放牧方式下,地上生物量差异显著(P<0.05,图2); 禁牧区地上平均生物量 [(455.9±219)g]显著(P<0.05)大于游牧区 [(268.4±130)g]和连续放牧区 [(122.2±54)g],连续放牧区的平均地上生物量最低。地上生物量与实测覆盖度之间有较好的相关(R2=0.71,P<0.05,图3)。
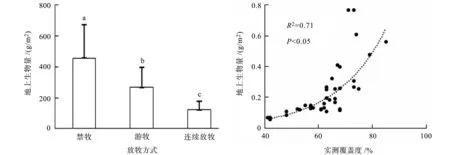
图2 不同放牧方式下的平均地上生物量 图3 地上总生物量与实测覆盖度之间相关性 Fig.2 Average aboveground biomass under Fig.3 Correlation between aboveground different grazing systems biomass and measured cover 注: 不同字母表示显著差异(P<0.05),下同。
3.2 不同放牧方式下的主要植物物种比较
蒙古高原典型草原,原属的植被样及其组成样都为大针茅与克氏针茅群系[8]。研究区是由原来的克氏针茅群系和大针茅草原退化而来的克氏针茅草原共同组成,其主要植物种为克氏针茅、羊草、冷蒿和局部大针茅等[16]。
本研究中,游牧区、连续放牧区、禁牧区植物总数分别为48、46和33种。由表1可得知游牧区草场群落内原生建群种重要值占比比连续放牧区和禁牧区高。连续放牧区大针茅优势度为零,其建群种由大针茅变为克氏针茅,草地沙化退化指示植物猪毛菜(Salsolacollina)进入植物群落的主要优势种行列,并且其在主要植物种中占比为10.46%。由于长期禁牧,禁牧区群落中优势种的比重较低,植物种类多样性下降,且一年生植物在主要植物种中占比17.5%要比连续放区(10.45%)和游牧区(0%)高。可见,不同放牧方式下,植物群落组成存在一定差异。
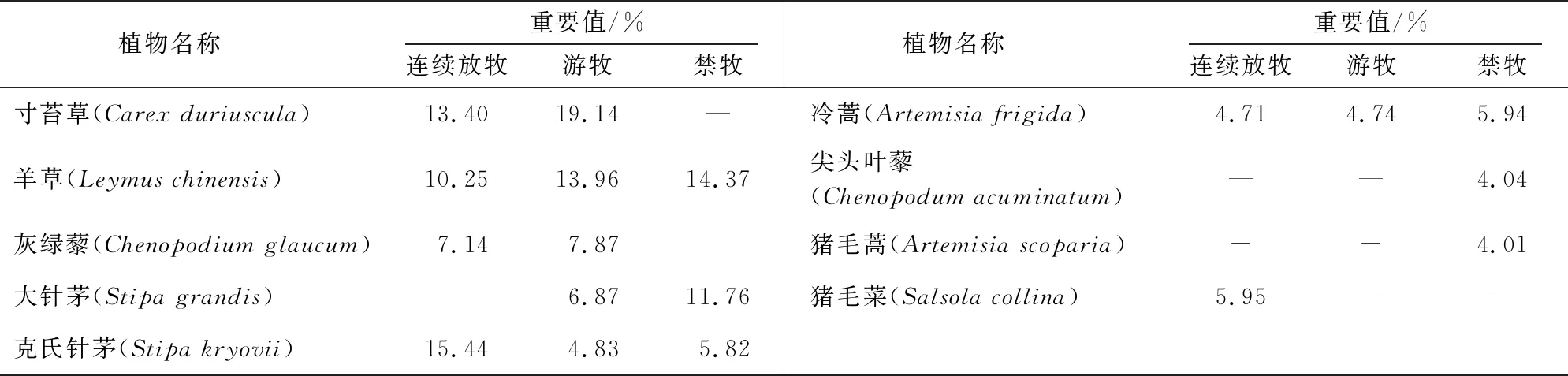
表1 研究区主要植物的重要值Tab.1 Important value of main plants in the study area
3.3 植被覆盖度计算结果验证
为验证NDVI像元二分模型应用于植被覆盖度反演是否有效,本研究在61个采样点中选取40个1 m×1 m 样点进行实地采样验证,用GPS定位,获得采样点的经纬度坐标。最后根据经纬度坐标将从影像上获取的估算结果与实地测量结果进行相关性分析,选取的影像为2016年7月28日数据。结果表明,应用NDVI像元二分模型估算的结果与实测值间有较高的相关性,相关系数R2达到0.86(图4),表明用像元二分模型估算的植被覆盖度是可靠的,该结果与佟斯琴等[17]的结果较一致。
3.4 不同放牧方式下植被覆盖度变化分析
为确保植被覆盖度具有代表性,对于61个采样点像素,每一个采样点附近选取5个像素点,然后用6个像素点的平均值代替采样点植被覆盖度。结果表明,在1999年、2003年、2010年、2016年游牧区植被覆盖度均大于连续放牧区,且2003年、2010年和2016年差异显著(P<0.05,图5)。1987年连续放牧区植被覆盖度显著(P<0.05)大于游牧区。禁牧区除2010年以外植被覆盖度均高于游牧区。

图4 植被覆盖度估算结果与实测结果相关性分析 图5 不同放牧方式下植被覆盖度随时间的变化 Fig.4 Correlation analysis between estimated Fig.5 Changes of vegetation cover with time Ifvc and measured Ifvc under different grazing ways
3.5 不同放牧方式下植物群落整体稳定性分析
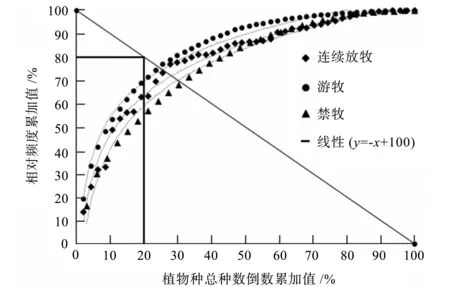
图6 不同放牧方式M.Gordon稳定性模拟曲线Fig.6 Stability simulation curves of M.Gordon with different grazing ways
从图6可知,M.Gordon植物群落稳定性测验方法,对样方进行拟合。得到游牧区草场的稳定性二次函数为y=22.018 ln (x)+3.690 4,连续放牧区草场的稳定性二次函数为y=26.753 ln (x)-20.864,禁牧区草场的稳定性二次函数为y=26.753 ln (x)-20.864。分别与一次函数y=x+100交于点(22,72)、(24,68)和(26,67)三点,三个交点距离稳定点坐标(20,80)的距离分别为8.2、12.65和14.32。可见,游牧区距离稳定点坐标更近,其次是连续放牧区,禁牧区距离稳定点坐标最远。总体而言,游牧区植被群落更稳定。因此,适度放牧对植被群落稳定性有一定好处。
4 讨论与结论
放牧干扰是影响草原植被群落特征、造成草地退化的主要因子[7-10]。从放牧方式转变来看,蒙古国陶森巴格的放牧方式到目前为止变化较缓慢,基本保持以浩特为单元进行游牧,随着社会的发展和牲畜的所有权变化(1990年前公社制,其后私有制),游牧距离和次数变少,但放牧方式基本维持游牧状态,只转为限定苏木区域的游牧。而我国那仁宝拉格苏木1983年后是从浩特为单元的集体游牧方式逐渐变成今日的以牧户浩特为单元的草畜双全个体经营的定居放牧方式,即分级别限区(嘎查、浩特、分浩特、牧户浩特和子牧户浩特),转向为小区高频度两季轮牧方式[2-3]。由于牧户所分得的草场面积是固定的,但是人口越来越多[16],草场不断细分,人均草场面积逐步缩小,放牧压力增大[18]。在此背景下,草场植被群落特征也相应产生变化。
家畜主要通过采食、践踏及排泄三种途径影响草地植被群落结构组成、枯落物及地上生物量。放牧后草地生物量、凋落物量及土壤特征均会发生变化[18]。本研究中,禁牧区地上平均生物量显著大于游牧区和连续放牧区(如图2所示),这与已有研究相似[19]。在放牧过程中,家畜会采食植株的一部分,造成植被地上现存量减少; 相应的,植株在正常枯死之前就已被采食,枯落物相对减少; 并且家畜在游走过程中的践踏作用也会造成枯落物量的相对降低,地上生物量随着放牧强度的增加而逐渐降低[20]。不同放牧方式下,群落物种组成不同。本研究发现游牧区主要植物种的优势度比连续放牧和禁牧区要高。连续放牧区草地沙化退化指示植物猪毛菜成为植物群落的优势种。禁牧区由于长年禁牧,植物多样性下降。研究表明,合理放牧下植物群落可进行一定程度物种组成的调整,连续放牧下,家畜的连续采食抑制了多年生植物生长,一年生植物重要值比例上升,不利于草地植物群落稳定结构。适度放牧是保护植物功能群,物种多样性,维护生态系统基本功能和健康状态的有效途径[2,5]。
M.Gordon稳定性操作方法简单,主要用于群落结构的评估,即假定物种数目和频度按照某种排列组合时,群落结构达到稳定状态,群落内物种不易消失,外来物种不易入侵[21]。物种消失易导致群落某种植物功能的丧失,而入侵物种会改变群落的原有组成,二者均会对群落稳定性产生影响。此外,围封年限也会对群落M.Gordon稳定性产生影响。短期围封有助于围栏内群落M.Gordon稳定性的增加。随着围封时间的增加,围栏外的群落显得更加稳定[22]。通过M.Gordon植物群落稳定性测定法,发现游牧区距离稳定点坐标更近,其次是连续放牧区,禁牧区距离稳定点坐标最远,说明游牧区植物群落稳定性为最好,连续放牧区次之,禁牧区最差,这与廖晗茹等[23]的研究结论相一致。

图7 1989-2016年研究区年降水量的年际变化Fig.7 Inter-annual variability of annual precipitation in the study areaduring 1989-2016
除1987年,其他所有时间点上,游牧区的植被覆盖均大于连续放牧区(如图5所示),表明连续放牧区草地退化程度大于游牧区。很多研究结果表明,轮牧可以提高草料产量和草地利用率[18,20],较连续放牧、游牧能更好地维持草场健康。但也有研究表明,降水强烈影响了蒙古高原植物群落的演化[7]。通过对研究区内近30年来的降水数据分析后发现,在1987年至2016年期间,研究区整体降水的减少(图7),诠释了整个研究区植物群落的退化。但在2010年游牧区的植被覆盖度高于禁牧区植被覆盖度则表明,适当的放牧导致最大程度的植物和功能多样性,因此可能导致植物物种的丰富度和生物量整体增加。
本研究结果表明,不同放牧方式下草场植被群落特征发生相应变化。
草场群落内植物组成结构出现显著差异。游牧区草场群落内原生建群种重要值较高,连续放牧区多年生植物比重降低,植物的一年生现象比较严重。禁牧区植物种类多样性下降。
禁牧区地上生物量大于游牧区和连续放牧区,地上生物量与植被覆盖度高度相关。游牧区植被覆盖度均大于连续放牧区的2003年、2010年和2016年; 1987年连续放牧区植被覆盖显著大于游牧区。
游牧区草场植物群落的稳定性高于连续放牧区和禁牧区,禁牧区草场植物群落的稳定性最低。
轮牧比起连续放牧能更好地维持草场健康,游牧放牧是相对最优的放牧方式,更有利于草地资源可持续利用和畜牧业健康发展。
