日本细毛环腹瘿蜂生物学特性研究
2021-03-30李银煳杨凡萱张晓明陈国华
李银煳,张 峰,杨凡萱,肖 春,张晓明*,陈国华*
( 1. 云南农业大学植物保护学院/云南生物资源保护与利用国家重点实验室,昆明650201;2. 中国农业科学院植物保护研究所,农业农村部-CABI 生物安全联合实验室,植物病虫害生物学国家重点实验室,北京100193)
斑翅果蝇DrosophilasuzukiiMatsumura,1931,又称铃木氏果蝇,隶属于双翅目Diptera果蝇科Drosophilidae果蝇属Drosophila,在我国多个省份均有发生(钱远槐等,2006)。1916年于日本山梨县草莓园中第一次被发现,1931年由Matsumura描述并命名,随后扩散到美洲、欧洲等国家(Hauseretal.,2011)。斑翅果蝇不仅取食表皮受损的果实或地上的落果,还能够危害成熟期间的新鲜果实(Keeseyetal.,2015)。雌虫用锯齿状产卵器划破较为成熟或已经成熟的新鲜软皮水果表皮,并将卵产于果肉中,其幼虫在果实内危害,果实以取食点为中心向四周迅速腐烂软化,失去食用价值,给水果产业造成巨大的经济损失(Atallahetal.,2014)。由于斑翅果蝇适应能力强、寄主植物多样、发育速度快、繁殖率高,以及在入侵地缺乏自然天敌,已引起了国内外广泛的关注(Chabertetal.,2012;张开春等,2014;Kenisetal.,2016)。近年调查发现,国内斑翅果蝇对樱桃、蓝莓、杨梅以及葡萄危害的报道日趋增多,云南省杨梅成熟期受斑翅果蝇危害较为严重,危害率在30%~40%,在杨梅全部成熟之时,斑翅果蝇的危害率可达100%(蔡普默等,2017;刘冰,2017)。斑翅果蝇入侵北美和欧洲以来已经成为世界性的危害浆果类和核果类水果的重要害虫之一(Walshetal.,2011;Cinietal.,2012;Asplenetal.,2015)。
由于斑翅果蝇的危害特点,使用传统化学农药很难取得较好的防治效果,大量使用化学农药容易产生“3R”等一系列问题,同时也危及到许多有益昆虫如天敌和授粉昆虫的生存(倪伯杰,2015; 蔡普默等,2017)。以天敌昆虫为对策的斑翅果蝇生物防治措施在成本效益和环境安全方面均具有优势,是重要的防治措施之一(Hayeetal., 2016)。
云南省斑翅果蝇寄生性天敌昆虫较为丰富,调查到斑翅果蝇寄生性天敌昆虫有5 种,即:丽盾瘿蜂Ganaspisbrasiliensis、日本细毛环腹瘿蜂Leptopilinajaponica、反颚茧蜂Asobarasp.、果蝇锤角细蜂Trichopriadrosophilae和蝇蛹金小蜂Pachycrepoideusvindemmiae。其中丽盾瘿蜂、细毛瘿蜂和反颚茧蜂为斑翅果蝇的幼虫寄生蜂,果蝇锤角细蜂和蝇蛹金小蜂为斑翅果蝇的蛹寄生蜂(Daaneetal.,2016;方圆等,2019)。日本细毛环腹瘿蜂Leptopilinajaponica,隶属于膜翅目Hymenoptera环腹瘿蜂科Figitidae细毛瘿蜂属Leptopilina,可以在斑翅果蝇体内完成发育(Kasuyaetal.,2013;Daaneetal.,2016)。在云南调查到的日本细毛环腹瘿蜂占寄生蜂总数的34.92%,是斑翅果蝇的主要寄生性天敌昆虫之一(方圆等,2019)。目前国内对斑翅果蝇的蛹寄生蜂果蝇锤角细蜂和蝇蛹金小蜂的研究有零星报道(羊绍武等,2019;仪传冬,2019;周长清等,2019),而对斑翅果蝇的幼虫寄生蜂鲜有报道,本文对斑翅果蝇幼虫寄生蜂日本细毛环腹瘿蜂生物学特性进行系统研究,以期为日本细毛环腹瘿蜂的繁育、保护利用和斑翅果蝇的绿色防控提供科学依据。
1 材料与方法
1.1 实验材料
斑翅果蝇:采自云南省昆明市甸尾乡杨梅园。在实验室条件23±2℃,RH60%±5%,于养虫笼中饲养,提供剥皮香蕉饲喂,香蕉每48 h更换一次。
日本细毛环腹瘿蜂:采自云南省昆明市甸尾乡杨梅园。在实验室环境条件下,饲养于养虫笼中,提供20%蜂蜜水喂养和提供斑翅果蝇产卵48 h的香蕉柱供其产卵寄生。
香蕉:购买于沃尔玛超市都乐牌香蕉。
果蝇养虫笼:长50 cm×宽50 cm×高50 cm的纱网养虫笼。
寄生蜂养虫笼:长38 cm×宽38 cm×高38 cm的纱网养虫笼。
养虫罐:长12 cm×宽9 cm×高12 cm的玻璃罐。
1.2 不同营养条件下日本细毛环腹瘿蜂成虫寿命测定
收集羽化0.5 h内的日本细毛环腹瘿蜂,单独置于指形管(直径1 cm×高7 cm)中,每管1头,将浸有质量浓度分别为10%、20%蜂蜜水、清水的脱脂棉团和不加任何物质的脱脂棉团分别塞于指形管口,实验室条件下饲养,以不加任何物质的脱脂棉团为空白对照。每个处理观察雌蜂、雄蜂各20头,共40头虫。每24 h更换补充营养源,每24 h观察1次,观察直到所有供试寄生蜂死亡为止,记录寄生蜂的死亡时间和数量,统计寄生蜂寿命(杨琪,2017)。
1.3 日本细毛环腹瘿蜂行为节律观察
1.3.1日本细毛环腹瘿蜂的羽化节律
选取同批次供日本细毛环腹瘿蜂寄生的斑翅果蝇蛹放到垫有润湿棉花的培养皿(D=9 cm)上保湿培养,每个培养皿放供试的斑翅果蝇蛹100头,将培养皿放入塑料饲养盒(直径12 cm×高4.5 cm)中,3次重复,共300头蛹。在实验室条件下观察寄生蜂羽化,6 ∶00-20 ∶00每2 h观察1次,记录各时段羽化的寄生蜂数量。观察直到3 d内没有寄生蜂羽化为止(杨琪,2017)。
1.3.2日本细毛环腹瘿蜂的产卵活动节律
取日本细毛环腹瘿蜂雌蜂10头与雄蜂10头配对分别放入养虫罐中,并提供有斑翅果蝇幼虫的香蕉段供其寄生产卵,每48 h更换香蕉段,并提供20%蜂蜜水作为补充营养。于6 ∶00至次日2 ∶00 每2 h观察1次,每次持续观察1 h,连续观察4 d。寄生蜂在香蕉段上活动并有刺探行为视为具有产卵活动行为,记录具有产卵活动行为的寄生蜂数量和时间。
1.4 日本细毛环腹瘿蜂寄主选择与寄生动态测定
1.4.1对斑翅果蝇不同龄期幼虫非选择性寄生比较
将装有剥皮香蕉切段的培养皿放入斑翅果蝇养虫笼内供其产卵6 h后取出,待斑翅果蝇发育到1龄、2龄、3龄幼虫时,分别放入养有若干日本细毛环腹瘿蜂的养虫笼内产卵20 h后取出分别放入饲养盒中,在实验室条件下进行饲养观察至斑翅果蝇和寄生蜂完全羽化,分别记录其羽化的虫口数量,每处理设4次重复。
1.4.2对斑翅果蝇不同龄期幼虫选择性寄生比较
不同龄期的斑翅果蝇幼虫获取同上。分别将携带斑翅果蝇1龄幼虫和2龄幼虫的5段香蕉放入养虫笼中内侧两个边角,在笼口正下方放有20%蜂蜜水脱脂棉团作为补充营养。在日本细毛环腹瘿蜂雌蜂产卵高峰时段接入20头已交配无产卵经历的日本细毛环腹瘿蜂雌蜂于有20%蜂蜜水脱脂棉团上,持续观察2 h,记录其第一次对携带斑翅果蝇各龄期幼虫香蕉的选择与刺探的寄生蜂数量,后移除香蕉在实验室条件下饲养观察至斑翅果蝇和寄生蜂完全羽化,记录其羽化的虫口数量。设4次重复,每次重复调换携带不同龄期幼虫香蕉位置。
1.4.3日本细毛环腹瘿蜂产卵动态
取羽化2 h内的日本细毛环腹瘿蜂雌、雄寄生蜂1对放入养虫罐中,提供有斑翅果蝇幼虫的香蕉段供其产卵寄生48 h,每48 h移除香蕉段,并更换新的有斑翅果蝇幼虫的香蕉段,直至该对寄生蜂蜂死亡为止,提供20%蜂蜜水补充营养,观察并记录斑翅果蝇和寄生蜂羽化的虫口数量,对无果蝇也没有寄生蜂羽化的蛹进行解剖镜检,确认其是否被产卵,设4次重复。
1.5 日本细毛环腹瘿蜂不同发育阶段发育历期测定
将剥皮香蕉切段,放入斑翅果蝇养虫笼内产卵6 h后取出,待斑翅果蝇发育至2龄幼虫时,放入养有若干寄生蜂养虫笼供其寄生6 h后取出,在实验室条件下饲养观察,每隔6 h从香蕉段内取10头斑翅果蝇幼虫进行解剖,记录寄生蜂发育情况,当寄生蜂卵全部孵化后,每24 h观察一次,直至寄生蜂羽化,设4次重复。
1.6 数据处理及分析
采用Microsoft Office Excel计算均值。寄生蜂寿命用单因素方差分析(ANOVA)并应用Tukey 方法进行多重比较分析。寄生蜂幼虫存活率、蛹存活率、性比、发育历期、选择偏好采用独立样本t检验进行差异显著性分析。试验数据在SPSS19.0软件中进行统计分析。性比为雌性所占百分比。
2 结果与分析
2.1 不同营养条件下日本细毛环腹瘿蜂寿命
在不同营养条件下日本细毛环腹瘿蜂的成蜂寿命有差异(表1),与对照相比,清水、10%蜂蜜水、20%蜂蜜水处理都能不同程度的延长雌蜂和雄峰的寿命,雌蜂的寿命给予20%蜂蜜水处理达到最长,为34.55 d,显著高于10%蜂蜜水、清水和对照的寿命(F3,79=55.24,P<0.001),给予10%蜂蜜水处理的寿命次之,为25.75 d,显著高于清水和对照的寿命,雌蜂的寿命给予清水和对照处理无显著差异;雄蜂的寿命给予20%和10%蜂蜜水处理,分别为30.25 d和21.60 d,显著高于清水和对照处理(F3,79=49.39,P<0.001),雄蜂的寿命给予20%蜂蜜水、10%蜂蜜水处理无显著差异,给予清水和对照处理无显著差异。总体统计结果显示,不同补充营养条件下,日本细毛环腹瘿蜂成虫的寿命长短依次为:20%蜂蜜水>10%蜂蜜水>清水>对照,给予清水和对照处理的寄生蜂寿命无显著差异,其它各处理间差异显著(表1)。
2.2 日本细毛环腹瘿蜂行为节律
2.2.1日本细毛环腹瘿蜂的羽化节律
从第一头日本细毛环腹瘿蜂羽化开始至寄生蜂全部羽化持续时间共8 d。第3-5天羽化的寄生蜂数量最多,其中第4天为寄生蜂羽化高峰期(图1A)。在羽化期中每天6 ∶00-20 ∶00均有寄生蜂羽化,早晨6 ∶00-8 ∶00羽化的寄生蜂数量为(8 头/2 h),之后寄生蜂羽化的虫口数量迅速上升,至10 ∶00-12 ∶00羽化的寄生蜂数量最多(26 头/2 h),达到寄生蜂羽化的峰值,之后逐渐下降,12 ∶00-14 ∶00羽化的寄生蜂数量为(12.33 头/2 h),18 ∶00-20 ∶00羽化的寄生蜂数量仅为(1.33头/2h),20 ∶00至次日6 ∶00无寄生蜂羽化(图1B)。从羽化的总体数量统计结果显示,上午6 ∶00至12 ∶00羽化的寄生蜂数量(56.33头)高于下午12 ∶00-20 ∶00羽化的寄生蜂数量(34.67头)。
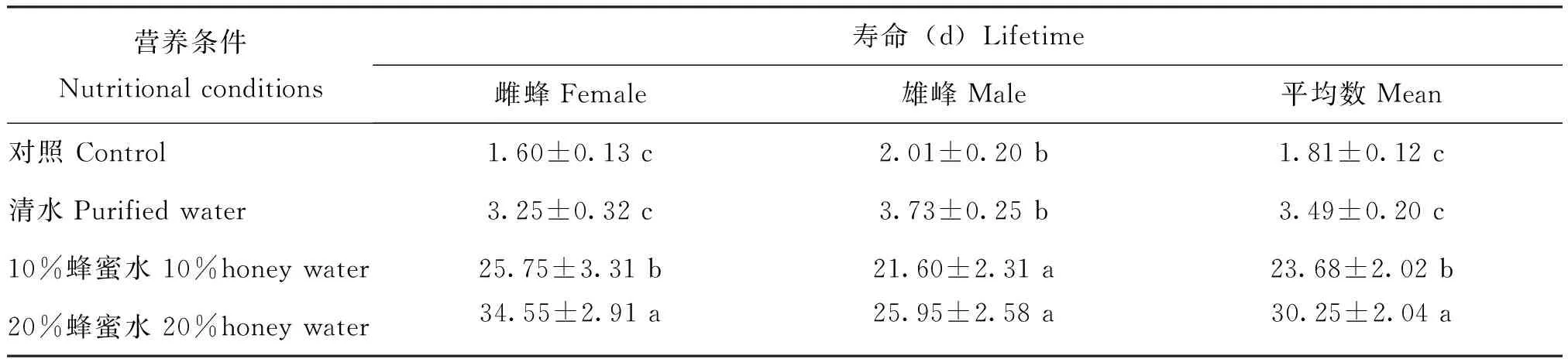
表1 不同营养条件下日本细毛环腹瘿蜂寿命

图1 日本细毛环腹瘿蜂的不同时间羽化节律(A)和不同时间段羽化节律(B)Fig.1 Emergence rhythm of Leptopilina japonica at different day (A)and different duration (B)注:图中数值为平均值±标准误,图2-4同。Note: The data were mean±SE, the same for figure 2-4.
2.2.2日本细毛环腹瘿蜂的产卵活动节律
在日本细毛环腹瘿蜂产卵期,每天6 ∶00至次日2 ∶00均有产卵活动行为。6 ∶00-14 ∶00具产卵活动行为的寄生蜂数量缓慢增加,14 ∶00-22 ∶00具有产卵行为寄生蜂数较多,16 ∶00-18 ∶00具产卵活动行为的日本细毛环腹瘿蜂数量达到高峰,为4.50头,22 ∶00至次日2 ∶00具产卵活动行为的寄生蜂数逐渐减少(图2)。

图2 日本细毛环腹瘿蜂不同时间段的产卵活动节律Fig.2 Oviposition activity rhythm of Leptopilina japonica at different duration
2.3 日本细毛环腹瘿蜂寄主选择与寄生动态
2.3.1日本细毛环腹瘿蜂对斑翅果蝇不同龄期幼虫非选择性寄生
日本细毛环腹瘿蜂对斑翅果蝇不同龄期幼虫的非选择性寄生比较发现,日本细毛环腹瘿蜂只寄生斑翅果蝇的1龄幼虫和2龄幼虫,斑翅果蝇的3龄幼虫均没有被寄生。
日本细毛环腹瘿蜂寄生不同龄期斑翅果蝇幼虫对其生长发育有一定的影响,表现为寄生不同龄期斑翅果蝇幼虫对寄生蜂后代幼虫的存活率(t=0.39,df=6,P=0.71)、雌蜂(t=0.56,df=6,P=0.60)和雄蜂(t=1.87,df=6,P=0.11)的发育历期没有显著的影响,但对日本细毛环腹瘿蜂后代蛹的存活率(t=3.57,df=6,P<0.05)、性比(t=11.55,df=6,P<0.01)、总体的发育历期(t=3.08,df=6,P<0.05)影响显著(表2)。寄生于斑翅果蝇2龄幼虫的日本细毛环腹瘿蜂后代蛹的存活率、性比显著高于寄生斑翅果蝇1龄幼虫,说明斑翅果蝇2龄幼虫对日本细毛环腹瘿蜂有更高的适合度,有利于寄生蜂的繁衍。

表2 日本细毛环腹瘿蜂对斑翅果蝇不同龄期幼虫非选择性寄生比较
2.3.2日本细毛环腹瘿蜂对斑翅果蝇不同龄期幼虫选择性寄生
日本细毛环腹瘿蜂对斑翅果蝇不同龄期幼虫选择性寄生的结果显示(图3),日本细毛环腹瘿蜂选择携带斑翅果蝇2龄幼虫香蕉段的寄生蜂数为12.25头,显著高于选择携带斑翅果蝇1龄幼虫香蕉段的寄生蜂数4.75头(t=8.43,df=6,P<0.05)。再将经日本细毛环腹瘿蜂产卵寄生的携带不同龄期斑翅果蝇幼虫香蕉段放入养虫罐进行进一步饲养观察,结果表明,从携带斑翅果蝇2龄幼虫香蕉段中羽化出的日本细毛环腹瘿蜂数量为11.50头,显著高于从携带斑翅果蝇1龄幼虫香蕉段中羽化出的寄生蜂数量3.25头(t=7.20,df=6,P<0.05)。说明日本细毛环腹瘿蜂对斑翅果蝇2龄幼虫具有选择偏好。
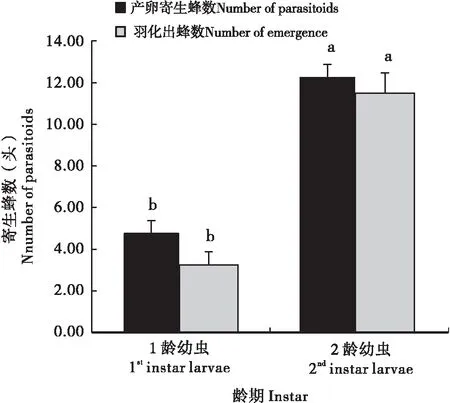
图3 日本细毛环腹瘿蜂对斑翅果蝇不同龄期幼虫选择性寄生比较Fig.3 Choice test comparison between different larvae instar of Drosophila suzukii by Leptopilina japonica注:图柱上不同字母表示不同龄期幼虫在产卵寄生蜂数或羽化出蜂数经t法检验在 P<0.05 水平上差异显著。Note: Different letters above bars indicated that significant difference the parasitoids or the number of emergence between different larvae instar at P<0.05 level by t-test.
2.3.3日本细毛环腹瘿蜂产卵寄生动态
日本细毛环腹瘿蜂羽化后的第1-23天都有产卵寄生行为,3日龄的日本细毛环腹瘿蜂产卵寄生量达到高峰,为13.38头,随着寄生蜂日龄的增加,日平均产卵寄生量缓慢下降。日本细毛环腹瘿蜂平均单雌产卵寄生量为147.50头(图4A)。1~7日龄的日本细毛环腹瘿蜂雌蜂所产后代的性比较高,为77.78%~84.38%,性比变化平稳,9日龄的日本细毛环腹瘿蜂所产后代的性比急剧下降,11~15日龄的日本细毛环腹瘿蜂所产后代的性比为4.00%~10.00%,17~23日龄的日本细毛环腹瘿蜂所产后代只有雄性,无雌性后代(图4B)。
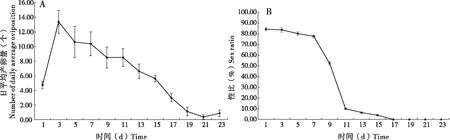
图4 日本细毛环腹瘿蜂日平均产卵寄生量(A)与后代性比(B)Fig. 4 Number of daily average oviposition (A)and offspring sex ratio (B)of Leptopilina japonica
2.4 日本细毛环腹瘿蜂不同发育阶段发育历期
日本细毛环腹瘿蜂是一种跨幼虫期、蛹期的跨期寄生蜂,日本细毛环腹瘿蜂的生长发育经历卵、幼虫、蛹和成虫4个发育阶段,卵期至蛹期均在斑翅果蝇体内发育,雌性日本细毛环腹瘿蜂将卵产在斑翅果蝇1~2龄幼虫体内,并在斑翅果蝇幼虫期孵化。日本细毛环腹瘿蜂幼虫期跨斑翅果蝇的幼虫期至蛹期,在实验室饲养条件下日本细毛环腹瘿蜂的卵期平均为1.30 d,幼虫期平均8.67 d,雄蜂蛹期平均为8.86 d,雌蜂蛹期平均为10.27 d,雄蜂的羽化时间比雌蜂平均提前1.41 d。
3 结论与讨论
日本细毛环腹瘿蜂成虫的寿命与补充营养关系密切,不同浓度的蜂蜜水及清水相对于无补充营养的对照来说,都能不同程度的延长成蜂寿命,且补充不同浓度蜂蜜水可显著延长成蜂寿命,其中20%蜂蜜水处理的寿命最长。这与其他种类寄生蜂的研究报道类似,补充营养对寄生蜂的寿命有重要影响,寄生蜂活动需要消耗大量的能量,通过摄取糖源营养物质获得能量,保证自身营养的消耗(杨琪,2017;李小敏等,2018)。Wang等(2018)在提供50%蜂蜜水与寄主的条件下,测得日本细毛环腹瘿蜂雌蜂寿命为18.7 d,低于本研究的25.75 d。这一差异可能是寄生蜂取食糖液浓度不同所致,高浓度糖液可导致虫体内渗透压升高,进而影响生理生活能力(李小敏等,2018),并且本研究是在无寄主幼虫的情况下测得其寿命,而寄生蜂对寄主搜寻与产卵都需要消耗更多的能量从而导致寿命减短。通过对日本细毛环腹瘿蜂行为活动节律观察得到,日本细毛环腹瘿蜂羽化行为与产卵行为呈现明显的昼夜节律,夜间无寄生蜂羽化。日本细毛环腹瘿蜂羽化期为8 d,第4天为羽化高峰期,羽化期每日10 ∶00-12 ∶00为羽化高峰期,其活动节律与黄色潜蝇茧蜂和哥德恩蚜小蜂表现相似(邹游兴,2012;杨琪,2017);每日16 ∶00-18 ∶00为产卵活动高峰期与蝶蛹金小蜂类似(江婷等,2018)。研究结果为日本细毛环腹瘿蜂保护利用提供了科学依据依据。在日本细毛环腹瘿蜂对不同寄主龄期非选择性试验中发现,日本细毛环腹瘿蜂不寄生斑翅果蝇 3龄幼虫,斑翅果蝇幼虫的龄期对日本细毛环腹瘿蜂的生长发育有一定的影响,日本细毛环腹瘿蜂对斑翅果蝇2龄幼虫表现出更高的适合度,斑翅果蝇2龄幼虫有利于日本细毛环腹瘿蜂的繁育。在日本细毛环腹瘿蜂对寄主龄期的选择性试验结果显示,日本细毛环腹瘿蜂表现出对斑翅果蝇 2龄幼虫的显著寄生偏好。Wang等(2018)研究发现,在人工饲料上,与3~4日龄斑翅果蝇幼虫相比,日本细毛环腹瘿蜂显著偏向于寄生早期1~2日龄的果蝇幼虫与本研究结果类似。日本细毛环腹瘿蜂从卵到成虫的发育历期为雌蜂20.17 d,雄蜂17.85 d,平均为19.57 d。Girod等(2018)的研究日本细毛环腹瘿蜂雌蜂发育历期31.0 d,雄蜂发育历期26.8 d,Wang 等(2018)的研究日本细毛环腹瘿蜂总体发育历期28.1 d。发育历期差异可能由于供试昆虫的地理种群不同所致,同种昆虫的不同地理种群的发育历期,寿命和产卵量等基本生物学特性存在一定的差异(朱文超等,2015)。日本细毛环腹瘿蜂雄蜂的羽化时间比雌蜂平均提前1.41 d,周思聪等(2018)研究同属的另一种Leptopilinaboulardi也得到了类似的结果。Wang等(2018)发现日本细毛环腹瘿蜂雌蜂在羽化后1~2 d内获卵量达到最大,王问学(1990)指出影响性比分配的主要因素有:雌蜂年龄、寄主的大小和质量好坏、雌蜂营养等。本研究中1~23日龄的日本细毛环腹瘿蜂都有产卵寄生行为,3日龄雌蜂的产卵寄生量达到高峰,随着雌蜂日龄的增加,雌性后代比例逐渐下降。斑翅果蝇存在较强的寄主免疫反应,在入侵地,有部分幼虫寄生蜂种类能在斑翅果蝇幼虫体内产卵,但是不能完成发育或者卵和幼虫被其免疫细胞包裹导致死亡(蔡普默等,2017;Alessiaetal.,2018)。本研究在对日本细毛环腹瘿蜂的产卵寄生动态研究中发现其能在斑翅果蝇幼虫体内完成发育且未发现卵和幼虫被免疫细胞包裹导致死亡的现象。由于地理种群不同,日本细毛环腹瘿蜂的发育历期和寄生不同寄主龄期幼虫表现出不同适合度(朱文超等,2015)。因此,利用日本细毛环腹瘿蜂防治斑翅果蝇实践应该注意选择合适的寄生蜂种群,放蜂时间应注意选择斑翅果蝇幼虫的发育龄期,以提高寄生蜂的寄生效率。
调查发现日本细毛环腹瘿蜂为斑翅果蝇优势寄生性天敌之一,对斑翅果蝇具有一定的控制潜能,本文对日本细毛环腹瘿蜂的生物学特性进行了观察研究,丰富和补充了前人的研究结果,为该蜂的保护利用提供重要的科学依据。对进一步深入研究该蜂的寄生行为、寄生率,对生态环境的适应性、控害效应和控害特性等奠定了基础,对斑翅果蝇综合防治、保护生态环境、减少农药的使用等方面有重要意义。
