miR-27a和miR-605基因的多态性与严重生精障碍的相关性研究
2021-03-18孙文文柯红利阿周存
孙文文,柯红利,阿周存
(1.大理大学基础医学院,云南大理 671000;2.大理大学农学与生物科学学院,云南大理 671003)
生精障碍是男性不育最常见病因,在其发病过程中,遗传因素起着重要作用〔1〕。目前,已发现一些生精障碍的遗传病因,但这些已知的遗传病只能解释一小部分病例,仍有大量的相关基因有待进一步阐明〔2〕。作为一类基因表达的重要调控因子,MicroRNA(miRNA)通过与靶mRNA 的3′-UTRs碱基配对形成双链,介导靶mRNA 裂解或抑制mRNA 翻译形成蛋白质,从而在转录后水平的基因表达调控中发挥关键作用〔3〕。研究表明,miRNA 参与精子发生过程中许多基因转录后水平的表达调控,其异常可能导致生精障碍〔4-6〕。因此,miRNA 基因也被认为是一类新的生精障碍候选基因,研究鉴定生精障碍相关的miRNA 基因,对生精障碍遗传病因的阐明具有重要意义。
miR-27a和miR-605基因是近些年来发现的与哺乳动物和人类精子发生密切相关的miRNA 基因,也被认为是生精障碍和男性不育的重要候选基因〔7-9〕。目前,尚未有miR-27a和miR-605基因的多态性与严重生精障碍的研究报道。本研究在正常男性和严重生精障碍患者中,对miR-27a和miR-605基因内的常见单核苷酸多态性(single nucleotide polymorphism,SNP)位点rs895819 和rs2043556 的多态性分布进行分析和比较,从基因变异的角度,初步探索miR-27a和miR-605基因与严重生精障碍的关系。
1 材料与方法
1.1 研究对象病例组由318例严重生精障碍患者组成,包括严重少精症(精子密度小于5×106个∕mL)患者和无精症患者,病例来自于四川大学华西医院和大理大学第一附属医院。对照组包括234例精子密度正常的男性。所有研究对象均签署知情同意书。
1.2 试剂和仪器DNA提取试剂盒(TIANGEN公司,批号:DP304);dNTPs(TaKaRa公司,批号:AJ30388A);PCR引物(上海生工生物工程股份有限公司);Taq DNA聚合酶(TaKaRa 公司,批号:DR001AM);限制性内切酶DraⅢ和HinfI(Thermo Fisher 生物公司,批号:ER1231、ER0801);PCR 扩增仪(Eppendorf 公司,型号:AG 22331 Hamburg);凝胶电泳仪(Bio-RAD 公司,型号:041BR);凝胶成像分析系统(Bio-RAD 公司,型号:Gel Doc100)。
1.3 方法
1.3.1 PCR 扩增 用DNA 提取试剂盒(TIANamp Genomic DNA Kit)从200 μL 外周血中提取基因组DNA。miR-27a和miR-605的PCR 扩增引物序列、扩增片段长度、退火温度见表1。PCR 扩增反应的体积为25 μL,反应条件为:94 ℃预变性5 min,35个循环(94 ℃变性30 s,57~58 ℃退火30 s,72 ℃延伸40 s),最后72 ℃延伸5 min。
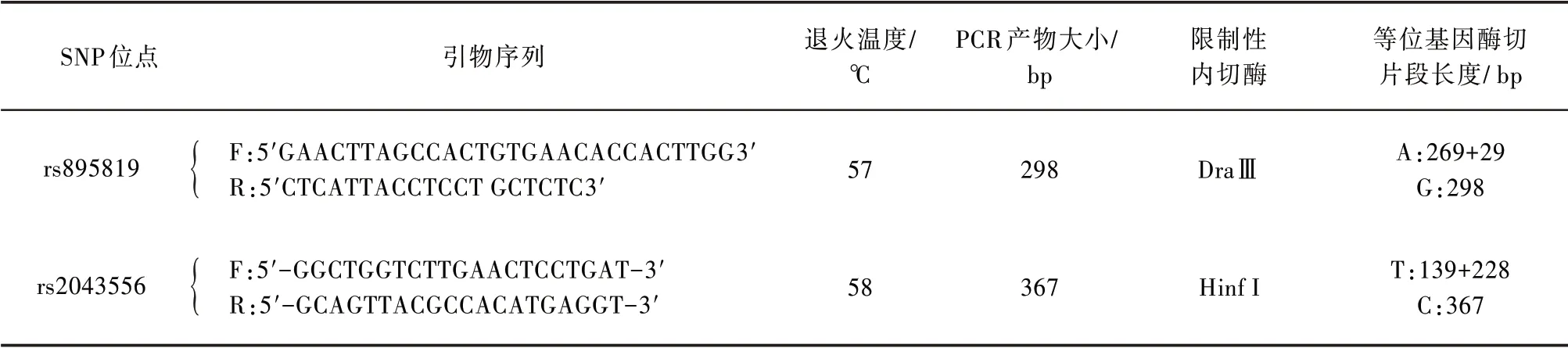
表1 PCR扩增的引物序列和扩增产物大小,RFLP分析所用的限制性内切酶和等位基因的酶切片段长度
1.3.2 基因分型 利用限制性片段长度多态性(restriction fragment length polymorphism,RFLP)分析方法对SNP rs895819和rs2043556进行基因分型。取PCR 产物10 μL,用10 个单位的限制性内切酶消化过夜,然后在2%(rs895819)和3%(rs2043556)的琼脂糖凝胶上进行电泳。所用的限制性内切酶、各等位基因的酶切片段见表1。通过对部分不同基因型的部分标本进行PCR 产物直接测序,对电泳分型结果进行验证。
1.3.3 统计学分析 计数计算基因频率和基因型频率。用HWE 计算器进行Hardy-Weinberg 平衡检验。采用χ2检验比较病例组和对照组之间等位基因和基因型频率之间的差异,检验水准设为P<0.05。
2 结果
miR-27a基因的SNP rs895819 等位基因频率和基因型频率分布在严重生精障碍患者和正常男性之间差异无统计学意义(P>0.05);miR-605基因的SNP rs2043556 等位基因频率和基因型频率分布在严重生精障碍患者和正常男性间的差异具有统计学意义,严重生精障碍患者中rs2043556 的等位基因C 的频率(36.2%vs.29.1%,P=0.013,OR=1.383,95%CI为1.070~1.788)和带有等位基因C 的个体(TC∕CC)的频率(57.2%vs.48.3%,P=0.037,OR=1.433,95%CI为1.021~2.012)显著高于正常男性,而TT 基因型的频率(42.8%vs.51.7%,P=0.037,OR=0.698,95%CI为0.497~0.980)则显著低于正常男性。见表2。
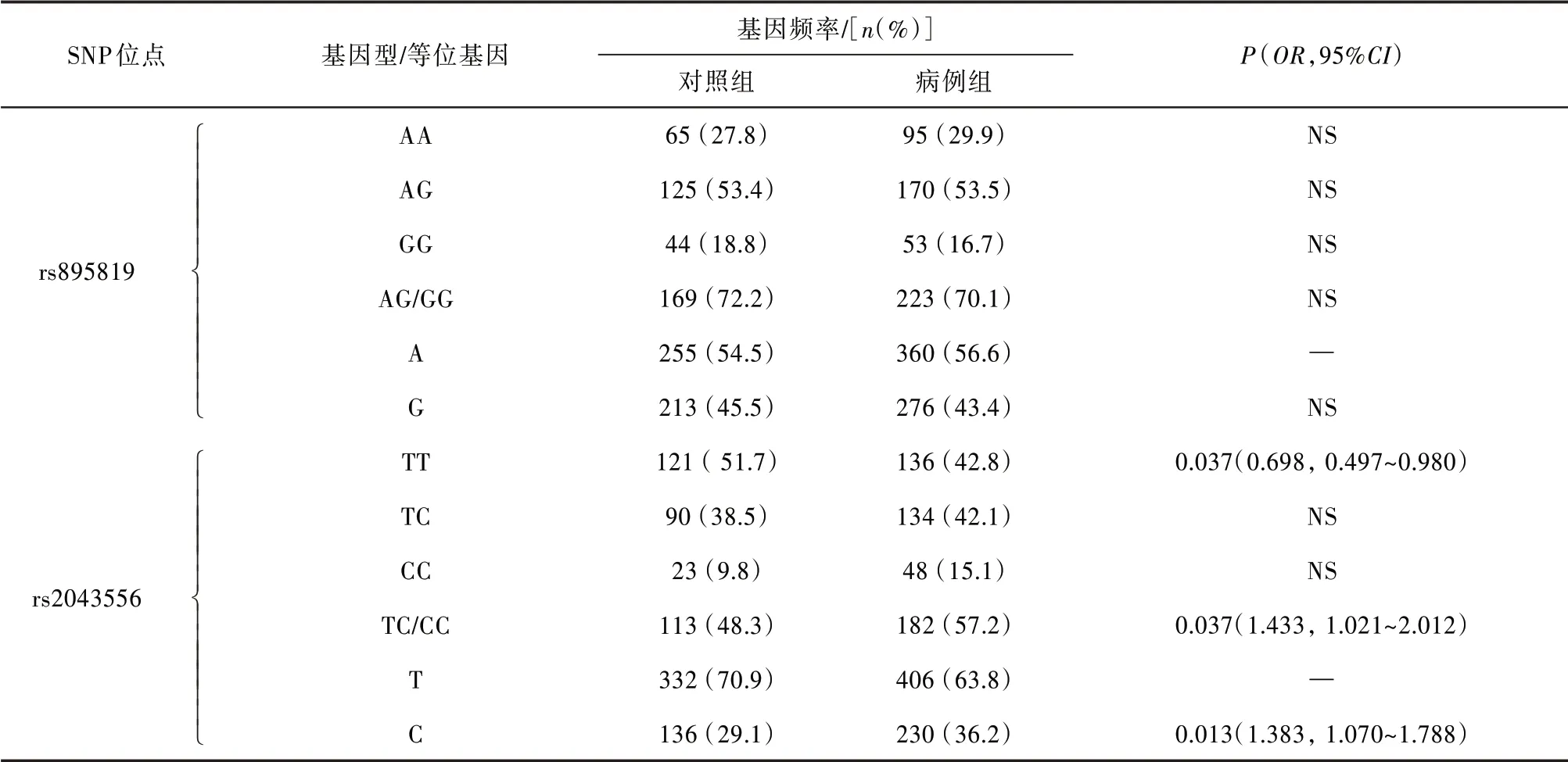
表2 对照组和病例组SNP rs895819 和rs2043556的等位基因频率和基因型频率分布
Hardy-Weinberg 平衡检验结果显示,SNP rs895819 和rs2043556 的基因型分布在严重生精障碍患者和正常男性中符合Hardy-Weinberg 平衡,提示本研究的研究对象具有群体代表性。
SNP rs895819 和rs2043556 的电泳分型结果与测序结果一致,表明本研究RFLP 分型的结果准确、可靠。见图1。
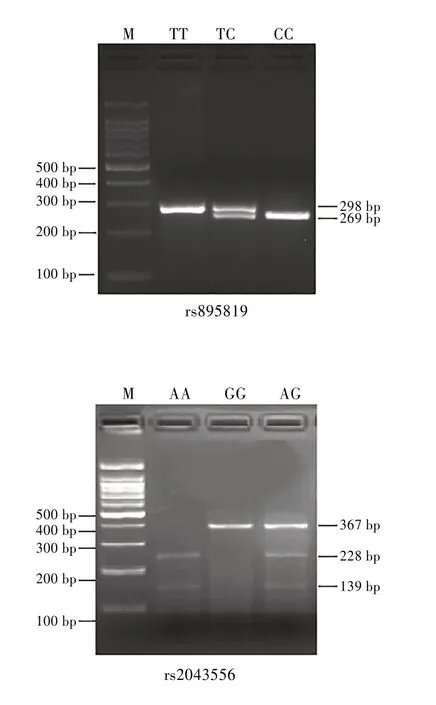
图1 SNP rs895819和rs2043556的电泳分型结果
3 讨论
本研究对生精障碍的候选基因miR-27a和miR-605的常见SNP rs895819 和rs2043556 的多态性与严重生精障碍的相关性进行了调查。调查结果显示,miR-27a基因的SNP rs895819 的等位基因频率和基因型频率分布在严重生精障碍患者和正常男性间差异无统计学意义,提示SNP rs895819 的多态性与严重生精障碍不相关。然而,在miR-605基因的SNP rs2043556位点上,严重生精障碍患者的等位基因C 频率和带有等位基因C 的个体(TC∕CC)的频率显著高于正常男性,基因型TT的频率则显著低于正常男性。这些结果表明,SNP rs2043556的多态性与严重生精障碍相关联,基因型TT是严重生精障碍的保护性基因型(降低发病风险),而等位基因C可能增加严重生精障碍的发病风险。
上述结果显示,SNP rs2043556 影响严重生精障碍的发病风险,但具体原因尚未清楚。SNP rs2043556 是一个位于pre-miR-605结构区域的C∕T变异。该变异影响转录后pre-miR-605的加工效率,进而影响成熟miR-605的表达水平。与等位基因T 相比,等位基因C 的miR-605表达水平显著降低〔10〕。有研究显示,miR-605能在mRNA 水平和蛋白水平抑制靶基因MDM2表达,其表达水平降低将导致MDM2过表达〔10-12〕。MDM2基因是一个在生精过程中发挥重要作用的基因,其过表达可能会对正常的生精功能造成较大的影响,导致生精障碍,如无精症和严重少精症〔13-14〕。因此,SNP rs2043556通过影响miR-605的表达水平,进而引起miR-605一些在生精过程中有重要作用的靶基因的表达异常,最终影响正常的生精功能,导致生精障碍。这可能是SNP rs2043556 影响严重生精障碍发病风险的原因之一。
本研究首次对miR-27a和miR-605基因的常见SNP rs895819 和rs2043556 多态性与严重生精障碍的关系进行了调查,发现miR-605的SNP rs2043556的多态性与严重生精障碍相关,其变异影响严重生精障碍的发病风险,提示miR-605可能参与生精障碍的病理过程。研究结果有可能为生精障碍的遗传病因学提供有价值的参考。
