天然产物抑制黄曲霉生长及其产毒的研究进展
2021-03-18朱晓嫚谢岩黎
李 倩,赵 颖,朱晓嫚,谢岩黎
河南工业大学 粮油食品学院,粮油食品安全检测与控制重点实验室,河南 郑州 450001
黄曲霉(AF)属于真菌界子囊菌门盘菌亚门发菌科曲霉菌属[1],易污染储藏农作物及加工食品。受污染的农作物较为广泛,如粮食作物(玉米、水稻、小麦等)和油料作物(花生、大豆、棉籽等)等[2-3]。多数黄曲霉代谢产生聚酮类产物——真菌毒素(AFT),在目前已知的20余种黄曲霉毒素中,按照毒性从强到弱依次为AFB1、AFM1、AFG1、AFB2、AFM2、AFG2,AFB1毒性最强,约为氰化钾的10倍、砒霜的68倍、标准致癌物二甲基亚硝胺的75倍、三聚氰胺的416倍[4-5]。除此之外,黄曲霉还会引起人和动物的曲霉病,如霉菌性角膜炎[6];降低人体免疫力,诱发原发性胃癌、肺癌[7-8];还能以肝脏为靶器官,与慢性肝炎病毒协同致癌[9]。鉴于黄曲霉及其毒素的巨大危害,如何抑制黄曲霉的生长和毒素的合成有着重要的研究意义。
近年来,许多天然产物安全、环保、来源广泛,展现了良好的抑菌效果,其抑菌活性被广泛研究。如绿叶挥发物(GLVs)抑制储藏玉米中黄曲霉孢子的萌发[10]、香菇纤维素衍生物GPX抑制黄曲霉毒素的产生[11]、肉桂精油抑制莲子中黄曲霉菌丝的生长及毒素的产生[12]。部分天然产物的最小抑菌浓度(MIC)和最小杀菌浓度(MFC)如表1所示。作者根据化学式结构,将天然产物分为醛类、黄酮类、烯类、醇类、多酚类、醌类、肽类和其他类化合物,重点阐述不同类别的天然产物对黄曲霉生长及毒素合成的抑制机理。
1 醛类化合物
醛类是广泛存在于植物精油中的挥发性小分子化合物,对柠檬醛和肉桂醛的研究较多。关于植物精油中醛类的抑菌机理,近年来的报道均证明是多途径复合作用的结果,主要体现在以下几个方面:(1)通过破坏细胞壁、细胞膜的结构,抑制黄曲霉孢子萌发及菌丝生长,如2-羟基-4-甲氧基苯甲醛(HMB)[27]、邻香草醛(未发表)诱导细胞内容物渗出破坏细胞膜通透性,其中HMB还能够破坏菌丝隔膜,破坏细胞壁完整性。(2)损伤线粒体功能,造成能量代谢以及胞内酶系统紊乱,如Ma等[10]发现青叶醛降低乙酰辅酶A、ATP和线粒体脱氢酶的活性。相似的结论在罗曼等[28-29]的试验中已得到验证。(3)多途径下诱导黄曲霉细胞凋亡。Tian等[30]报道紫苏醛通过诱导活性氧(ROS)、钙离子及细胞凋亡蛋白酶增加引发孢子凋亡。
2 黄酮类化合物
黄酮类化合物广泛存在于芸香科、唇形科、豆科、伞性科、银杏科与菊科等植物中,具有抗菌消炎、抗肿瘤、抗病毒等生物活性功能[31]。李钟美[32]研究发现,高良姜中主要的抑菌活性成分是黄酮醇类物质,主要为高良姜素、槲皮素、山萘黄酮醇等。其中,高良姜素抑制黄曲霉菌丝生长,其抗菌特性可能与其B环上羟基的缺乏有关[33],可强烈阻止肝脏内AFB1向黄曲霉毒素B1-8,9-环氧化物的转化,而后者是黄曲霉毒素的最终致癌物[34]。槲皮素通过下调孢子发育相关基因(AFLA-083110和AFLA-044790)的表达抑制黄曲霉孢子萌发[35]。同时,槲皮素还具有抗氧化能力,能够激活抗氧化系统转录因子Yap1,极大提高抗氧化酶活性,并通过下调产毒调控基因AflR、AflS的表达抑制黄曲霉毒素的合成[15]。此外,也有研究表明,槲皮素能通过竞争性结合黄曲霉毒素合成途径的关键酶——聚酮化合物合成酶的活性位点发挥抑制作用[36]。另外,黄酮中的代表化合物白杨素,对许多致病菌有显著的抑制作用,如意大利青霉[37]和金黄色葡萄球菌[38]等。Yang等[37]对此类黄酮的构效关系研究表明,不同碳位置上的羟基可能是抗真菌效果的潜在影响因素,如C4位置上的羟基会增强白杨素对意大利青霉的抑菌活性,C3位置上的羟基则相反,C2和C3之间不饱和键的氢化会降低白杨素对意大利青霉的抑菌活性;糖基化结合位点的不同和糖类成分的差异对抑菌活性有不同的影响。此外,有相关文献表明具有抑菌作用的黄酮中有一半以上为二氢黄酮类化合物[39],如乔松素[40]、二氢杨梅素等。熊大胜等[41-42]的试验表明二氢杨梅素对黄曲霉、金黄色葡萄球菌等均有抑制作用,但也有研究表明,二氢杨梅素对霉菌抑制作用并不明显[43],这可能与菌株或是药敏试验中选用浓度不同有关。本课题组前期研究表明,二氢杨梅素能够显著抑制黄曲霉孢子萌发,其最小抑菌浓度为4 000 μg/mL(未发表),抑菌机理正在研究中。然而,目前对二氢黄酮类化合物的抑菌机制研究尚有待深入,羟基对其构效关系的影响也有待试验验证。

表1 天然产物抑制黄曲霉的最小抑菌浓度和最小杀菌浓度
3 烯类化合物
植物中具有抑菌活性的烯类化合物有柠檬烯、α-蒎烯、香叶烯、金合欢烯、松油烯、石竹烯、桧烯等。梁海燕等[44]发现,柠檬烯对多种真菌,如黄曲霉、寄生曲霉、金黄色葡萄球菌都具有抑制作用,是一种广谱抑菌剂,与α-蒎烯[45-46]、罗勒烯相似,其抑菌作用靶点均为细胞膜,除此之外,柠檬烯还能够抑制黄曲霉毒素的产生[47]。另有报道,某些烯类化合物能够降低AFB1所致小鼠肝癌的发生率,如银杏叶提取物中的萜烯类化合物下调肝组织肝癌相关基因MXR7的表达、提高肝细胞抑制因子P16蛋白的表达[48]。但此试验只是降低了肝癌的发生率,并没有完全防止肝癌的发生,其抑制机理还有待研究。
4 醇类化合物
醇类物质也是广泛存在于植物中的重要活性成分,其中如香叶醇、橙花醇等已被《食品安全国家标准 食品添加剂使用标准 GB 29938—2013》列入允许使用的食品天然香料名单。梁海燕等[44]研究发现芳樟醇通过抑制线粒体呼吸及破坏细胞膜抑制黄曲霉生长。香叶醇则主要通过诱导细胞内ROS积累发挥抑制作用[49]。橙花醇抑制黄曲霉的作用机理还与钙离子超载有关,二者的积累引起线粒体功能障碍,增加氧化应激反应,从而激活细胞色素c的释放和提高半胱天冬酶活性[22]。这3种醇的化学结构式见图1,它们互为同分异构体,其中香叶醇与橙花醇因甲基和氢原子顺反结构的不同互为立体异构,芳樟醇中有一个双键与香叶醇和橙花醇有位置上的差别,因此是位置异构体[50]。橙花醇、芳樟醇、香叶醇抑制黄曲霉的MIC分别为0.5、0.6、0.5 μL/mL[25-26],它们之间的差异可能和结构上的差别相关。除此之外,具有抑制黄曲霉生成及毒素合成的醇类化合物还有香茅醇、香芹醇等[51]。
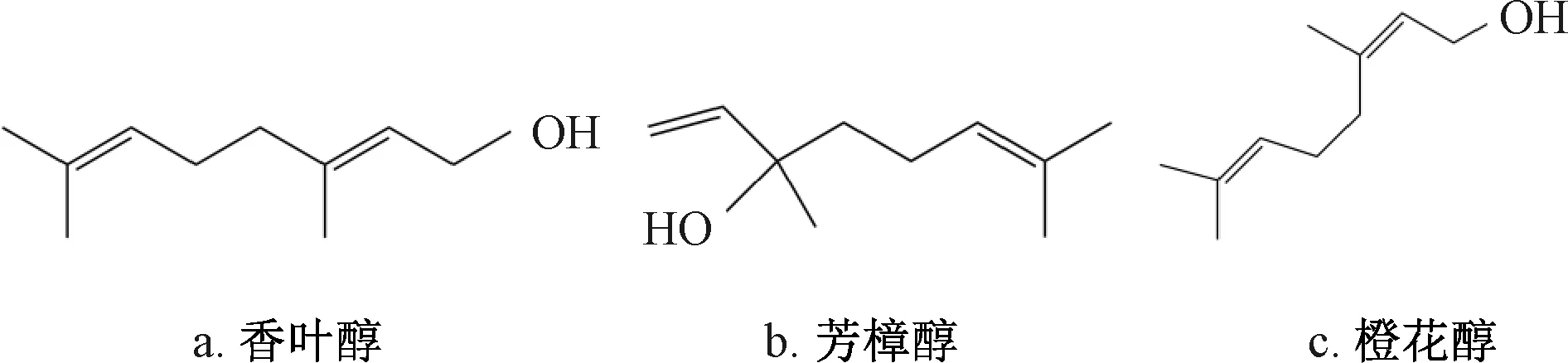
图1 香叶醇、芳樟醇和橙花醇的化学结构式
5 多酚类化合物
多酚类化合物中研究较多的有茶多酚、姜黄素等。茶多酚是广泛存在于茶叶中的一种天然抗氧化剂,也是普洱茶中的主要抑菌活性成分[52]。茶多酚不仅抑制黄曲霉的生长,而且对黄曲霉毒素合成有显著抑制效果[53]。茶多酚可严重破坏黄曲霉菌丝体的超微结构:(1)破坏细胞壁,瓦解黄曲霉菌的固有形态;(2)破坏细胞膜,使胞内外物质交换受阻,抑制细胞膜上的酶正常发挥作用,降低细胞对营养物质的吸收,阻碍生物大分子合成。姜黄素是来自香料姜黄的天然多酚化合物,Ferreira等[54]发现,姜黄素可抑制黄曲霉毒素的产生,机理可能涉及细胞氧化应激和过氧化。此外,姜黄素还是一种有效的光敏剂,经可见光在60 J/cm2、420 nm照射处理后抑菌活性显著增强[55]。百里香酚是一种高效的抑杀黄曲霉孢子的酚类化合物,凋亡机制涉及以下两个方面:(1)增加ROS和NO含量[56];(2)特异靶向细胞膜上的电压门控钾离子通道β亚甲基KCNAB[57]。
6 醌类化合物
醌类化合物从结构上可分为苯醌、萘醌、菲醌和蒽醌,其中萘醌是植物和微生物中活性最高的天然代谢产物,具有抗菌、抗肿瘤等活性。凤仙花醌、胡桃醌、β-拉帕醌均属于萘醌,抑制酵母菌机理涉及破坏线粒体功能[58]。2-甲氧基-1,4-萘醌(MeONQ),可显著抑制意大利青霉的生长,作用靶点包括细胞壁、细胞膜、线粒体氧化还原酶等[59],但对黄曲霉是否有抑制作用还有待研究。其他抑制黄曲霉的醌类物质还有芦荟大黄素、兰雪醌、紫草素、百里香醌等[60]。然而许多醌类化合物,如凤仙花醌、胡桃醌对哺乳动物细胞有毒性[58],这种毒性机理包括:(1)催化NADPH和O2引发的氧化还原循环,使O2转化为有害的ROS;(2)醌环上的未取代碳原子结合亲核生物分子。因此,如果将醌类化合物用于食品防霉制剂,需要经过严格的药物安全性评价。
7 肽类化合物
从植物和微生物中分离到的抗菌肽是拮抗黄曲霉的重要组分。如短小芽孢杆菌HY1分泌的抗真菌肽[61]、黄曲霉拮抗菌B10-6-1分泌的脂肽[62]等能够显著抑制黄曲霉孢子萌发和菌丝生长。在韩玉竹等[25]的试验中,辣椒籽抗菌肽显著降低黄曲霉摄取营养物质的能力,这很可能与辣椒籽抗菌肽破坏了细胞膜的通透性有关。这与黄现青[63]报道的环状脂肽通过抑制营养物质的吸收抑制点状青霉的观点一致。从链霉菌、玫瑰茄、海洋海绵和海洋细菌中分离出来的环二肽——L-亮氨酰-L-脯氨酰,在3.5 mg/mL时可抑制黄曲霉毒素相关基因aflR、hexB、pksL1和dmtA的转录,但对黄曲霉生长影响很小,只有当质量浓度大于6 mg/mL时,才能够抑制其生长。此二肽具有3个异构体,不同浓度下,它们抑制代谢产物——降散盘衣酸的积累、孢子形成的程度也有所不同,抑制作用可能与某些酶基因的下调相关[64]。这是关于环二肽抑制黄曲霉毒素合成的首次报道,其详细机理及具体应用还有待探讨。
8 其他类化合物
抑制黄曲霉生长及其毒素合成的化合物还有生物碱[4]、哌啶化合物[65]、多糖[11, 66]等。吲哚属于生物碱的一种,MIC为100 μg/mL,一定范围内,抑菌效果随吲哚浓度升高而增强,低浓度的吲哚会促进黄曲霉毒素的产生,只有当吲哚浓度接近或者大于MIC时,才可有效抑制AFT,此抑制机理与吲哚对产毒基因aflR的调控有关[26]。 Caceres等[67]发现胡椒碱是一种哌啶化合物,它通过调节细胞的氧化应激反应抑制AFT合成,并从基因层面上对此发现进行了更深入的探究:(1)在胡椒碱的作用下,参与AFB1生物合成的27个基因中有25个基因被显著下调,其中9个基因的表达被轻度抑制(2~3倍),如aflD、aflQ、aflC等,13个基因的表达呈现出4~8倍的抑制,其中以aflN、aflW、aflK下调最为明显,而aflR、aflT则无显著变化;(2)参与编码氧化应激的20个基因中16个被显著调节,其中包括转录因子(msnA、srrA)、全调因子VeA、双加氧酶ppoc和细胞受体gprp在内的5个下调基因,氧化应激反应因子(sskA、ap1、atfA、atfB、sod1、cat、cat2)、氧化脂质生物合成因子(ppa、ppoB)、GPR1和GPRH这11个上调因子,其中aflB、sod1上调效果最为显著;(3)过氧化物酶和超氧化物歧化酶过表达。李红波等[66]对几种多糖进行研究发现,大多数的β-葡聚糖抑制黄曲霉生长及产毒,部分α-葡聚糖,如马铃薯直链淀粉也具有这种抑制功能,但也有一些多糖的作用相反,如A型大豆多糖能促进黄曲霉生长和产毒。由于葡聚糖分子量较大,不易进入真菌细胞内,推测其细胞膜可能存在多糖受体介导抑菌机制[66]。香菇纤维素衍生物GPX也是一种β-葡聚糖,GPX能够促进黄曲霉细胞内囊泡过早与大液泡融合[11]。由于AFT合成最后几步在囊泡中完成[68],然后囊泡经过运输和胞吐释放AFT,因此,囊泡过早融合成大液泡会使AFT的合成、转运、释放受阻[11]。综上所述,多糖也是一种潜在的可以抑制黄曲霉的天然产物。
9 展望
在对黄曲霉及其毒素的污染防控技术中,传统的化学法处理简单、成本低,但污染严重,可能会破坏产品营养成分。物理法操作简单、方便高效,但仪器成本高。微生物法因其特异性强、污染小、能够保证食品营养安全等优点成为最具发展前景的方法[69],然而微生物法目前尚处于实验室阶段。利用天然产物抑制黄曲霉生长及其毒素合成是防控中的研究热点。
近年来,多数抑菌研究已经从简单的生理生化指标检测转向靶标探索,如细胞壁、细胞膜、线粒体成为了多种抑菌剂的共同靶标。除此之外,宏观的整体的研究已经转向亚细胞乃至分子层面的探索,如某些参与黄曲霉生长和毒素合成的基因/蛋白的表达。这些研究都为开发靶向抑菌剂提供了重要的理论基础。鉴于黄曲霉对多种抑菌剂,包括天然的和合成的化合物均衍生出了一定的耐受机制[24],因此,如何改造、修饰天然产物以及如何将有效活性成分合理复配,以提升抑菌剂的稳定性及长期的抑菌效果仍然是目前防霉控霉方向的重要问题,是延长粮食储藏期和食品货架期、提高我国粮食出口安全以及食品品质的重要发展方向。
