濒危植物九龙山榧与其同属广布种榧树叶绿素荧光特性的比较
2021-03-27郑英茂周钰鸿潘成椿刘菊莲蓝本宽康华靖
郑英茂,周钰鸿,潘成椿,刘菊莲,蓝本宽,康华靖
(1.浙江九龙山国家级自然保护区管理局,浙江 遂昌 323300;2.浙江大盘山国家级自然保护区管理局,浙江 磐安322300;3.遂昌县自然资源和规划局,浙江 遂昌 323312;4.温州市农业科学研究院,浙江 温州 325006)
植物濒危过程是该物种生活史各个环节及其竞争力、适应力等与环境因子相互作用的生态学过程,因此有必要从植物生活史的不同侧面探求其濒危的原因。研究表明,大多濒危植物的濒危机制与其对立地条件,如光、水、肥、气、热等因子的要求密不可分,而光合作用对植物生长发育起着至关重要的作用[1-3]。因此对植物光合生理生态特性进行研究,可阐明植物生存及其分布的内在机制[4],从而为制定保护方案提供科学依据。濒危植物在生理生态学方面一般存在生存力和适应力较差的共性[5]。在相同条件下,多数濒危植物的光合、呼吸和蒸腾等生理代谢速率比其非濒危近缘种要低[6-9],如,柴胜丰等[9]研究表明,与茶(Camelliasinensis)相比,毛瓣金花茶(C.pubipetala)对光强的适应范围狭窄,并推测这可能是其分布狭窄的重要生理原因。因此,研究濒危植物的光合特征,并与其广布近缘种进行比较,从而了解两者在光合生理生态特征上的差异,对于揭示濒危植物的濒危机制至关重要[10]。然而,目前相关研究往往只在野生或栽培两种生境之一下进行,并不能反映植物在变化环境中的适应能力,如能同时在野生和栽培条件下分别比较濒危种和广布近缘种的光合特性的差异,研究结果将更有说服力。
榧属(Torreya)植物为红豆杉科(Taxaceae)的常绿乔木。榧属为古老珍稀的孑遗植物,具有重要经济、科学研究价值,全部野生种(包括变种)均被列为国家二级保护植物[11]。九龙山榧是1995年发现的形态界于长叶榧和榧树之间的一个类群,当时作为榧树的变种T.grandisvar.jiulongshanensis命名发表,后被提升为种级T.jiulongshanensis,此后在天台、磐安、丽水莲都和松阳有少量发现[12]。总之,九龙山榧分布范围窄,数量极少。
九龙山榧如此狭窄的地理分布必然与其自身的生理生态特性有关,而至今有关九龙山榧生理生态特性的研究尚未见报道。为此,本文以濒危植物——九龙山榧(雌、雄株),及其广布近缘种——榧树为研究对象,分别对野生和栽培环境下两者的光合生理特性进行比较,以期为揭示九龙山榧濒危机制及进一步保护提供科学依据。
1 研究方法
1.1 研究概况
研究所在地分别位于浙江省丽水市遂昌县西南的龙洋乡西坑下村(N28.337°,E118.933°)和浙江九龙山国家级自然保护区的泗州庙(N40.542°,E116.118°)。该地区气候属于中亚热带湿润季风气候,四季分明、雨水充沛、光照适宜,相对湿度较高。西坑下村附近的榧树群落中,共有榧树和九龙山榧共计20余株,树龄均介于400~500 a生;泗州庙分布处1株九龙山榧雌株。自1995年发现以来,当地政府一直较为重视对九龙山榧的保护。2015年对西坑下村群落进行了保护性改造,同时在雌株附近搭建了塔台,以便观测研究。
1.2 试验设计与数据测量
由于九龙山榧雌雄异株,叶片的大小有明显区别。因此,在测量时,九龙山榧雌雄株均分开测量,单独记录。另外,在试验同时,比较了栽培条件下榧树、九龙山榧(雌、雄)幼苗叶片的光合特征。榧树实生苗于2015年2月播种,以作为砧木和幼苗对照备用。接穗采自于县龙洋乡西坑下村的雌雄母株。2018年3月中旬嫁接,嫁接后置于西畈乡繁殖基地的塑料大棚中常规管理养护,夏季使用遮阳网遮阳,透光率为50%左右。
试验均于2018年9月上旬晴天的9:00~11:30进行。野外条件下,分别在九龙山榧母树的雌雄株上各选取长势较为一致的7张叶片待测(每株树上选取不同方位的叶子2~3片),所选叶子均为2a生枝条。由于受野外限制,选取同群落中榧树雄株作为对照;栽培条件下,分别选取长势一致的嫁接幼苗植株雌雄各7株,每株上选取顶部当年生枝条2片叶子待测,未嫁接的实生榧树苗作为对照(由于榧树较小,尚无法区分雌雄株)。所选叶子均为当年生枝条上完全发育成熟且健壮的叶片。由于树的高度及野外条件限制,野外母株的数据均利用便携式光合仪(LI-6400-40,LI-COR INC.,USA)进行连体测量,或利用搭建的观测台架,或借助地势结合三脚架进行。
测量前进行开机检查及预热,流速为500 μmol/s,CO2浓度设置为400 μmol/mol,由内置CO2小钢瓶提供。由于叶片革质且较小,难以充满其叶室,试验只测量其快速光曲线(不考虑其气体交换参数)。光合有效辐射强度(PAR)设置为1 800,1 600,1 400,1 200,1 000,700,400,200,150,100和50 μmol/(m2·s),并应用自动测量程序自动记录荧光参数,由此得到光系统II(PSII)反应中心的激发能捕获效率(Fv'/Fm')、PSII实际的光化学量子效率(ΦPSII)、光系统II的电子传递速率(ETRII)、光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)等参数对光强的响应曲线。每次记录最小等待时间为60 s,最大等待时间为90 s[13]。
幼苗的测量均采用Dual-PAM-100(WALZ,德国)双通道叶绿色荧光仪进行数据测量,由此得到光系统I(PSI)的电子传递速率(ETRI)、PSI最大荧光(P'm)等荧光参数。仪器开机预热15 min后测量其快速光曲线。由于大棚有遮阳,光强相对较弱,且预试验结果表明,在较高光强下,电子传递速率值出现异常。因此,光合有效辐射强度(PAR)设置为10,18,36,94,172,214,330,501和759 μmol/(m2·s)(仪器中设置档位所对应的光强)。测量其快速光曲线,同时获得光系统I(PSI)和光系统II(PSII)的相关荧光参数。
1.3 数据处理
运用光合计算软件4.1(井冈山大学生命科学学院/数理学院),分别应用光响应机理模型(model I)、直角双曲线模型(model II)、非直角双曲线模型(model III)和双指数方程模型(model IV)拟合光系统II电子传递速率(ETRII)对光强的响应曲线。4种模型的表达式参考叶子飘[14]的描述,除光响应机理模型外,其余模型拟合的饱和光强(PARsat)计算参考叶子飘[14]的方法。SPSS12.5版本(SPSS,Chicago,IL)软件进行计算、处理和方差差异显著性分析,用Excel 2010作图。
2 结果与分析
2.1 栽培条件下九龙山榧幼苗和榧树幼苗的荧光参数对光强的响应
2.1.1光系统I(PSI)荧光参数对光强的响应

图1 榧树和九龙山榧(雌、雄)叶片P'm和ETRI对光强的响应
由图1(a)可以看出,九龙山榧(雌、雄株)和榧树的幼苗PSI最大荧光(P'm)整体随着光强的增加而升高,在光强大于500 μmol/(m2·s)之后趋于稳定。比较P'm值可以看出,九龙山榧雌株幼苗叶片的P'm值整体上显著高于榧树及九龙山榧的雄株(P<0.05)。由图1(b)可以看出,随着光强的升高,九龙山榧(雌、雄株)幼苗叶片的ETRI均呈增加趋势;而榧树幼苗叶片的ETRI则呈先上升后下降的趋势,在330 μmol/(m2·s)光强下达到最高。两者表现出了较大差异。
2.1.2光系统II(PSII)荧光参数对光强响应
由图2可以看出,在较低光强(小于214 μmol/(m2·s))下,九龙山榧(雌、雄株)和榧树幼苗叶片的PSII电子传递速率(ETRII)均呈直线上升趋势;之后,其增加相对平缓而趋于稳定。从图2还可以看出,在较低光强(小于214 μmol/(m2·s))下,九龙山榧和榧树幼苗叶片的ETRII均较为接近;而在较高光强下,九龙山榧幼苗(雌、雄株)叶片的ETRII值相对较高。

图2 榧树和九龙山榧(雌、雄)叶片光系统II电子传递速率对光强的响应
以榧树幼苗数据为例,应用4种光响应模型对其ETRII对光强的响应数据进行拟合,其拟合结果如图3所示。从4种光合模型拟合的效果图来看,除直角双曲线模型(model II)不能很好拟合高光强下电子传递速率下降的趋势外,其余模型均能较好拟合测量值的变化趋势。从确定系数(R2)来看,4种模型的确定系数均高于0.9(表1)。

注“О”为测量值;“×”为光响应机理模型(model I);“﹡”为直角双曲线模型(model II);“●”为非直角双曲线模型(model III);“◆”为双指数方程模型(model IV)

表1 光系统II电子传递速率对光响应曲线的拟合结果(平均数±标准误差)
从拟合得到的最大电子传递速率来看,除直角双曲线模型(model II)外,其余光响应模型拟合给出的值均与实测值较为吻合(P>0.05)。然而,从拟合得到的饱和光强值来看,4种模型拟合的效果存在较大差异。整体来看,光响应机理模型(model I)拟合得到的饱和光强值远高于其他模型所给出的值(P<0.05),且与实测值较为接近。由此判断,光响应机理模型(model I)对榧树和九龙山榧的拟合效果较好。从model I拟合结果来看,榧树、九龙山榧雄株和九龙山榧雌株幼苗的光饱和点分别为454.130,550.640和788.581 μmol/(m2·s),所对应的最大电子传递速率分别为35.339,39.546和40.690 μmol/(m2·s)。
由图4(a)可以看出,榧树和九龙山榧(雌、雄株)幼苗叶片光化学猝灭系数(qP)随着光强的升高而呈线性下降,三者之间的qP值均较为接近(P>0.05)。由图4(b)可以看出,榧树和九龙山榧(雌、雄株)幼苗叶片非光化学猝灭系数(NPQ)随着光强的升高而增加,其中,九龙山榧雌株幼苗的NPQ值整体较高,在较高光强下则显著高于其雄株及榧树(P<0.05),而九龙山榧雄株和榧树之间的NPQ整体较为接近。
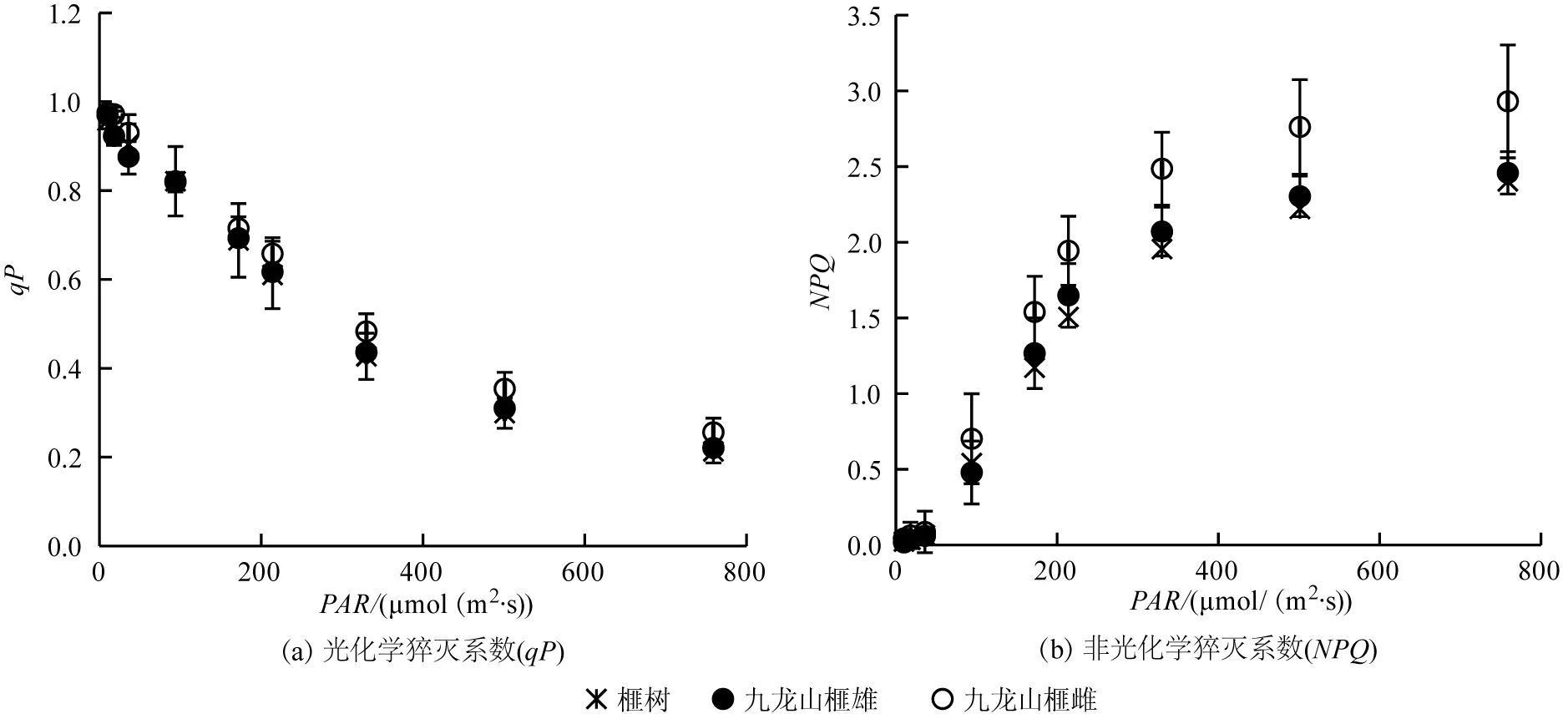
图4 榧树和九龙山榧(雌、雄株)叶片光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)对光强的响应
2.2 野生条件下九龙山榧和榧树叶片的荧光参数对光强的响应
2.2.1野生条件下九龙山榧(雌、雄株)和榧树叶片ETRII对光强的响应
利用不同光响应机理模型拟合野外测量的榧树和九龙山榧(雌、雄株)叶片ETRII对光的响应曲线,拟合效果结果如图5所示(以榧树数据为例)。从拟合效果来看,所选的4种模型均能较好地拟合榧树叶片ETRII对光的响应曲线。

注“О”为测量值;“×”为光响应机理模型(model I);“”为直角双曲线模型(model II);“●”为非直角双曲线模型(model III);“◆”为双指数方程模型(model IV)
以光响应机理模型拟合结果为例,对榧树和九龙山榧(雌雄)进行比较(表2)。由表2可以看出,九龙山榧雌株的最大电子传递速率为80.322 μmol/(m2·s),显著较高(P<0.05);九龙山榧雄株和榧树的最大电子传递速率较为接近(P>0.05),分别为66.308和64.743 μmol/(m2·s)。从饱和光强来看,九龙山榧雄株的光饱和点最高,为1 416.817 μmol/(m2·s),其次为九龙山榧雌株,榧树则最低(P<0.05)。

表2 利用光响应机理模型(model I)拟合ETRII 对光响应曲线的拟合结果(平均数±标准误差)
2.2.2野生条件下九龙山榧(雌、雄株)和榧树叶片qP和NPQ对光强的响应
由图6(a)可以看出,野生条件下榧树和九龙山榧(雌、雄株)母株叶片qP随着光强的升高而呈线性下降,三者之间的qP值均较为接近(P>0.05)。由图6(b)可以看出,当光强较低时(小于400 μmol/(m2·s)),野生条件下榧树和九龙山榧(雌、雄株)母株叶片NPQ随着光强的升高而增加;当光强大于400 μmol/(m2·s)时,随着光强的升高,九龙山榧树(雄株)和榧树叶片的NPQ基本保持稳定,且两者极为接近,而九龙山榧树雌株则在 1 000μmol/(m2·s)时达到稳定,且NPQ值显著较高(P<0.05)。

图6 榧树和九龙山榧(雌、雄株)叶片光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)对光强的响应
3 讨论
叶绿素荧光动力学技术作为研究植物光合功能快速、无损伤的探针,在测定叶片光合同化过程中,光系统对光能的吸收、传递、耗散、分配等方面具有显著的优势。因此,本试验基于荧光参数对九龙山榧及其同属广布种——榧树的光合生理进行探讨。
了解植物叶片的光饱和点对于深入探讨其最大羧化速率等光合参数,以及指导植物的引种栽培等均具有重要意义。目前,确定植物的光饱和点主要应用光响应模型对光响应曲线进行拟合而得到[15-16]。由于模型的推导机理不同,不同模型存在一定的优缺点[15,17]。本试验运用了4种模型对野外和栽培条件下九龙山榧及榧树叶片的快速光曲线进行了拟合。从拟合给出的光饱和点来看,光响应机理模型(model I)的拟合结果最为接近测量值。以model I拟合的结果为例,栽培条件下,榧树、九龙山榧雄株和九龙山榧雌株幼苗的光饱和点分别为454.130,550.640和788.581 μmol/(m2·s);而野外环境下,榧树、九龙山榧雄株和九龙山榧雌株的光饱和点分别为914.207,1 416.817和1 165.361 μmol/(m2·s)。由此可见,不管是栽培条件下的幼苗,还是野外的母树,榧树的光饱和点均显著较低(P<0.05)。结合野外观察,推测与榧树叶片叶绿素含量较低相关,与榧树相比,九龙山榧植株叶片叶色整体更加亮绿、叶片生长势更加旺盛。由此可见,九龙山榧为喜光的阳生树种。
从最大电子传递速率的拟合结果来看(以model I拟合的结果为例),栽培条件下,九龙山榧(雌、雄株)整体上显著高于榧树(表1);而在野外条件下,九龙山榧雌株显著高于榧树(表2)。这与以往研究认为濒危植物光合能力往往低于其同属广布种的结果有所出入,如,石松利等[8]研究表明,濒危植物四合木(Tetraenamongolica)的光合速率和水分利用效率均低于其近缘种霸王(Zygophyllumxanthoxylon),光合能力和对干旱环境适应能力弱于霸王;Cui等[7]研究同样表明,濒危种Syringapinnatifolia的净光合速率和对CO2的吸收能力均低于其同属广布种S.Oblata。由此表明,光合能力并非九龙山榧的致濒因子。推测九龙山榧的数量较少与其结实率极低有直接关系。多年的野外观测表明,九龙山榧的结实率远低于同群落中的榧树。
光化学淬灭系数(qP)反映PSⅡ天线色素吸收的光能用于光化学电子传递的份额[18-19]。植物吸收的光能一部分除用于光合作用外,还有一部分可通过非光化学猝灭(NPQ)过程来调整过量能量的耗散,保护PSⅡ反应中心免受因吸收过多光能而引起的光氧化和光抑制伤害[20]。本试验结果表明,除九龙山榧雌株(幼苗和母株)的NPQ值均相对较高外,九龙山榧雌雄株以及榧树之间的qP和NPQ整体上极为接近。
另外,研究发现多种植物雌雄异株之间对水分[21]、温度[22-23]、养分[24]、CO2浓度[25]等环境变化的响应差异显著。综合比较九龙山榧雌雄株的荧光特性发现,九龙山榧雌株幼苗的光饱和点、最大荧光参数(P'm)和NPQ值均显著较高(P<0.05);同样,九龙山榧雌株母树的最大电子传递速率也显著较高(P<0.05)。从光系统I(PSI)值来看,九龙山榧雌株整体上也显著高于榧树及九龙山榧的雄株(P<0.05)。作者推测,这与九龙山榧雌株的叶片相对较大,长势相对旺盛有关。然而,九龙山榧雌雄株之间差异的原因尚有待于进一步研究。
