樟子松人工林混交改造后土壤养分综合评价
2021-03-27张日升
石 亮,张日升
(辽宁省沙地治理与利用研究所,辽宁 阜新 123000)
科尔沁沙地位于内蒙古东南部西辽河中下游,总面积5.18万km2,是我国四大沙地之一。20世纪20—80年代,由于过度开垦、放牧、樵采等人类不合理利用自然资源活动[1],造成科尔沁地区疏林草原退化,土地沙漠化日趋严重,生态环境恶化[2]。目前,科尔沁地区以草地畜牧业发展为主的区域,植被处于逐渐恢复状态,而以种植业发展为主的区域,沙漠化逐渐加剧。国内学者认为,科尔沁沙地主要由固定沙地组成,范围内植被的水土保持功能仍然存在,地质构造上没有质的改变,有恢复生境的可能[3]。因此,科学合理的植被建设和防沙治沙措施,是推动科尔沁地区生态环境修复和区域经济可持续发展的关键因素。
樟子松(Pinussylvestrisvar.mongolica)具有耐寒耐旱、生态幅宽、适生地域广和生长周期长等优良特性,使其成为三北地区防护林重要组成树种[4]。但由于建植初期树种选择较为单一,且部分地区林木已经进入成、过熟期,生态功能和防护效益下降严重。科学合理的林分改造可延长樟子松人工林的生态防护周期,并使其向高功能地带性森林植被发展。Gadow等[5]提出了混交度的概念,并在此后的一段时间得到了广泛应用[6-9];高培军等[10]通过对11a生杉木拟赤杨混交林土壤理化性质的研究发现,各混交模式的水稳性团聚体、总孔隙度、通气度、持水量较纯林均有提高,尤其是土壤有机质更是提高了43.96%,说明混交林可以充分发挥水源涵养作用的同时,也能最大程度提升土壤肥力;杨菁等[11]应用高通量测序技术,比较了降香黄檀(Dalbergiaodorifera)不同混交林的土壤细菌多样性,发现黄檀×大果紫檀(Pterocarpusmacarocarpus)、黄檀×檀香(Santalumalbum)对土壤含水率、化学性质、酶活性、细菌丰富度和多样性的提升明显,pH值、多酚氧化酶及脲酶活性是主要影响因子。油松(Pinustabulaeformis)×刺槐(Robiniapseudoacacia)混交可增加土壤细菌、放线菌、真菌等微生物数量,提高土壤磷酸酶、脲酶、过氧化氢酶活性[12];侯本栋等[13]研究黄河三角洲刺槐混交林土壤养分变化时发现,有效磷、速效钾在5~20 cm土层低于刺槐纯林,除此之外的其他土壤养分指标均表现为混交林>纯林。目前,国内学者对樟子松开展了大量的生态学研究[14-15],但对科尔沁沙地樟子松人工纯林改造成针阔混交林后土壤养分变化规律和机制缺乏深入研究。本研究通过设置不同树种与樟子松人工林进行带状混交,分析各混交带内土壤化学性质和酶活性差异性,明确科尔沁沙地樟子松人工林混交后土壤养分变化规律和影响机制,从而为樟子松人工林更新改造及可持续经营提供理论依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于辽宁省阜新市彰武县章古台镇辽宁省沙地治理与利用研究所(42°43′~42°51′N,121°53′~122°22′E),海拔多在245m左右。属温带半湿润干旱区,年降水量480mm左右,多集中在6—8月,年平均气温6.82 ℃,多年平均蒸发量1 500mm,相对湿度60.4%,年均风速2.8 m/s。该地区主要分布的乔木树种有樟子松、赤松(Pinusdensiflora)、云杉(Piceakoraiensisi)家榆(Ulmuspumila)、银中杨(Populusalba)、色木槭(Acermono)、蒙古栎(Quercusmongolica)等;草本和灌木有马唐(Digitariasanguinalis)、虎尾草(Chlorisvirgate)、茵陈蒿(Artemisiacapillaris)、披碱草(Elymusdahuricus)等。
1.2 样品采集及测定
试验样地为典型的科尔沁沙地樟子松人工林,总面积33 hm2,林龄为38a生初始的栽植密度为1 600株/hm2。2012年进行带状采伐改造,间伐宽度分别为9 m,13 m[16]。2013年春季,在间伐带内分别栽植樟子松、银中杨、色木槭幼苗,形成樟子松×樟子松(P.sylvestrisvar.mongolica×P.sylvestrisvar.mongolica,PP)、樟子松×银中杨(P.sylvestrisvar.mongolica×Populusalba,PPM)、樟子松×色木槭(P.sylvestrisvar.mongolica×Acermono,PAM)混交林,并以沙地樟子松纯林为对照。2018年9月,在海拔相近的样地内设置3个面积为50m2的样方,用土钻(Φ=5 cm)按“S”形在0~20 cm土层取样,每个土样重量约1 kg并带回实验室。土壤有机质采用重铬酸钾氧化法、碱解氮采用碱解扩散法、有效磷采用NaHCO3-钼锑抗比色法、过氧化氢酶采用高锰酸钾滴定法、蔗糖酶采用二硝基水杨酸比色法、磷酸酶采用苯磷酸二钠法、脲酶采用苯酚钠-次氯酸钠比色法。样地基本情况如表1所示。
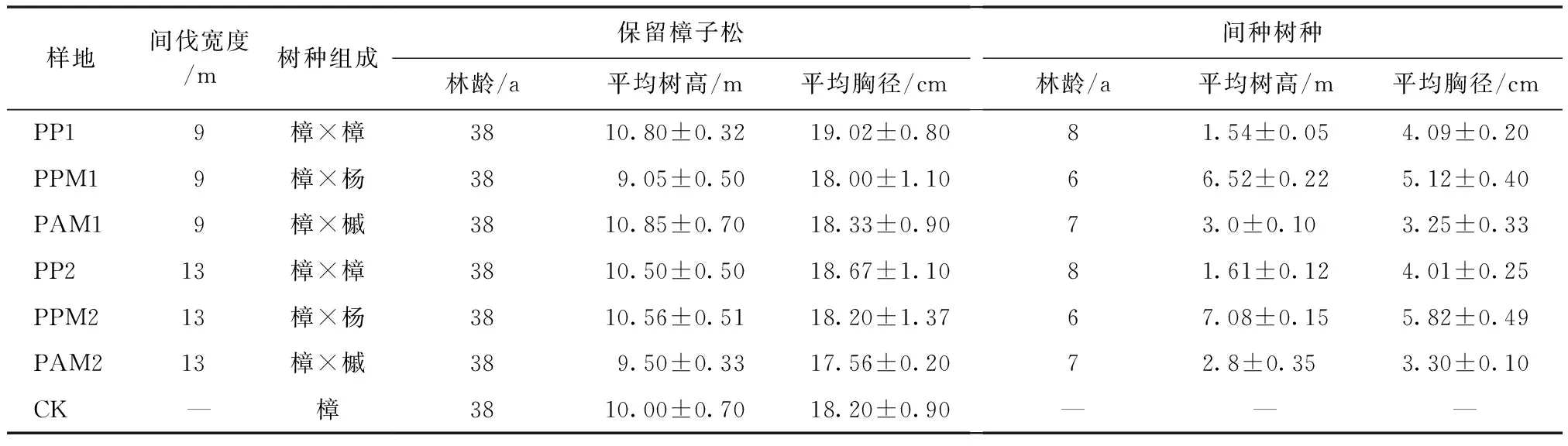
表1 样地基本概况
1.3 数据处理
本研究混交样地土壤养分差异性采用SPSS软件的单因素方差(One-way ANOVA)和并运用Duncan检验法进行多重比较(α=0.05)。采用两个样本T检验方法分析PP,PPM,PAM不同栽植宽度的差异。采用典范对应分析和主成分分析法对不同混交模式土壤性状进行综合评价。运用Canoco 4.5软件对不同样地各土壤性状因子进行典范对应分析(Canonical Correspondence Analysis)。采用CanoDraw,Origin95进行绘图制作。
2 结果与分析
2.1 混交改造对土壤化学性质的影响
混交改造显著改变了樟子松人工林的土壤化学性质。由图1可知,大部分样地土壤有机质、碱解氮、有效磷均显著高于CK,其中有机质含量最高为PAM2样地,碱解氮含量最高为PPM2样地,有效磷含量最高为PP1样地,三者分别较CK增加90.88%,159.07%,87.58%。除上述样地外,其他混交样地土壤化学性质之间并无显著差异(P>0.05)。通过样本T检验可知,不同间种宽度,相同间种树种的样地中,PPM2样地碱解氮含量显著高于PPM1,PAM2样地有机质、有效磷含量显著高于PAM1(P<0.05)。

2.2 混交改造对土壤酶活性的影响
混交改造对樟子松人工林土壤酶活性影响不同。
由图2可知,4种酶活性峰值分别出现在PPM1,PPM2,PPM2和PPM1样地,均为樟×杨混交样地。与CK样地相比,各样地土壤酶活性均呈现不同程度提高,其中磷酸酶和脲酶活性提高较为显著(P<0.05),但混交样地之间的差异性不显著(P>0.05),与土壤化学性质表现类似。通过样本T检验可知,只有PPM2样地蔗糖酶活性显著高于PPM1,说明本研究酶活性出现差异性主要受混交树种影响,而非间种宽度。

2.3 各混交模式土壤养分因子综合评价
从图3样点分布可以看出,碱解氮、蔗糖酶处于第一象限,与第一、二序列轴呈正相关关系;有效磷处于第二象限,与第二序列轴呈正相关关系;有机质、过氧化氢酶、磷酸酶、脲酶处于第四象限,与第一、二序列轴呈负相关关系。碱解氮、蔗糖酶与PPM2相关性程度较大,有效磷与樟×樟样地相关性程度较大,有机质、过氧化氢酶、脲酶、磷酸酶与PPM1,PAM1,PAM2相关性程度较大。

注AP为有效磷;AN为碱解氮;INV为蔗糖酶;OM为有机质;PHO为磷酸酶;CAT为过氧化氢酶;URE为脲酶。
为评估混交改造对樟子松人工林土壤养分的影响,对所有土壤养分因子(有机质、碱解氮、有效磷、过氧化氢酶、蔗糖酶、磷酸酶、脲酶)进行离差标准化处理,并进行主成分分析。由表2可见,综合得分由高到低依次是PPM2>PPM1>PAM2>PP1>PP2>PAM1>CK样地。本研究各处理提取前3个主成分的累计贡献率为90.80%,其中F1=0.85X1+0.89X2+0.60X3+0.88X4+0.75X5+0.88X6+0.82X7,权重为0.73;F2=0.01X1-0.16X2+0.65X3+0.22X4-0.43X5+0.40X6+0.29X7,权重为0.15;F3=-0.16X1+0.29X2+0.47X3-0.32X4+0.44X5-0.23X6-0.30X7,权重为0.12。

表2 土壤养分因子综合评价
3 讨论与结论
3.1 讨论
长期实施林分改造能够有效改良林下土壤养分状况,提高森林生态服务功能[17]。对西伯利亚红松进行混交林分改造后,由于地表凋落物的分解和地下根系的周转发生改变,增强了有机质质量分数的变异的程度[18]。李丽红[19]通过对10a生马尾松大叶相思混交林土壤肥力的研究发现,马尾松大叶相思单行混交的土壤有机质含量较马尾松纯林增加59%。本研究各混交样地的土壤有机质含量均显著高于对照,主要是由于土壤碳库储备主要来自于植物残体和凋落物,针阔混交可丰富碳素的来源途径,阔叶林有机碳储量高于针叶林[20],加之适宜的水热条件,促使碳素的养分归还速率加快。除PPM2样地外,其他樟×杨、樟×槭样地土壤碱解氮含量与樟×樟样地无显著差异,甚至含量更低,土壤有效磷含量也有相似的规律。雷静品等[21]在研究带状改造对柏木林土壤性质的影响发现,柏木林混交刺桐、刺槐后碱解氮含量低于柏木纯林,与本研究结果相近。这可能是由于种群生态位关系造成不同树种的竞争,阔叶幼树从土壤中汲取更多的氮、磷元素来满足生长需求。磷素作为科尔沁地区沙地的主要限制因素,在土壤中易形成难溶性盐沉淀[22],阔叶幼树根系需要更强的活化作用来吸收磷素。
土壤酶是土壤物质循环和能量转化的重要媒介,是反应土壤质量和健康较为敏感的生物指标[23]。本研究中各混交样地的土壤过氧化氢酶活性均高于对照,可能是由于研究区范围内土壤类型以沙土为主,土壤条件受外部环境变化影响明显,混交改造丰富了林内物种多样性,土壤动物、微生物残骸、植物根系分泌等使得土壤环境得到改善[24],提高了土壤的生物化学活性,说明樟子松混交林较樟子松纯林有更强的抵御过氧化氢毒害的能力 。针阔混交样地磷酸酶活性显著高于对照(P>0.05),说明樟子松纯林经过混交改造有利于林内植被生长及土壤中微生物的生物活动,加快有机磷的水解和循环速度,提高磷的利用效率。各样地土壤蔗糖酶、脲酶活性均高于对照,但混交改造样地之间无显著差异(P<0.05),这可能一方面是由于混交改变了原有植被类型,凋落物分解条件和土壤微生物状况都发生了不同程度的变化[21],需要更长的时间来重新构建新的分解系统;另一方面是由于沙土保水蓄水能力差,使得酶活性变弱,氮素流失或没有完全释放。
运用典范对应分析可知,PP1,PP2样地处于第二象限,与有效磷存在显著关系,说明樟×樟样地相较于针阔混交林更有利于土壤磷素的积累。第一序列轴由左向右,土壤有效磷含量逐渐降低,磷酸酶活性逐渐升高,一方面,说明樟子松人工林混交改造有利于提高土壤酶活性,另一方面,说明混交改造中的阔叶幼树为满足自身生长需求,在磷酸酶活性的作用下,会消耗一部分土壤中的磷元素。由于科尔沁沙地土壤磷含量较低[25],且磷元素易于流失,有机磷矿化速率慢且有效性差[26],说明磷元素是影响樟子松人工林改造效果的关键因素。PPM2,PPM1样地分别位于第一、四象限,第二序列轴自下而上间伐宽度逐渐增大,相关的土壤养分因子由有机质、过氧化氢酶、脲酶、磷酸酶变为碱解氮、蔗糖酶,说明杨树对外界环境的变化较为敏感,适宜的间种带宽使林内水热条件向更利于幼树生长的方向变化,进而提高了土壤中微生物的活动频率,动植物残体及凋落物的死亡分解和淋溶作用加强,有利于土壤碳库、氮库、磷库储备的增加。土壤酶活性显著提高,尤其是磷酸酶活性的增加可增加磷素的矿化和转化速率,一定程度上缓解了科尔沁沙地磷素的限制。
目前,国内关于林分改造、异龄混交的研究较多,如冀北山地华北落叶松人工林与白桦混交经营后,6 年的短期异龄混交改造使林内土壤动物显示出较高的多样性水平[27]。曹小玉等[28]通过对湖南地区杉木改造土壤肥力的研究,认为改造后 3 个龄组杉木生态公益林土壤养分含量都有不同程度提高,综合评价指数提高。不同的混交树种对土壤的养分的影响也不尽相同,葛乐等[29]通过对杉木林套种12种植物发现,在种植初期麻栎和毛红椿改良效果最佳。但是温远光等[30]发现,马尾松与红锥异龄混交林土壤微生物群落多样性和土壤酶活性明显低于马尾松纯林,提出应重视天然更新的观点。本研究表明,针阔混交林对栽植空间的响应更加敏感,通过主成分分析樟子松人工林混交改造对土壤化学性质、酶活性的特征发现,各样地得分由高到低依次为PPM2(0.84)>PPM1(0.41)>PAM2(0.29)>PP1(0.27)>PP2(0.09)>PAM1(-0.42)>CK(-1.49)。总的来说,混交样地的土壤养分因子综合得分高于对照,针阔混交林普遍高于樟子松异龄林。因此,科学合理地选择乡土树种和栽植空间,将樟子松人工林改造为针阔混交林,可有效促进土壤生态改善,延长防护林的生命周期。此外,土壤磷是影响樟子松人工林改造最重要的因子(图3),同时也是樟子松成熟林生长主要的限制因子[26]。固氮植物有助于提高土壤磷酸酶活性、增加磷的循环速率[31],从而提高土壤中氮、磷的有效性[32]。在混交林建植初期,建议增施磷肥或在间伐林带间种植固氮植物,以提供充足的养分供给阔叶植被幼苗生长,保证樟子松人工林的可持续发展。
3.2 结论
1)混交改造可有效改善土壤化学性质,大部分样地土壤有机质、碱解氮、有效磷含量显著提高,但混交样地之间差异性不明显。
2)针阔混交样地土壤酶活性高于樟×樟,二者均高于对照,过氧化氢酶、蔗糖酶、磷酸酶和脲酶活性的峰值均出现在了樟×杨样地。
3)针阔混交林综合得分较高,表现最好的是樟×杨样地,可作为科尔沁沙地樟子松人工林改造的首选模式。在混交林建植初期,应适当增施磷肥或在间伐林带间种植固氮植物。
