不同耕作措施下添加秸秆对土壤有机碳及其相关因素的影响
2021-03-25王碧胜于维水武雪萍高丽丽李景宋霄君李生平卢晋晶郑凤君蔡典雄
王碧胜,于维水,武雪萍,高丽丽,李景,宋霄君,李生平,卢晋晶,郑凤君,蔡典雄
不同耕作措施下添加秸秆对土壤有机碳及其相关因素的影响
王碧胜1,2,于维水2,武雪萍2,高丽丽3,李景4,宋霄君2,李生平2,卢晋晶2,郑凤君2,蔡典雄2
1青岛农业大学农学院,山东青岛 266109;2中国农业科学院农业资源与农业区划研究所,北京 100081;3中国农业科学院农业环境与可持续发展研究所,北京 100081;4河北地质大学水资源与环境学院,石家庄 050031
【】探究添加秸秆对不同耕作措施下土壤有机碳及其相关因素的影响,为北方旱作农田固碳增产管理提供理论依据。采集长期进行传统耕作(CT)和免耕(NT)的大田土壤样品进行室内培养试验,共设置4个处理,分别为传统耕作土壤不加秸秆(CT)、免耕土壤不加秸秆(NT)、传统耕作土壤加秸秆(CTS)和免耕土壤加秸秆(NTS),每个处理15次重复。在25℃恒温培养箱中进行通气培养,培养时间共180 d,此间定期取样进行有机碳含量、水稳性团聚体构成、土壤微生物量碳和相关土壤酶活性的测定。(1)添加秸秆显著提高土壤有机碳含量和大团聚体含量。与CT相比,CTS提高土壤有机碳含量15%—46%;与NT相比,NTS提高土壤有机碳含量12%—21%;培养结束时,CTS、NTS处理的有机碳含量较初始分别提高26.8%和7.0%。CTS和NTS处理以2 000—250 μm团聚体含量最高,占全部团聚体的41%—50%,CTS较CT提高>250 μm团聚体比例235%—310%,NTS较NT提高>250 μm团聚体比例96%—149%。(2)添加秸秆显著增加土壤有机碳δ13C值,CTS处理为80.93‰—115.22‰,NTS为48.92‰—80.49‰;CTS秸秆来源碳所占比例显著高于NTS,较NTS处理提高13%—66%。(3)添加秸秆显著提高微生物量碳(MBC)含量、β-葡萄糖苷酶(BG)、β-纤维二糖苷酶(CBH)和β-木糖苷酶(BXYL)活性。CTS较CT提高MBC含量239%—623%,提高BG、CBH和BXYL活性58%—170%、52%—337%和117%–170%;NTS较NT处理提高MBC含量124%—555%,提高BG、CBH和BXYL活性28%—181%、4%—304%和13%—118%。(4)土壤有机碳含量与BG、CBH和BXYL活性、MBC及>2 000 μm、2 000—250 μm团聚体比例呈显著正相关关系,与250—53 μm、<53 μm团聚体比例呈显著负相关关系;BG、CBH、BXYL 3种酶活性彼此之间表现为极显著正相关关系,且均与MBC、>2 000 μm团聚体、2 000—250 μm团聚体显著正相关,与<53 μm团聚体极显著负相关。线性相关分析结果表明水稳性大团聚体(>250 μm)可解释有机碳变化的48%,MBC可解释有机碳变化的45%,BG、CBH和BXYL酶活性分别可解释有机碳变化的66%、44%、53%。添加秸秆可显著提高土壤有机碳和大团聚体含量,促进微生物数量增加和土壤酶活性增强,且对传统耕作土壤有机碳及其相关因素的影响更大,有机碳在土壤中的固定除了受团聚体物理保护外,还受土壤中微生物作用的调节。
团聚体;有机碳;δ13C;土壤酶;添加秸秆;耕作措施
0 引言
【研究意义】土壤有机碳(SOC)是评价土壤肥力的重要指标[1],同时由于土壤有机碳库含量巨大,其微小变化即可对大气二氧化碳(CO2)浓度产生剧烈作用[2],因此,SOC不仅对土壤质量和作物产量有直接影响[3],而且在环境保护、农业可持续发展等方面均起着极其重要的作用[4],确保农业生态系统中SOC的永续性具有重要意义[4-5]。我国玉米秸秆资源数量大、分布广[6],但多被农民采取就地燃烧的方法处理,这不仅会增加CO2排放,造成环境污染,而且不利于农业可持续发展[7]。山西省是全国玉米秸秆主产区,秸秆中有机物含量高,是重要的生物质资源[8],将其还田能够有效增加SOC含量、提高土壤肥力[9-10]。因此,探究添加秸秆对SOC的影响及其机理研究既具有理论价值又有现实价值。【前人研究进展】相关研究表明,如果每年0—30 cm土层中有机碳储量增加4‰,即可抵消当年CO2排放量[11]。ZHANG等[12]通过长期定位试验研究得出在中国北方秸秆还田可增加SOC储量0.07—1.461 t·hm-2·a-1。目前关于有机碳固存的机制主要包括化学保护机制、物理保护机制和生物化学稳定机制[13-14]。有研究表明外源有机物料的施用促进水稳性团聚体的稳定性,微团聚体内有机碳的降解过程耗能较大,受到物理保护作用较大,所以微团聚体中有机碳含量的提高有助于SOC的持久稳定[15-16]。刘哲等[16]的研究表明添加秸秆显著提高了>2 000 μm和2 000—250 μm粒级水稳性大团聚体有机碳的贡献率,从而加强了有机碳在土壤中的稳定性及固持能力。由上述研究可知添加秸秆能够有效提高SOC含量。【本研究切入点】目前关于秸秆增加SOC的原因多归于物理保护作用,且研究对象主要为单一土壤添加秸秆与不加秸秆处理。耕作和秸秆还田是驱动农田土壤碳库周转的关键因素[17-18],但关于长年经过不同耕作处理的土壤添加秸秆的研究相对较少。此外,土壤微生物特性,如酶活性、微生物丰度和群落组成等,已被广泛用于评估秸秆还田的影响[19],为探究添加秸秆提高有机碳的机制扩宽了思路。【拟解决的关键问题】本文以长期保护性耕作农田土壤为基础,采用室内培养方法,旨在明确添加秸秆对传统耕作和免耕土壤SOC、团聚体分布、微生物生物量碳及相关酶活性的影响;同时利用13C标记技术定量分析秸秆碳对SOC的影响,结合物理、生物两方面阐明添加秸秆增碳的机制,为北方旱作农田固碳增产管理提供理论依据。
1 材料与方法
1.1 试验区概况
试验区位于山西省寿阳县宗艾村(东经112°— 113°,北纬37°—38°),年平均气温7.4℃,年平均降雨量为461.8 mm,属中纬度暖温带半湿润偏旱大陆性季风气候区。试验地点选在地势较平缓的褐土上,质地为砂质壤土,属于全年无灌溉雨养地。初始时0—20 cm土层基础理化性状如表1所示。大田试验开始于2003年,每年保持相同处理,设置传统耕作(CT)和免耕(NT)两种耕作处理,具体操作为:CT处理秋收后秸秆移出,秋季耕翻,春季播前撒施化肥并春耕;NT处理秋收后将秸秆顺行推倒免耕覆盖,春季免耕,顺行开5 cm深小槽,点播玉米种子,在两播种点之间穴施化肥。种植作物为春玉米,一年一作,供试玉米品种为当地优势品种。试验小区面积为5 m×5 m=25 m2,重复3次,各处理N、P2O5施肥量均为105 kg·hm-2。
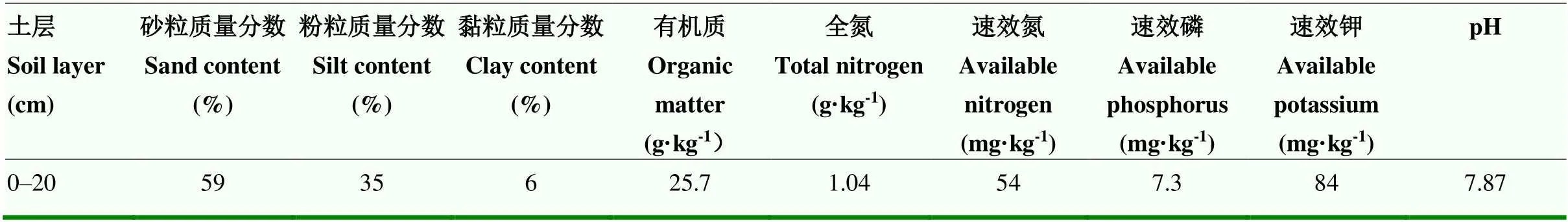
表1 试验地 0—20 cm土层基础理化性状
1.2 供试材料
2016年(试验开始后第14年)秋季收获后,采集0—20 cm土层原状土样和混合土样。原状土样带回实验室,在通风阴凉处自然风干后沿土块天然断裂面轻轻掰开,将大的植物残体和石子砂砾去除,通过6 mm筛子,用于测定初始土样水稳性团聚体。混合土样分为两部分,一部分迅速放入保鲜箱中带回实验室,过2 mm筛后放入-20℃冰箱中保存,用于测定微生物生物量碳和土壤酶活性;另一部分风干后通过2 mm筛子,用于室内培养试验[20]。CT和NT土壤培养前初始养分含量分别为:有机碳21.65和28.51 g·kg-1,全氮1.05和1.55 g·kg-1,速效磷12.0和46.3 mg·kg-1,速效钾106.0和173.0 mg·kg-1,pH 8.12和7.76。供试秸秆为13C标记玉米秸秆,采用连续标记的方法,在玉米拔节期至大喇叭口期,用Ba13CO3(98 atom%,上海化工研究院)和盐酸反应产生的13CO2连续标记15 d,每天标记时间为9:00—17:00,标记15 d后即收取全株玉米,烘干,粉碎通过2 mm筛,有机碳含量为424.3 g·kg-1,全氮18.7 g·kg-1,δ13C值为855.25‰。
1.3 培养试验
试验设置4个处理,即传统耕作土壤不加秸秆(CT)、免耕土壤不加秸秆(NT)、传统耕作土壤加秸秆(CTS)和免耕土壤加秸秆(NTS),每个处理15次重复。添加秸秆处理均为取5 g标记玉米秸秆与过2 mm筛的风干土样100 g(秸秆占风干土的质量百分数为5%)[21-22]混匀后装入500 mL的玻璃广口瓶中,加入蒸馏水至土壤最大持水量的70%,用中间带透气滤纸的塑料膜封闭瓶口,在25℃恒温培养箱中通气培养,每周称重保持土壤水分。培养周期为180 d,分别于培养后第15、30、60、90和180天取各处理3次重复样品进行指标测定。
1.4 测试方法
土壤样品进行各项目测定前用镊子将土壤中可见秸秆挑出。
土壤水稳性团聚体:称取50 g风干土平铺于2 mm筛子上,室温下用蒸馏水浸润5 min后手动上下振动筛子,幅度为3 cm,振动2 min,共50次。振动完毕后用蒸馏水将筛子上的土样冲洗进铝盒中。按照上述方法将土样依次通过0.25 mm和0.053 mm筛,分别获得>2 000 μm、2 000—250 μm、250—53 μm和<53 μm四部分团聚体。
土壤有机碳测定采用干样燃烧法,所用仪器为元素分析仪(Vario Macro C/N,Germany),测定前土样经1 mol·L-1盐酸处理,去除无机碳。土壤有机碳δ13C值采用稳定同位素质谱仪ISOPRIME100(Isoprime,英国)测定。
土壤微生物生物量碳(MBC)采用氯仿熏蒸-硫酸钾浸提方法测定[17],具体计算方法参照马想等[18]。
β-葡萄糖苷酶(BG)、β-纤维二糖苷酶(CBH)、β-木糖苷酶(BXYL)活性采用荧光微孔板检测技术测定[23],具体操作及计算方法参照宋霄君等[24]。
1.5 计算方法
土壤中秸秆来源碳在SOC中所占比例(F)计算公式为:
F= (δ13Ct-δ13C0) / (δ13Cstraw- δ13C0) (1)
式中,δ13Ct为培养各时期添加秸秆土壤有机碳的δ13C值;δ13C0为培养初始未加秸秆时土壤有机碳δ13C值;δ13Cstraw为添加秸秆的δ13C值。
1.6 统计分析
采用Microsoft Excel 2007和SAS9.2进行数据处理和作图,采用一般线性模型进行方差分析和相关分析,最小显著极差法(LSD法)进行多重比较,显著性水平<0.05。数据为平均值±标准偏差(=3)。
2 结果
2.1 土壤有机碳含量
由图1可知,保护性耕作和添加秸秆显著提高SOC含量。除15 d外,NT处理SOC含量较CT提高32%—38%,达到显著水平。添加秸秆15—180 d,CTS处理SOC含量为23.09—31.73 g·kg-1,较CT提高15%—46%;NTS处理SOC含量为30.11—33.25 g·kg-1,较NT提高12%—21%。CT和CTS处理SOC含量随时间变化趋势一致,即0—15 d显著增加,15—60 d显著下降;不同的是CT处理SOC在60 d 后基本保持不变,而CTS处理SOC显著增加。NT和NTS处理SOC含量随时间变化趋势一致,即0—30 d显著增加,随后下降。培养结束时,CT、NT、CTS、NTS处理SOC含量分别为18.8、25.1、27.4和30.5 g·kg-1,其中CT、NT处理的SOC含量较初始分别降低13.0%、11.9%,CTS、NTS处理的SOC含量较初始分别提高26.8%和7.0%。综上可知,添加秸秆不仅能够显著提高SOC含量,且对传统耕作处理效果更佳。
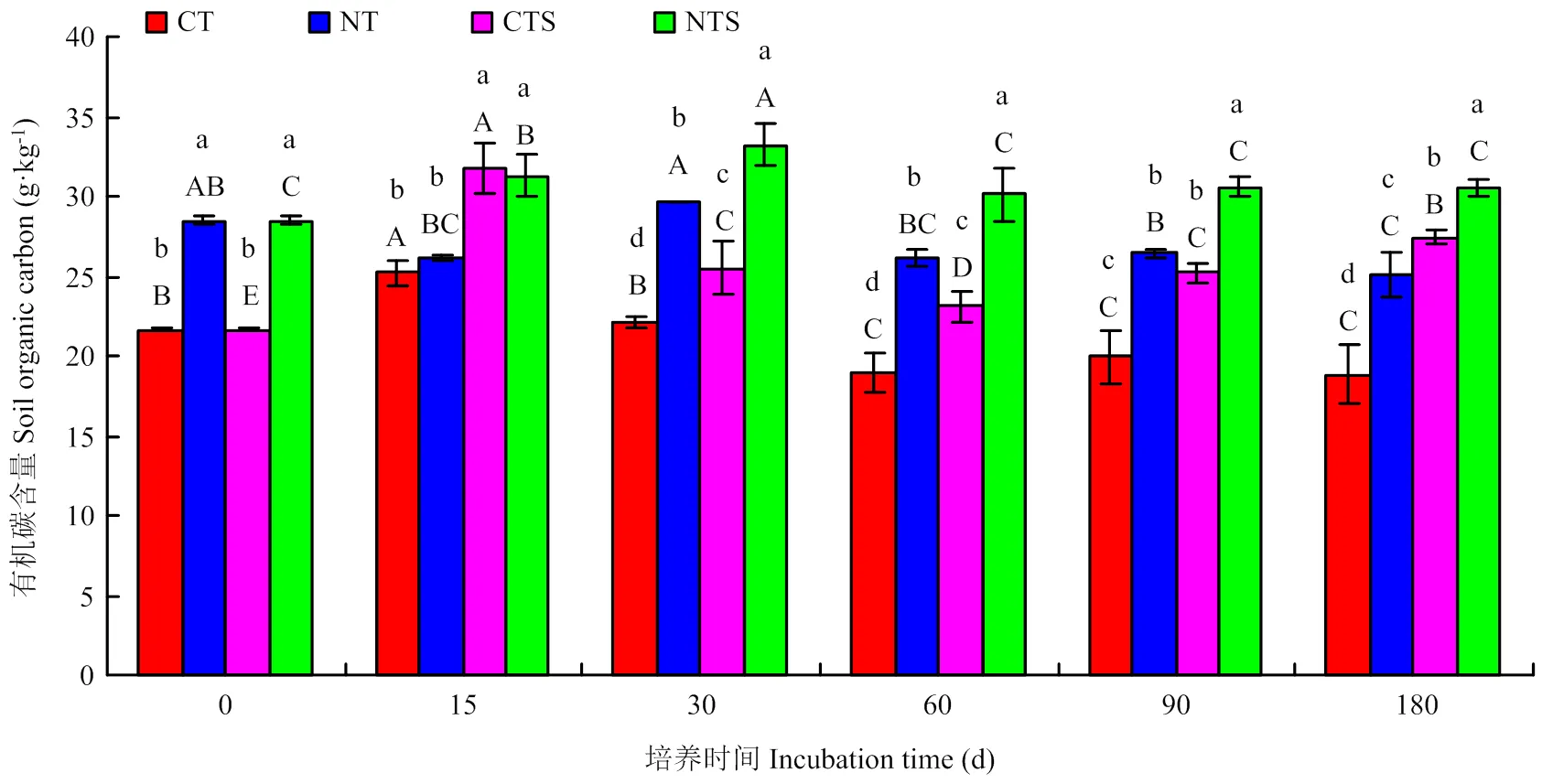
图中小写字母代表处理间差异显著,大写字母代表不同培养时期间差异显著(P<0.05)
2.2 土壤水稳性团聚体构成变化
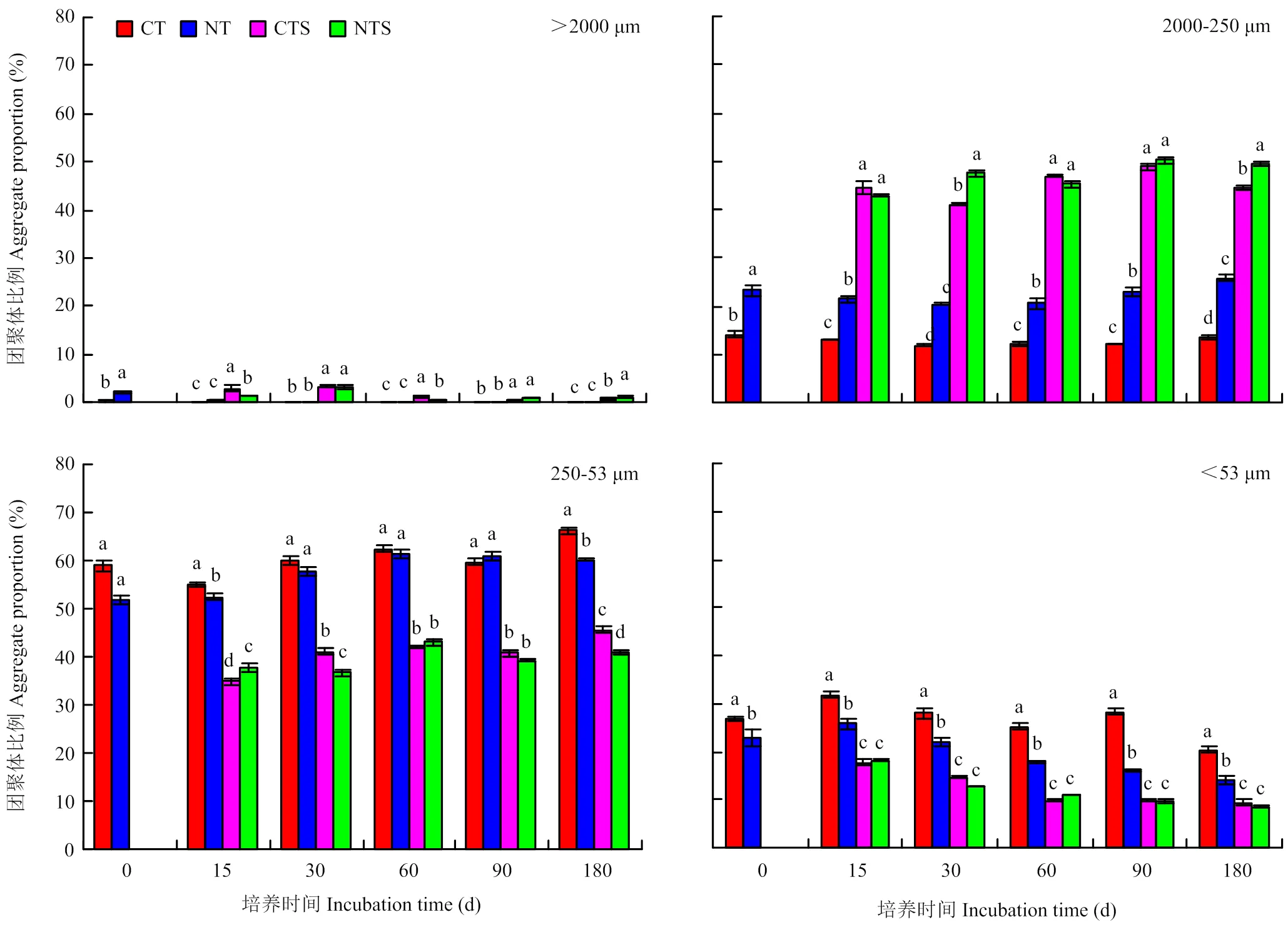
由图2可知,在未加秸秆时,CT和NT处理土壤水稳性团聚体以250—53 μm粒级为主体,占全部团聚体的52%—66%;NT显著提高>250 μm团聚体比例,较CT提高66%—92%。添加秸秆对团聚体构成有较大影响。CTS和NTS处理团聚体以2 000—250 μm粒级最高,占全部团聚体的41%—50%。15 d—180 d中,CTS处理>250 μm团聚体比例为44.4%—49.4%,较CT处理提高235%—310%;250—53 μm和<53 μm团聚体比例分别为34.8%—45.6%和9.4%—17.7%,较CT处理分别降低31%—37%和44%—65%。NTS处理>250 μm团聚体比例为44.1%—50.9%,两部分总比例较NT处理提高96%—149%;250—53 μm和<53 μm团聚体比例分别为36.7%—43.0%和8.6%—18.1%,较NT处理分别降低28%—36%和30%—42%。由此可知,添加秸秆能够显著改变土壤水稳性团聚体构成,并且对传统耕作土壤的影响更大。此外,CTS和NTS各级团聚体含量仅在个别培养时期差异显著,其他培养时间两处理间无显著差异,说明添加秸秆可减小传统耕作与免耕土壤之间团聚体含量的差异。
2.3 土壤有机碳δ13C值及秸秆新碳比例
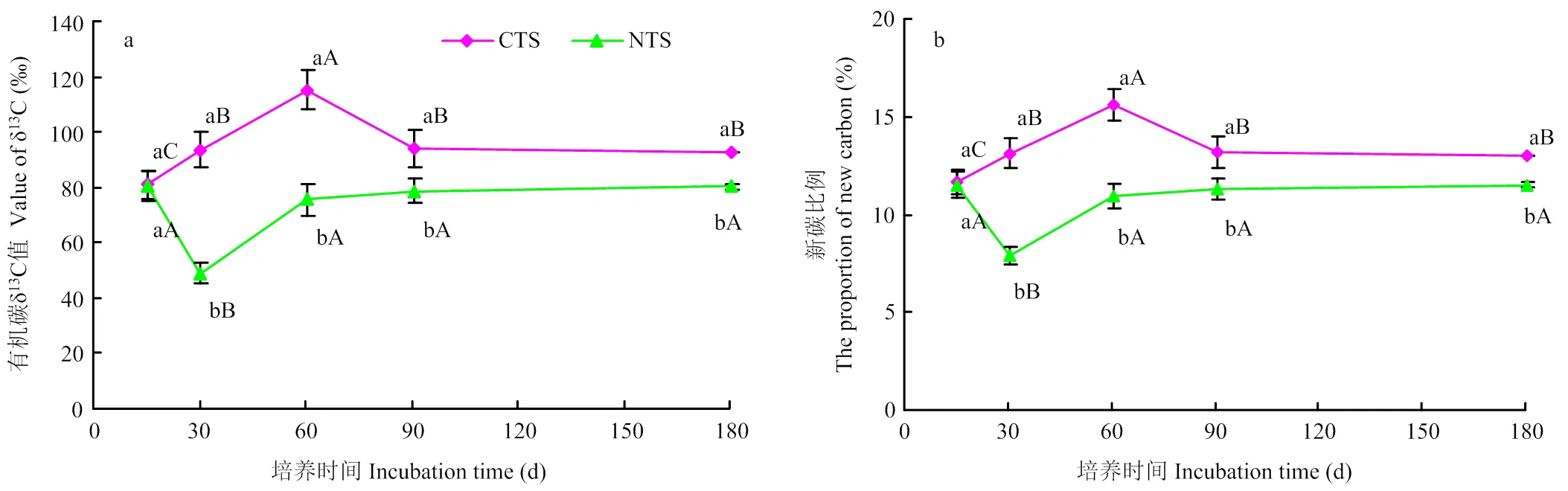
未加秸秆处理CT和NT有机碳δ13C值在整个培养期间变化很小,分别为-21.30‰±0.04‰和-20.43‰± 0.06‰。添加秸秆后土壤有机碳δ13C值增加显著(图3-a)。CTS处理有机碳δ13C值为80.93‰—115.22‰,NTS为48.92‰—80.49‰。CTS有机碳δ13C值在15—60 d持续增加,随后开始减小,至90 d后维持稳定;NTS有机碳δ13C值在15—30 d减小,随后开始增加,至60 d后维持稳定。在整个培养期间,除第15天外,CTS处理有机碳δ13C值均显著高于NTS。综上可知,CTS和NTS处理有机碳δ13C值随时间的变化趋势相反,且CTS增加高于NTS。由图3-b可知,秸秆来源碳占SOC的比例与有机碳δ13C值变化趋势一致。15—180 d(除第15天外),CTS处理秸秆来源碳所占比例均显著高于NTS,为11.7%—15.6%,较NTS提高13%—66%。由此可知,秸秆碳对传统耕作SOC的影响更大。
图中误差线为标准误差,不同小写字母代表不同处理间差异显著(P<0.05)
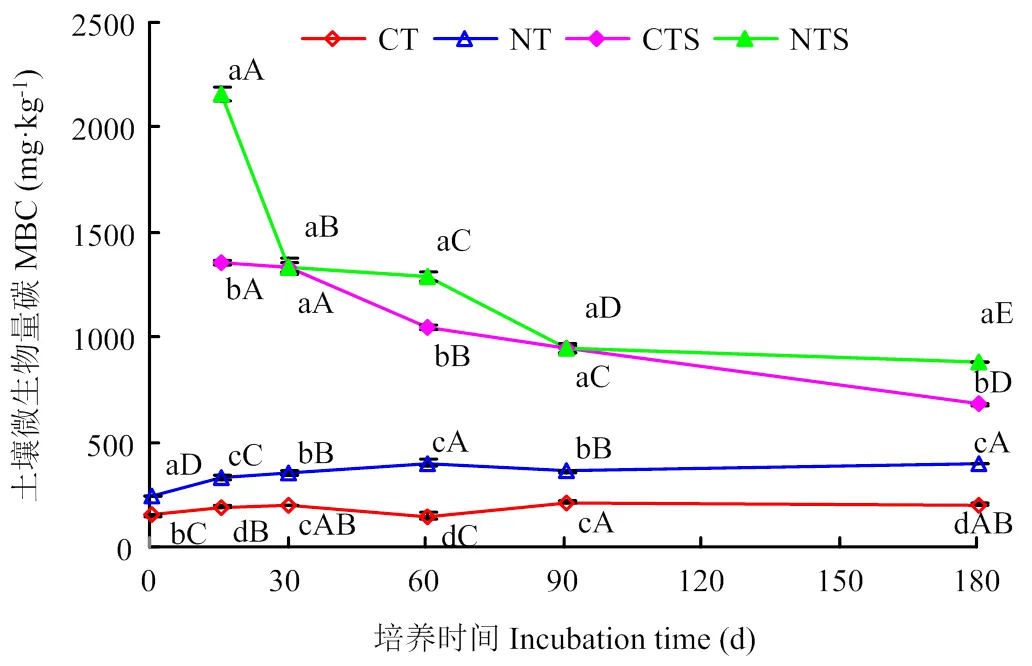
2.4 土壤微生物量碳变化
由图4可知,添加秸秆能显著提高土壤MBC含量,15—180 d期间,各处理MBC含量大小顺序为NTS>CTS>NT>CT。CTS处理MBC含量为679— 1 354 mg·kg-1,较CT提高239%—623%;NTS处理MBC含量为880—2 162 mg·kg-1,较NT提高124%—555%。未加秸秆时,NT较CT提高MBC含量72%—178%;添加秸秆后,NTS与CTS间MBC含量差距减小,较CTS提高幅度为1%—60%。随时间延长,CTS和NTS处理MBC含量均呈下降趋势,CT和NT处理呈先升高后降低的趋势。培养结束时,CT、NT、CTS、NTS处理MBC含量分别为200、392、679和880 mg·kg-1,较初始MBC含量分别提高34%、61%、353%和262%。综上可知,添加秸秆对传统耕作土壤MBC的提高作用更大。
图中小写字母代表处理间差异显著,大写字母代表不同培养时期间差异显著(P<0.05)
2.5 土壤酶活性变化
由图5-a可知,添加秸秆和不加秸秆条件下免耕处理β-葡萄糖苷酶活性均显著高于传统耕作处理,且添加秸秆能显著提高β-葡萄糖苷酶活性。在整个培养期间,NT较CT提高β-葡萄糖苷酶活性38%—211%。CTS处理β-葡萄糖苷酶活性大小为96—433 nmol·g-1·h-1,较CT提高58%—170%;NTS处理为227—527 nmol·g-1·h-1,较NT提高28%—181%。随时间延长,各处理β-葡萄糖苷酶活性表现为升高–降低的变化趋势,最高值均出现在培养第15 d。培养结束时,CT、NT处理β-葡萄糖苷酶活性分别为61、86 nmol·g-1·h-1,较培养初期分别降低31%和59%;CTS和NTS分别为96和242 nmol·g-1·h-1,较培养初期分别提高9%和16%。
图中小写字母代表处理间差异显著,大写字母代表不同培养时期间差异显著(P<0.05)
由图5-b可知,添加秸秆和不加秸秆条件下免耕处理β-纤维二糖苷酶活性显著高于传统耕作处理,且添加秸秆能显著提高β-纤维二糖苷酶活性。整个培养期间,NT处理较CT提高β-纤维二糖苷酶活性52%—337%。CTS处理β-纤维二糖苷酶活性大小为14—105 nmol·g-1·h-1,较CT提高80%—411%;NTS处理为27—158 nmol·g-1·h-1,较NT提高4%—304%。随时间延长,CT和CTS处理β-纤维二糖苷酶活性表现为升高–降低的变化趋势,NT表现为降低趋势,NTS表现为升高–降低–升高的变化趋势。培养结束时,CT、NT、CTS和NTS处理β-纤维二糖苷酶活性分别为6、12、14和46 nmol·g-1·h-1,较培养初始值分别降低64%、78%、8%和18%。
图5-c为β-木糖苷酶活性变化情况,添加秸秆和不加秸秆条件下免耕处理β-木糖苷酶活性均显著高于传统耕作处理。培养期间,NT较CT提高β-木糖苷酶活性31%—320%。CTS处理β-木糖苷酶活性大小为21—64 nmol·g-1·h-1,较CT提高117%—170%;NTS处理为32—121 nmol·g-1·h-1,较NT提高13%—118%。随时间延长,各处理β-木糖苷酶活性变化趋势与β-葡萄糖苷酶活性一致。培养结束时,CT、NT、CTS和NTS处理β-木糖苷酶活性分别为10、21、23和42 nmol·g-1·h-1,较培养初始值分别降低59%、71%、4%和44%。
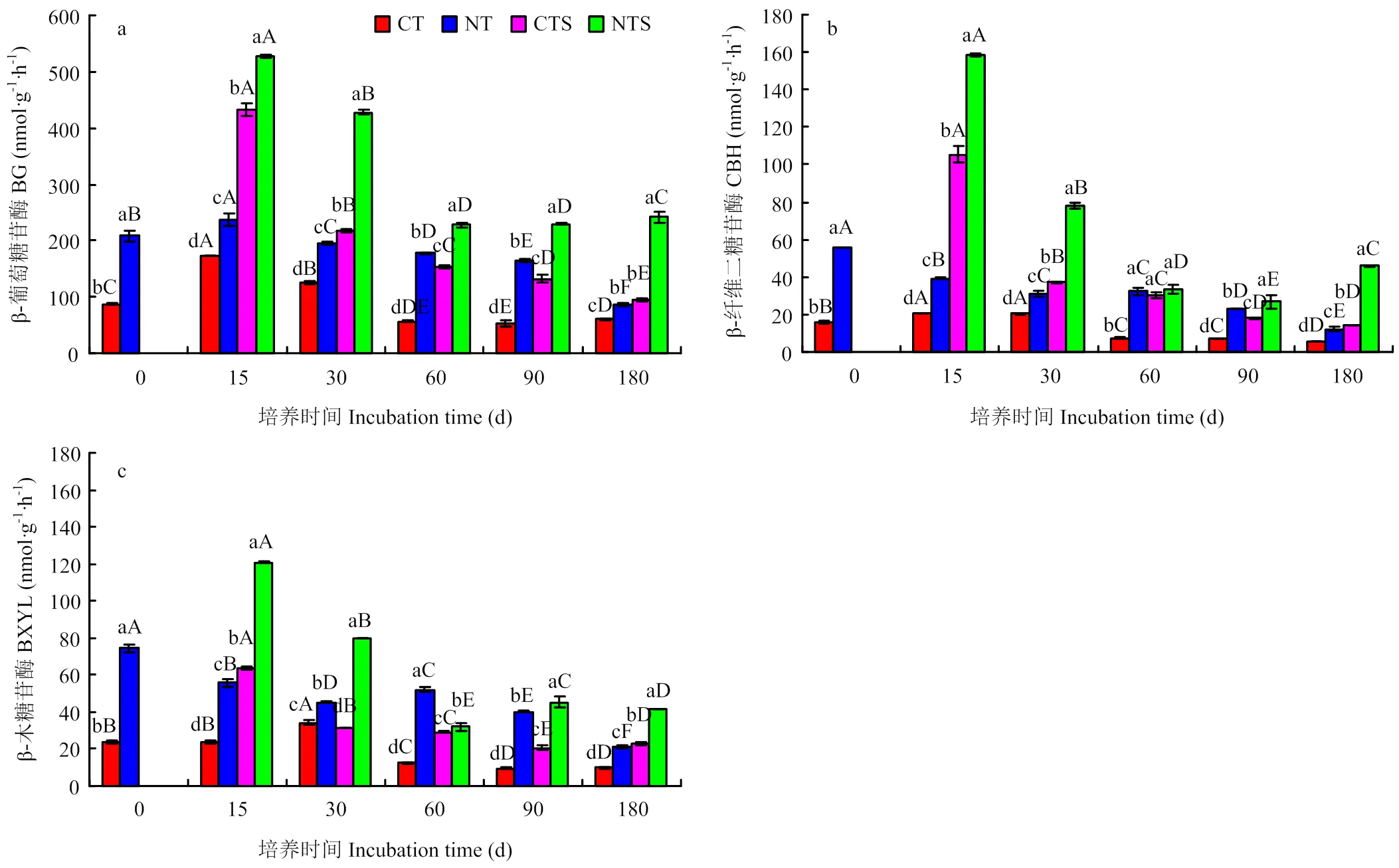
图中小写字母代表处理间差异显著,大写字母代表不同培养时期间差异显著(P<0.05)
2.6 土壤有机碳、土壤酶活性及团聚体组成的相关分析
由表2中结果可知,BG、CBH、BXYL 3种酶活性彼此之间均表现为极显著正相关关系,且均与MBC、>2 000 μm团聚体、2 000—250 μm团聚体(除BXYL外)显著正相关,与250—53 μm团聚体显著负相关。SOC含量与BG、CBH、BXYL 3种酶活性、MBC及>2 000 μm、2 000—250 μm团聚体比例显著正相关,与250—53 μm、<53 μm团聚体比例呈显著负相关关系。通过线性相关分析可知(表3),>2 000 μm、2 000—250 μm团聚体比例可解释SOC变化的25%和48%,MBC可解释SOC变化的45%,BG、CBH和BXYL酶活性,分别可解释SOC变化的66%、44%、53%(表3)。
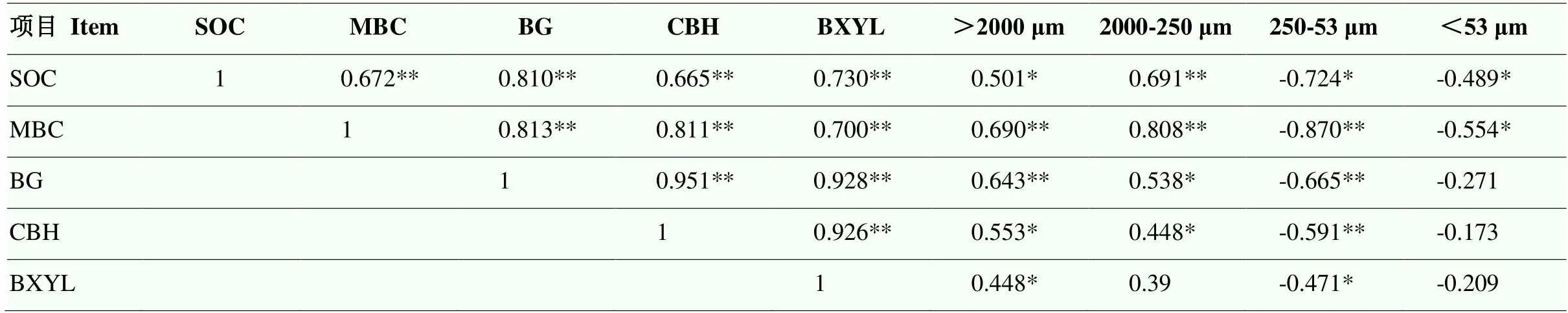
表2 土壤有机碳、土壤酶活性与微生物量碳、团聚体组成的相关系数
表中“*”代表相关性达到显著水平(<0.05),“**”代表相关性达到极显著水平(<0.01),“>2000 μm、2000-250 μm、250-53 μm和<53 μm”表示土壤中对应粒级团聚体所占质量百分比
The symbol “*” indicates significant correlation between items (<0.05) and “**”represents a highly significant correlation between items (<0.01); “>2000 μm、2000-250 μm、250-53 μm and <53 μm” represent the mass proportion of aggregates in soil
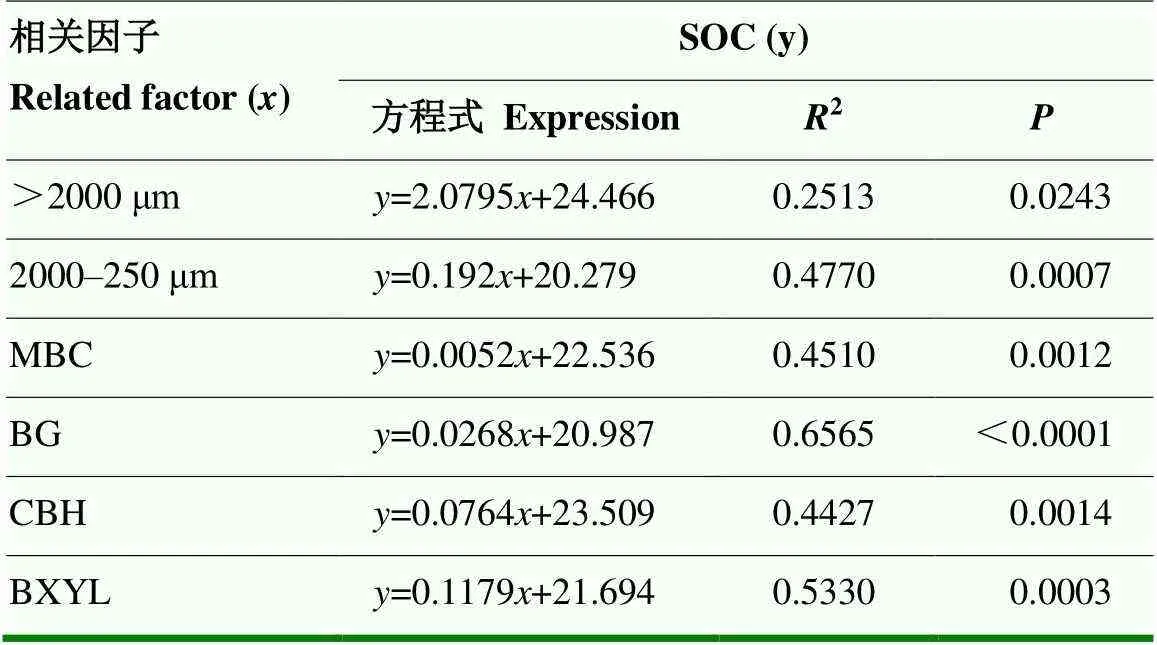
表3 有机碳与各相关因子关系表达式
3 讨论
3.1 不同耕作措施及添加秸秆对土壤有机碳的影响
不同耕作措施由于对土壤的扰动程度不同,因此对SOC含量的影响不同。本研究中,NT处理SOC含量较CT提高32%—38%,达到显著水平。这主要是因为经过长期的保护性耕作,对土壤翻动小,降低了SOC的矿化,同时在田间操作过程中增加了有机物质输入,因此积累了较多的SOC。张霞等[25]的研究结果也表明,连年免耕能够显著增加土壤表层SOC含量。秸秆还田作为改善土壤肥力的重要农田管理措施,被认为是增加土壤碳储量和保持良好结构的重要技术[26]。本研究中添加秸秆显著提高了各耕作SOC含量,CTS、NTS处理的SOC含量分别较CT、NT处理提高15%—46%和12%—21%。徐国鑫等[27]的研究得到类似结果,发现秸秆还田能提高SOC含量15.30%—23.04%。其原因一方面是由于秸秆作为外源有机碳直接补充进SOC含量,另一方面添加秸秆对土壤团聚体构成的改善也发挥着重要作用,本研究中添加秸秆显著提高了大团聚体含量,由12%—37%提升至39%—54%,而团聚体对SOC的固定作用被认为是最重要的机制[27-28]。秸秆进入土壤后可作为胶结物质参与到团聚体的形成过程并成为团聚体的一部分,同时土壤大团聚比例提高能够增强土壤结构稳定性,进而减少SOC矿化[22]。本研究还发现,添加秸秆对SOC的影响在传统耕作土壤中更加显著,这可从Pei等[29]的研究找到解释,其研究结果表明秸秆碳对低肥力SOC的贡献率大于高肥力土壤,主要是由于高肥力土壤中微生物活动高于低肥力土壤,导致秸秆碳分解转化迅速,因此对高肥力SOC作用小[8],由初始养分含量可知,本研究中CTS和NTS处理的土壤正好与低肥力和高肥力水平相对应。
3.2 不同耕作措施及添加秸秆对土壤微生物量碳和酶活性的影响
耕作措施和添加秸秆除了影响土壤物理化学性状外,对土壤生物性质也存在显著影响。本研究结果表明,免耕和添加秸秆能够显著提高土壤微生物量碳含量。这一方面得益于秸秆为土壤微生物提供了充足的能量,促进微生物大量繁殖和生长[30],另一方面耕作和秸秆可通过对土壤团聚体形成的改变进而影响土壤微环境,包括土壤水分孔隙和空气孔隙、土壤孔隙度、温度等,这些对土壤中微生物数量和群体结构都会产生显著影响[31]。HELGASON[32]的研究表明土壤大团聚体对土壤中水分渗透和氧气有效性具有显著影响,这与本研究中微生物量碳与2 000—250 μm团聚体含量呈极显著正相关关系、与250—53 μm、<53 μm团聚体间存在显著负相关关系的结果相一致。本研究还发现添加秸秆对传统耕作土壤微生物量碳的提高作用更大,可能原因为传统耕作对土壤破坏性大、无秸秆还田,因此本身肥力水平较低,微生物数量较少,在这种情况下添加秸秆为土壤微生物提供了充足的养分,使其生长迅速;而免耕土壤由于破坏性小、有秸秆覆盖还田,因此本身土壤肥力较高,能够保证一定数量微生物生长,在此条件下添加秸秆虽然同样提供了充足的养分,但对微生物数量提升作用有限[8]。β-葡萄糖苷酶(BG)、β-纤维二糖苷酶(CBH)和β-木糖苷酶(BXYL)是参与有机碳转化的主要调节者[33],本研究表明添加秸秆能够显著提高β-葡萄糖苷酶、β-纤维二糖苷酶和β-木糖苷酶活性,这一结果与前人研究结果[23,33-34]类似。这主要是因为外源有机物为土壤微生物提供了丰富的能源原料,增强它们的新陈代谢,而土壤酶主要来自微生物和植物根系的分泌作用,因此土壤酶活性得到提升。β-葡萄糖苷酶、β-纤维二糖苷酶和β-木糖苷酶活性与微生物量碳呈显著正相关关系也说明了这一点。相关研究表明玉米秸秆在添加后12—14 d分解率达到最大,此后逐渐下降并趋于稳定[8],与本研究中β-葡萄糖苷酶、β-纤维二糖苷酶和β-木糖苷酶活性在第15天活性最高,然后降低的趋势一致。这主要与微生物代谢利用物质相关,起初阶段其主要分解秸秆中易分解组分,如淀粉、蔗糖、低聚糖等,代谢速度快;后期主要利用纤维素、木质素等难分解物质,微生物代谢速度慢[8]。由于在水解纤维素时需要β-葡萄糖苷酶、β-纤维二糖苷酶协作,木质纤维素中含有的半纤维素需要β-木糖苷酶水解,因此3种酶活性存在极显著正相关关系(<0.01)。同时相关研究表明,在纤维素酶系统中β-葡萄糖苷酶含量应足够多才能保证水解完全并防止水解产物的抑制作用[35],本研究中β-葡萄糖苷酶酶活性显著高于β-纤维二糖苷酶和β-木糖苷酶活性,可以从这里获取解释。
3.3 不同耕作措施下添加秸秆提高有机碳机制分析
添加13C标记秸秆显著提高了土壤有机碳δ13C值,这与大多数研究结果相同,说明外源秸秆有机碳很快进入到土壤中[36]。添加相同秸秆后不同土壤中的δ13C值存在显著差异,即CTS处理>NTS处理,说明相同条件下,进入到土壤中的秸秆数量为CTS>NTS。不同的是NTS处理δ13C值先下降后升高,而CTS先升高后下降,由微生物量碳含量和β-葡萄糖苷酶、β-纤维二糖苷酶、β-木糖苷酶活性可知,NTS处理微生物活动较强,因此能够在短时间内分解矿化掉秸秆碳中易分解部分,造成δ13C值出现下降情况。CTS和NTS处理的初始SOC含量差异显著,正是由于初始条件的差异造成两种土壤固定秸秆来源有机碳的能力不同[37],SOC含量较高处理(NTS)土壤中固定有机碳位点被占用数量高于SOC含量较低处理(CTS),当大量外源有机碳进入土壤时,对其固定量必然较小[20]。这与土壤团聚体构成相关,研究结果显示当土壤中大团聚体(>250 μm)比例较高时,SOC含量随之升高,由相关关系可知,大团聚体可解释SOC变化的48%。秸秆碳在土壤中的分布还受土壤微生物活动的影响。本研究中添加秸秆处理显著增加了各处理土壤中微生物量碳含量,这是由于秸秆刺激了土壤微生物的生长和活性,而秸秆碳在不同处理土壤中分布的差异性除了团聚体的作用外,很大程度上受微生物作用调节,本研究结果显示微生物量碳可解释SOC变化的45%。当土壤经过处理(耕作或添加秸秆),土壤中SOC降解可能由细菌主导途径转换为真菌主导途径[38]。在不同耕作处理土壤中添加秸秆后其周转途径受微生物影响可能不同。ZHENG等[39]研究得出,由于在大团聚体中存在较多的新近有机碳和易分解有机碳,因此会导致大团聚体中细菌和真菌之间的竞争增强;而在微团聚体中存在较多的耐分解有机碳,因此这部分土壤中细菌和真菌间的协同作用会更强以达到分解有机碳获得能量的目的。此外,本研究中添加秸秆显著提高了β-葡萄糖苷酶、β-纤维二糖苷酶和β-木糖苷酶活性,分别可解释SOC变化的66%、44%、53%;但对β-纤维二糖苷酶提高幅度最大,而β-纤维二糖苷酶的主要来源是真菌,因此添加秸秆后SOC周转可能以真菌为主,因为真菌释放的胞外酶范围广,对于木质素、纤维素等难降解有机碳的能力更强[40]。综上可知,秸秆在土壤中的周转不单取决于团聚体的物理保护,还受微生物的数量和构成影响。
4 结论
不同耕作措施下土壤添加秸秆可显著提高土壤有机碳和大团聚体含量,促进微生物数量增加和β-葡萄糖苷酶、β-纤维二糖苷酶和β-木糖苷酶活性增强。土壤有机碳变化与水稳性大团聚体(>250 μm)含量、微生物量碳、上述3种酶活性呈显著正相关关系,与微团聚体(<250 μm)含量呈显著负相关关系。可知,有机碳在土壤中的固定除了受团聚体物理保护外,还受土壤中微生物作用的调节。针对不同耕作土壤,添加秸秆对传统耕作土壤有机碳及其相关因素的影响更大。由此我们建议,从快速增加土壤有机碳角度出发,可根据实际情况采用粉碎翻压方式进行秸秆还田。
[1] 范亚琳, 刘贤赵, 高磊, 汪亚及, 彭新华. 不同培肥措施对红壤坡耕地土壤有机碳流失的影响.土壤学报, 2019, 56(3): 638-649.
FAN Y L, LIU X Z, GAO L, WANG Y J, PENG X H. Effects of fertility-building practices on soil organic carbon loss with sediment in sloping cropland of red soil. Acta Pedologica Sinica, 2019, 56(3): 638-649. (in Chinese)
[2] HAN L, SUN K, JIN J, XING B S. Some concepts of soil organic carbon characteristics and mineral interaction from a review of literature. Soil Biology and Biochemistry, 2016, 94: 107-121.
[3] 苏永中, 赵哈林. 土壤有机碳储量、影响因素及其环境效应的研究进展. 中国沙漠, 2002(3): 19-27.
SU Y Z, ZHAO H L. Advances in researches on soil organic carbon storages, affecting factors and its environmental effects. Journal of Desert Research, 2002(3): 19-27. (in Chinese)
[4] LIANG C H, YIN Y, CHEN Q. Dynamics of soil organic carbon fractions and aggregates in vegetable cropping systems. Pedosphere, 2014, 24(5): 605-612.
[5] 于维水, 李桂花, 王碧胜, 武红亮, 赵雅雯, 孟繁华, 卢昌艾. 不同施肥制度下我国东部典型土壤易分解与耐分解碳的组分特征. 植物营养与肥料学报, 2015, 21(3): 675-683.
YU W S,LI G H,WANG B S, WU H L, ZHAO Y W, MENG F H, LU C A. Component characteristics of soil labile and recalcitrant carbon under long-term different fertilization systems in eastern China.Journal of Plant Nutrition and Fertilizer, 2015, 21(3): 675-683. (in Chinese)
[6] 左旭, 王红彦, 王亚静, 王磊, 景丽, 王道龙. 中国玉米秸秆资源量估算及其自然适宜性评价.中国农业资源与区划, 2015, 36(6): 5-10, 29.
ZUO X, WANG H Y, WANG Y J, WANG L, JING L, WANG D L. Estimation and suitability evaluation of corn straw resources in China. Chinese Journal of Agricultural Resources and Regional Planning, 2015, 36(6): 5-10, 29. (in Chinese)
[7] LIANG G P, WU H J, HOUSSOU A A, CAI D X, WU X P, GAO L L, WANG B S, LI S P. Soil respiration, glomalin content, and enzymatic activity response to straw application in a wheat-maize rotation system. Journal of Soils and Sediments, 2018, 18(3): 697-707.
[8] 杨艳华, 苏瑶, 何振超, 喻曼, 陈喜靖, 沈阿林. 还田秸秆碳在土壤中的转化分配及对土壤有机碳库影响的研究进展.应用生态学报, 2019, 30(2): 668-676.
YANG Y H, SU Y, HE Z C, YU M, CHEN X J, SHEN A L. Transformation and distribution of straw-derived carbon in soil and their effect on soil organic carbon pool: A review . Chinese Journal of Applied Ecology, 2019, 30(2): 668-676. (in Chinese)
[9] ZHENG L, WU W L, WEI Y P, HU K L. Effects of straw return and regional factors on spatio-temporal variability of soil organic matter in a high-yielding area of northern China. Soil & Tillage Research, 2015, 145: 78-86.
[10] 王碧胜, 蔡典雄, 武雪萍, 李景, 梁国鹏, 于维水, 王相玲, 杨毅宇, 王小彬. 长期保护性耕作对土壤有机碳和玉米产量及水分利用的影响. 植物营养与肥料学报, 2015, 21(6): 1455-1464.
WANG B S, CAI D X, WU X P, LI J, LIANG G P, YU W S, WANG X L, YANG Y Y, WANG X B. Effects of long-term conservation tillage on soil organic carbon,maize yield and water utilization. Journal of Plant Nutrition and Fertilizers, 2015, 21(6): 1455-1464.(in Chinese)
[11] PAUSTIAN K, LEHMANN J, OGLE S, REAY D, ROBERTSON G P, SMITH P. Climate-smart soils. Nature, 2016, 532(7597): 49.
[12] ZHANG W J, WANG X J, XU M G, HUANG S M, LIU H, PENG C. Soil organic carbon dynamics under long-term fertilizations in arable land of northern China. Biogeosciences, 2010, 7(2): 409-425.
[13] SIX J, CONANT R T, PAUL E A. Stabilization mechanisms of soil organic matter: implications for C-saturation of soils. Plant and Soil, 2002, 241(2): 155-176.
[14] XIE J, PENG B, WANG R. Responses of crop productivity and physical protection of organic carbon by macroaggregates to long‐term fertilization of an Anthrosol. European Journal of Soil Science, 2018, 69(3): 555-567.
[15] TISDALL J M, OADES J M. Organic matter and water‐stable aggregates in soils. Journal of Soil Science, 1982, 33(2): 141-163.
[16] 刘哲, 孙增慧, 张瑞庆.秸秆添加对潮土团聚体及有机碳分布和稳定性的影响. 西南农业学报, 2018, 31(6): 1246-1252.
LIU Z, SUN Z H, ZHANG R Q. Effects of application of rice straw on distribution and stability of aggregates and organic carbon in fluvo-aquic. Southwest China Journal of Agricultural Sciences, 2018, 31(6): 1246-1252. (in Chinese)
[17] VANCE E D, BROOKES P C, JENKINSON D S. An extraction method for measuring soil microbial biomass C. Soil Biology & Biochemistry, 1987, 19(6): 703-707.
[18] 马想, 黄晶, 赵惠丽, 徐明岗, 姜慧敏, 段英华. 秸秆与氮肥不同配比对红壤微生物量碳氮的影响.植物营养与肥料学报, 2018, 24(6): 1574-1580.
MA X, HUANG J, ZHAO H L, XU M G, JIANG H M, DUAN Y H. Straw and nitrogen fertilizer ratios influence microbial biomass carbon and nitrogen in red soil. Journal of Plant Nutrition and Fertilizers, 2018, 24(6): 1574-1580. (in Chinese)
[19] LI Z Q, LI D D, MA L, YU Y Y, ZHAO B Z, ZHANG J B. Effects of straw management and nitrogen application rate on soil organic matter fractions and microbial properties in North China Plain. Journal of Soils and Sediments, 2019, 19(2): 618-628.
[20] 孙元宏, 高雪莹, 赵兴敏, 隋标, 王鸿斌, 赵兰坡. 添加玉米秸秆对白浆土重组有机碳及团聚体组成的影响. 土壤学报, 2017, 54(4): 1009-1017.
SUN Y H, GAO X Y, ZHAO X M, SUI B, WANG H B, ZHAO L P. Effects of corn stalk incorporation on organic carbon of heavy fraction and composition of soil aggregates in albic soil. Acta Pedologica Sinica, 2017, 54(4): 1009-1017. (in Chinese)
[21] 仇建飞, 窦森, 邵晨, 李明敏, 安丰华. 添加玉米秸秆培养对土壤团聚体胡敏酸数量和结构特征的影响. 土壤学报, 2011, 48(4): 781-787.
QIU J F, DOU S, SHAO C, LI M M, AN F H. Effects of corn stalk application on quantity and structural characteristics of humus acid in soil aggregates. Acta Pedologica Sinica, 2011, 48(4): 781-787. (in Chinese)
[22] 王碧胜, 于维水, 武雪萍, 高丽丽, 李景, 李生平, 宋霄君, 刘彩彩, 李倩, 梁国鹏, 蔡典雄, 张继宗. 添加玉米秸秆对旱作土壤团聚体及其有机碳含量的影响.中国农业科学, 2019, 52(9): 1553-1563.
WANG B S, YU W S, WU X P, GAO L L, LI J, LI S P, SONG X J, LIU C C, LI Q, LIANG G P, CAI D X, ZHANG J Z. Effect of straw addition on the formation of aggregates and accumulation of organic carbon in dryland soil. Scientia Agricultura Sinica, 2019, 52(9): 1553-1563. (in Chinese)
[23] 梁国鹏, Houssou A A, 吴会军, 武雪萍, 蔡典雄, 高丽丽, 李景, 王碧胜, 李生平. 施氮量对夏玉米根际和非根际土壤酶活性及氮含量的影响.应用生态学报, 2016, 27(6): 1917-1924.
LIANG G P, HOUISSOU A A, WU H J, WU X P, CAI D X, GAO L L, LI J, WANG B S, LI S P. Soil nitrogen content and enzyme activities in rhizosphere and non-rhizosphere of summer maize under different nitrogen application rates. Chinese Journal of Applied Ecology, 2016, 27(6): 1917-1924. (in Chinese)
[24] 宋霄君, 吴会军, 武雪萍, 李倩, 王碧胜, 李生平, 梁国鹏, 李景, 刘彩彩, 张孟妮. 长期保护性耕作可提高表层土壤碳氮含量和根际土壤酶活性.植物营养与肥料学报, 2018, 24(6): 1588-1597.
SONG X J, WU H J, WU X P, LI Q, WANG B S, LI S P, LIANG G P, LI J, LIU C C, ZHANG M N. Long-term conservation tillage improves surface soil carbon and nitrogen content and rhizosphere soil enzyme activities. Journal of Plant Nutrition and Fertilizers, 2018, 24(6): 1588-1597. (in Chinese)
[25] 张霞, 张育林, 刘丹, 杜昊辉, 李军, 王旭东. 种植方式和耕作措施对土壤结构与水分利用效率的影响. 农业机械学报, 2019, 50(3): 250-261.
ZHANG X, ZHANG Y L, LIU D, DU H H, LI J, WANG X D. Effects of planting methods and tillage systems on soil structure and water use efficiency. Transactions of the Chinese Society of Agricultural Machinery, 2019, 50(3): 250-261. (in Chinese)
[26] 王秀娟, 解占军, 董环, 赵颖, 刘慧屿, 娄春荣. 秸秆还田对玉米产量和土壤团聚体组成及有机碳分布的影响. 玉米科学, 2018(1): 108-115.
WANG X J, XIE Z J, DONG H, ZHAO Y, LIU H Y, LOU C R. Effects of straw returning on yield and soil aggregates composition and organic carbon distribution. Journal of Maize Sciences, 2018(1): 108-115. (in Chinese)
[27] 徐国鑫, 王子芳, 高明, 田冬, 黄容, 刘江, 黎嘉成. 秸秆与生物炭还田对土壤团聚体及固碳特征的影响. 环境科学, 2018, 39(1): 355-362.
XU G X, WANG Z F, GAO M, TIAN D, HUANG R, LIU J, LI J C. Effects of straw and biochar return in soil on soil aggregate and carbon sequestration. Environmental Science, 2018, 39(1): 355-362. (in Chinese)
[28] 李睿, 江长胜, 郝庆菊.缙云山不同土地利用方式下土壤团聚体中活性有机碳分布特征. 环境科学, 2015, 36(9): 3429-3437.
LI R, JIANG C S, HAO Q J. Impact of land utilization pattern on distributing characters of labile organic carbon in soil aggregates in Jinyun Mountain.Environmental Science, 2015, 36(9): 3429-3437. (in Chinese)
[29] PEI J B, LI H, LI S Y, AN T T, FARMER J, FU S F, WANG J K. Dynamics of maize carbon contribution to soil organic carbon in association with soil type and fertility level. PLoS One, 2015, 10(3): e0120825.
[30] ZHANG X F, XIN X L, ZHU A N, ZHANG J B, YANG W L. Effects of tillage and residue managements on organic C accumulation and soil aggregation in a sandy loam soil of the North China Plain. Catena, 2017, 156: 176-183.
[31] KUNTZ M, BERNER A, GATTINGER A, SCHOLBERG J M, MADER P, PFIFFNER L. Influence of reduced tillage on earthworm and microbial communities under organic arable farming. Pedobiologia, 2013, 56(4/6): 251-260.
[32] HELGASON B L, WALLEY F L, GERMIDA J J. No-till soil management increases microbial biomass and alters community profiles in soil aggregates. Applied Soil Ecology, 2010, 46(3): 390-397.
[33] 郑凤君, 王雪, 李景, 王碧胜, 宋霄君, 张孟妮, 武雪萍, 刘爽, 席吉龙, 张建诚, 李永山. 免耕条件下施用有机肥对冬小麦土壤酶及活性有机碳的影响. 中国农业科学, 2020, 53(6): 1202-1213.
ZHENG F J, WANG X, LI J, WANG B S, SONG X J, ZHANG M N, WU X P, LIU S, XI J L, ZHANG J C, LI Y S. Effect of no-tillage with manure on soil enzyme activities and soil active organic carbon. Scientia Agricultura Sinica, 2020, 53(6): 1202-1213. (in Chinese)
[34] 张鑫, 周卫, 艾超, 黄绍敏, 梁国庆. 秸秆还田下氮肥运筹对夏玉米不同时期土壤酶活性及细菌群落结构的影响. 植物营养与肥料学报, 2020, 26(2): 295-306.
ZHANG X, ZHOU W, AI C, HUANG S M, LIANG G Q. Effects of nitrogen management on soil enzyme activities and bacterial community structure in summer maize growing stages under straw incorporation. Journal of Plant Nutrition and Fertilizers, 2020, 26(2): 295-306. (in Chinese)
[35] MERINO S T, CHERRY J. Progress and challenges in enzyme development for biomass utilization. Advances in Biochemical Engineering/Biotechnology, 2007, 108: 95-120.
[36] 刘哲, 韩霁昌, 孙增慧,余正洪, 张卫华, 高红贝. 外源新碳对红壤团聚体及有机碳分布和稳定性的影响.环境科学学报, 2017, 37(6):2351-2359.
LIU Z, HAN J C, SUN Z H, YU Z H, ZHANG W H, GAO H B. Effects of fresh carbon on distribution and stability of aggregates and organic carbon in red soil. Acta Scientiae Circumstantiae, 2017, 37(6): 2351-2359. (in Chinese)
[37] 吕元春, 薛丽佳, 尹云锋, 高人, 马红亮, 杨玉盛. 外源新碳在不同类型土壤团聚体中的分配规律. 土壤学报, 2013, 50(3): 534-539.
Lü Y C, XUE L J, YIN Y F, GAO R, MA H L, YANG Y S. Distribution of fresh carbon in aggregate fractions of different soil types. Acta Pedologica Sinica, 2013, 50(3): 534-539. (in Chinese)
[38] GRITTITHS B S, DANIELL T J, DONN S, NEILSON R. Bioindication potential of using molecular characterisation of the nematode community: Response to soil tillage. European Journal of Soil Biology, 2012, 49: 92-97.
[39] ZHENG W, ZHAO Z Y, GONG Q L, ZHAI B N, Li Z Y. Responses of fungal–bacterial community and network to organic inputs vary among different spatial habitats in soil. Soil Biology and Biochemistry, 2018, 125: 54-63.
[40] MOORE-KUCERA J, DICK R P. Application of13C-labeled litter and root materials for in situ decomposition studies using phospholipid fatty acids. Soil Biology and Biochemistry, 2008, 40(10): 2485-2493.
Effects of Straw Addition on Soil Organic Carbon and Related Factors Under Different Tillage Practices
WANG BiSheng1,2, YU WeiShui2, WU XuePing2, GAO LiLi3, LI Jing4, SONG XiaoJun2, LI ShengPing2, LU JinJing2, ZHENG FengJun2, CAI DianXiong2
1College of Agronomy, Qingdao Agricultural University, Qingdao 266109, Shandong;2Institute of Agricultural Resources and Regional Planning, Chinese Academy of Agricultural Sciences, Beijing 100081;3Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences, Beijing 100081;4College of Water Resources and Environment, Hebei GEO University, Shijiazhuang 050031
【】Straw addition is an effective means to increase soil organic carbon, which is significant to ensure the sustainability of the organic carbon in the farmland system. This study aimed at investigating the effect of straw addition on soil organic carbon (SOC) and related factors under different tillage treatments, so as to provide a theoretical basis for the management of carbon sequestration and yield increase in northern dry farmland.【】The field soil samples from long-term conventional tillage (CT) and no-tillage (NT) for in-lab incubation experiment were collected. Four treatments were set up, namely, conventional tillage soil without straw (CT), no tillage soil without straw (NT), conventional tillage soil with straw (CTS), and no-till soil with straw (NTS), respectively. Each treatment owned 15 repetitions. The incubation experiment was conducted in a constant temperature incubator at 25 ℃ for 180 days, and the soil samples were periodically taken to determine the content of SOC, aggregate composition, microbial biomass carbon and the activity of related enzymes.【】(1) Straw addition significantly increased the content of soil organic carbon and large aggregates. Compared with CT, CTS increased SOC by 15%-46%; compared with NT, NTS increased SOC 12%-21%; compared to the initial organic carbon content, at the end of cultivation, CTS and NTS increased by 26.8% and 7.0%, respectively. CTS and NTS had the highest particle size of 2 000-250 μm, accounting for 41%-50% of all aggregates. Compared with CT, CTS increased the proportion of aggregates >250 μm by 235%-310%, and NTS increased the proportion of aggregates >250 μm by 96%-149%. (2) The addition of straw significantly increased the δ13C value of soil organic carbon. The CTS treatment was 80.93‰-115.22‰, NTS was 48.92‰-80.49‰; CTS straw-derived carbon was significantly higher than NTS by 13%-66%. (3) The addition of straw significantly increased the microbial biomass carbon (MBC) content, β-glucosidase (BG), β-cellobiosidase (CBH) and β-xylosidase (BXYL) activities. Compared with CT, CTS increased MBC content by 239%–623%, and increased BG, CBH and BXYL activity by 58%-170%, 52%-337% and 117%-170%, respectively; compared to NT, NTS increased MBC content by 124%-555%, and increased BG, CBH and BXYL activities by 28%-181%, 4%-304% and 13%-118%. (4) Soil organic carbon was significantly positively correlated with BG, CBH, BXYL activity, MBC and the proportion of >2 000 μm, 2 000-250 μm aggregates, and negatively correlated with the proportion of 250-53 μm and <53 μm aggregates. The activities of BG, CBH and BXYL showed a very significant positive correlation with each other, and were significantly positively correlated with MBC, >2 000 μm aggregates, 2 000-250 μm aggregates, and extremely negative with <53 μm aggregates. Linear correlation analysis results showed that water-stable macroaggregates (>250 μm) could explain 48% of organic carbon changes, MBC could explain 45% of organic carbon changes, and BG, CBH and BXYL enzyme activities could explain 66%, 44% and 53% of organic carbon changes, respectively.【】The addition of straw could significantly increase the content of soil organic carbon and macroaggregates, increase the number of microorganisms, and promote the soil enzyme activity. The impact on soil organic carbon and its related factors was greater in conventional tillage soils. In addition to the physical protection of aggregates, the sequestration of straw carbon in soil also depended on the role of microorganisms in the soil.
aggregate; soil organic carbon; δ13C; soil enzyme; straw addition; tillage practices

10.3864/j.issn.0578-1752.2021.06.009
2020-05-31;
2020-07-14
国家重点研发计划(2018YFD0200408、2016YFD0300804)、国家科技支撑计划课题(2015BAD22B03)、青岛农业大学博士基金(663/1120069)
王碧胜,E-mail:wangbisheng2@126.com。通信作者武雪萍,E-mail:wuxueping@caas.cn。通信作者蔡典雄,E-mail:caidianxiong@caas.cn
(责任编辑 李云霞)
