长期施氮对暖温带油松林土壤呼吸及其组分的影响*
2021-03-13汪金松赵秀海
杨 璐 汪金松 赵 博 赵秀海
(1.北京林业大学国家林业和草原局森林经营工程技术研究中心 北京 100083; 2.中国科学院地理科学与资源研究所生态系统网络观察与模拟重点实验室 北京 100101)
全球化肥使用增加及化石燃料燃烧,使氮排放速率由1860年的0.15亿 t N·a-1升至1995年的1.56亿 t N·a-1,预计到2050年全球氮沉降速率将达到2.7亿 t N·a-1(Gallowayetal., 2008)。氮沉降速率增加将对陆地生态系统碳循环产生显著影响(Pregitzeretal.,2008; Zhaoetal., 2018),并通过改变碳循环而影响未来气候的变化轨迹(Crowtheretal., 2019)。有研究表明,在氮限制型生态系统中,氮添加会显著降低生态系统土壤碳排放(Janssensetal., 2010; Yanetal., 2018),增加土壤碳储量并促进植物生长(Sunetal., 2014; 刘修元等, 2015),然而过量的氮输入也会导致土壤酸化(Tianetal., 2015; Lietal., 2018)、破坏土壤养分平衡(Xiaetal., 2017)等。
土壤呼吸是陆地生态系统与大气之间二氧化碳交换的第二大通量,其通量是化石燃料燃烧排放二氧化碳的10倍(Bond-Lambertyetal., 2010),在陆地碳循环中起着非常重要的作用(Savageetal., 2013)。森林生态系统作为陆地表层最大的碳库(Dixonetal., 1994),其土壤呼吸通量占整个森林生态系统呼吸通量的70%以上(Littonetal., 2007),因此土壤呼吸的微小变化都将对森林生态系统产生很大影响。土壤呼吸主要受土壤水热因子的协同影响(Laganièreetal., 2012; 汪金松等, 2013; Chenetal., 2014)。另外,土壤呼吸的温度敏感性指数(Q10)被认为是陆地生态系统中反馈碳循环的一个关键参数(Curiel Yusteetal., 2004)。土壤自养呼吸和异养呼吸的Q10分别代表不同的生态学过程,由于植物根系和土壤微生物对环境变化响应的敏感性不一致,导致自养呼吸和异养呼吸的Q10对环境变化响应的差异(Yuetal., 2017)。
虽然国内外已开展了诸多关于氮沉降影响森林土壤呼吸的研究,但由于试验时间、环境条件及生态系统类型的不同,氮沉降对森林土壤呼吸的影响至今没有统一结论。主要包括氮沉降对土壤呼吸促进(Tuetal., 2013; Lietal., 2019)、无显著影响(Allisonetal., 2008; Samuelsonetal., 2009)和显著降低(Yanetal., 2018; Zhaoetal., 2018)这3类结果。其中一个内在原因在于氮沉降对土壤呼吸不同组分(自养和异养呼吸)影响的大小和方向不一致,进而导致土壤呼吸的氮沉降响应差异(Zhouetal., 2014)。全球整合分析表明,氮沉降通过降低微生物生物量和改变微生物群落结构(Ramirezetal., 2012; 刘彩霞等, 2015; Zhengetal., 2018)使温带森林土壤异养呼吸速率显著降低17%(Janssensetal., 2010)。氮沉降降低异养呼吸速率的机制主要包括:氮沉降通过直接改变土壤含氮量进而抑制与有机质分解有关的酶活性(Chenetal., 2017; 刘星等, 2015),进而阻碍凋落物分解和养分释放(Sinsabaughetal., 2005),使异养呼吸速率降低(Zhaoetal., 2018);氮沉降引起土壤酸化而降低微生物生物量和改变微生物群落多样性(Keeleretal., 2009)进而抑制异养呼吸。而自养呼吸主要受光合产物供应的影响(Nadelhoffer, 2000; Lietal., 2018);同时在氮限制的森林生态系统中,氮沉降主要通过影响根系生长进而影响土壤自养呼吸(Sunetal.,2014)。Bowden等(2004)在北美温带阔叶林研究发现,施氮后第一年显著增加了土壤呼吸速率,这是由于施氮后根系生物量明显增加引起自养呼吸速率增加导致的;第二年施氮未改变土壤呼吸速率;在施氮的第13年,土壤呼吸速率显著降低了41%,这主要是由于长期施氮降低了土壤微生物活性进而抑制了异养呼吸,但未改变自养呼吸。前人研究表明,施氮对自养呼吸和异养呼吸的影响可能会随时间发生变化。因此,了解土壤呼吸及其组分对长期施氮的响应对理解森林生态系统土壤碳的动态和稳定性具有重要意义。
目前,土壤呼吸及其组分的温度敏感性对氮添加的响应还存在很大不确定性,进而限制了人们理解陆地生态系统与气候间的反馈关系。研究表明,氮沉降可通过影响植物生长和土壤微环境等进而改变自养呼吸和异养呼吸的Q10值(Zhaoetal., 2018; Zhengetal., 2018)。短期氮添加可通过减少光合产物向根际的分配进而使自养呼吸Q10值降低(Savageetal., 2013);氮沉降可抑制土壤中酚氧化酶活性(Maetal., 2013; 张艺等, 2017)进而降低异养呼吸的Q10值。另外施氮可抑制与有机质分解有关的酶活性并形成难降解的有机物,进而增加有机碳稳定性,亦会降低异养呼吸的Q10值(Chenetal., 2018)。Sun等(2014)在中国河北温带森林的氮添加试验研究表明,4年氮添加降低了自养呼吸和异养呼吸的Q10值,然而在同一研究样地,7年氮添加未显著改变自养速率和异养呼吸的Q10值(Yanetal., 2018);这表明自养呼吸和异养呼吸的Q10值可能会随施氮时间而变。尽管如此,关于长期氮添加影响自养速率和异养呼吸Q10值的研究还鲜见报道(Yanetal., 2018)。
油松(Pinustabulaeformis)是我国北方重要的用材树种和针叶树种之一。山西太岳山林区作为“油松之乡”,是我国油松天然林生态系统保存最完善的地区之一,油松天然林碳储量占整个太岳山林区的25%(伊锋等, 2017)。本研究以山西太岳山油松天然林为对象,于2009年开始进行了为期10年的施氮试验。前期研究发现,施氮后第3~7年(2011—2015)土壤呼吸速率未显著改变(Zhaoetal., 2018),但施氮第8~10年(2016—2018)的土壤呼吸及其组分如何响应仍需进一步揭示。鉴于此,本研究主要探讨生长季土壤呼吸及其组分对长期(8~10年)施氮的响应,拟回答以下2个科学问题:1)长期施氮条件下土壤呼吸及其组分对不同氮水平的响应差异;2)土壤呼吸及其组分的主要调控因素。
1 研究区概况
研究区位于山西省太岳山森林生态系统定位研究站(112.02°—112.40°E,36.52°—36.72°N),林区海拔1 150~2 088 m。该地气候属典型的温带半干旱大陆性季风气候,年均气温8.6 ℃,最高气温主要在7月份并可达22.4 ℃,最低气温集中在1月份并低至-4.6 ℃;年均降水量662 mm,降水集中在夏季;平均相对空气湿度65%。土壤为棕壤和褐土(杨璐等,2018)。自20世纪90年代初,由于严格禁伐,本研究区未受到任何人为干扰,其中90%以上的乔木都是油松,主要灌木有沙棘(Hippophaerhamnoides)、黄刺玫(Rosaxanthina)、虎榛子(Ostryopsisdavidiana)和胡枝子(Lespedezabicolor)等。
2 研究方法
2.1 试验设计
2009年7月,在太岳山森林生态系统定位研究站辖区内,选取有代表性的油松天然林,建立12块20 m×20 m固定样地,各样地间隔10 m以上。本地自然氮沉降量为21.2 kg N·hm-2a-1(Wangetal., 2015)。设置4个氮添加处理:低氮(LN,50 kg N·hm-2a-1)、中氮(MN,100 kg N·hm-2a-1)、高氮(HN,150 kg N· hm-2a-1)和对照(CK,0 kg N·hm-2a-1)。12块样地分为4个处理,采用随机区组试验设计,每个处理3次重复。各处理样地主要林分特征和土壤性质(0~10 cm土层)见表1。自样地建成当年9月开始进行施氮,将各处理样地每年所需氮素CH4N2O等分为6份,于每年5—10月的每月中旬取1份溶解在20 L水中,使用背式喷雾器将溶液均匀喷洒在每块样地内,同时对照样地内喷洒同质量的水以避免造成水分条件差异。
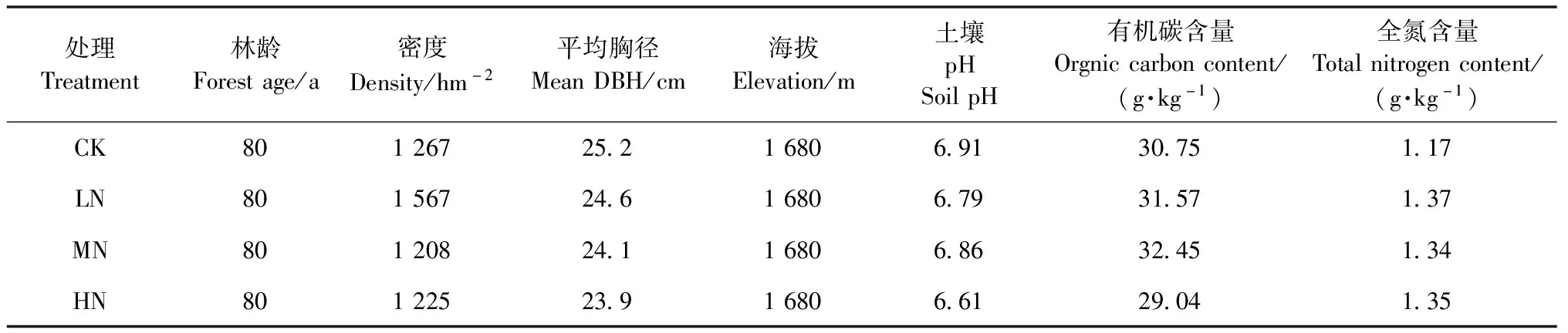
表1 各处理油松天然林样地主要林分特征和土壤(0~10 cm)性质
2009年9月,在每块固定样地内,随机设置2个2 m×2m样方,其中1个样方不做任何处理;另外一个采用壕沟法切断根系,壕沟深1 m,直到看不到根系,切断根系后用石棉瓦阻止外界根系进入,然后将土壤回填,尽量减少对土壤的扰动。同时在每个样方内插入1个PVC土壤环(内径20 cm,高10 cm),插入深度7~8 cm。2014年生长季结束后在各处理固定样地(共计12块)内随机布置4个1 m×1 m的凋落物收集框(40目尼龙网裁成的圆锥形网兜),网兜上部距离地面0.5 m。
2.2 土壤呼吸速率测定
2016—2018年,每年5—10月(由于天气原因,2016年10月末测)月底,选取晴朗天气,利用LI-8100(Li-Cor, Lincoln, NE, USA)土壤碳通量自动测量系统测定土壤呼吸速率,测量前一天使用剪刀贴地面清除土壤呼吸环内的杂草(Yanetal., 2018),同时为了最大程度地减少日内温度变化对土壤呼吸的影响,每次测量时间为10:00—16:00,同时利用LI-8100所配备的温湿度传感器测量5 cm深处的土壤温度和湿度(体积含水率)。通过挖壕法将土壤呼吸分为自养呼吸和异养呼吸,切断根系样方的土壤呼吸速率代表异养呼吸速率,不做任何处理样方与切断根系样方的土壤呼吸速率差值为自养呼吸速率。
2.3 土壤化学性质测定
在每块样地内,于2018年8月用直径2.5 cm的土钻随机(切除根系小样方除外)钻取5个0~10 cm土层的土芯,并混合为1个土样。去除植物残渣和根系后,将新鲜土样通过2 mm筛子并分为2份。一份自然风干测定土壤基本理化性质(pH值,有机碳、全氮、全磷含量);另一份在4 ℃保存,用于测定微生物生物量碳含量。土壤pH值测定采用电位法(土∶水比为1∶2.5);有机碳含量测定采用重铬酸钾氧化-外加热法;全氮含量测定采用凯氏定氮法;全磷含量测定采用钼锑抗比色法;微生物生物量碳含量测定采用氯仿熏蒸-K2SO4浸提法。
2.4 细根生物量及凋落物、细根氮磷含量测定
2018年8月,利用内径10 cm的根钻在每个2 m×2 m样方内随机(切除根系样方除外)钻取3个深10 cm的土芯。将土壤通过0.1 mm筛网,用水将筛完的根系冲洗干净后放入烘箱中,于65 ℃下烘48 h至恒质量,然后称干质量确定其细根(直径小于2 mm)生物量,然后将其粉碎以备测定细根中氮、磷含量。收集当月凋落物框中的凋落物,带回实验室进行分拣和烘干,并将凋落物研磨后过0.25 mm的筛网。在测定凋落物及细根的氮、磷含量之前,先用浓硫酸-过氧化氢消煮,定容、调色之后用全自动流动化学分析仪(Cleverchem 200+, DeChem-Tech GmbH, Germany)测定。
2.5 数据处理与分析
采用重复测量方差分析,检验不同处理和月份对土壤呼吸速率、自养呼吸速率、异养呼吸速率、土壤温度、土壤湿度的影响。采用单因素方差分析和多重比较,检验不同氮处理对土壤基本理化性质、细根生物量、细根氮磷含量及凋落物氮磷含量的影响。另外,利用公式1—3(Davidsonetal., 1998; Fangetal., 2001; Joffreetal., 2003)表示土壤呼吸速率与土壤温度及湿度间的关系,利用公式(4)(Fangetal., 2001)计算Q10值。
R=a1eb1T;
(1)
R=a2+b2W;
(2)
R=a3eb3TWc;
(3)
Q10=e10b1。
(4)
式中:R为土壤呼吸速率(μmol·m-2s-1);T为土壤温度(℃);W为土壤湿度(%);a1、a2、a3、b1、b2、b3和c为待拟合的模型参数;Q10为土壤呼吸温度敏感系数。同时采用Pearson法分析土壤呼吸速率、自养呼吸速率和异养呼吸速率与土壤化学性质、细根生物量、凋落物氮磷含量和细根氮磷含量的相关性。文中所有统计分析使用R 3.4.0进行,用Sigmaplot 12.5作图。
3 结果与分析
3.1 土壤呼吸速率、自养呼吸速率和异养呼吸速率
生长季土壤呼吸速率、自养呼吸速率和异养呼吸速率均表现为单峰曲线(图1)。2016—2018年3年间对照处理的土壤呼吸速率、异养呼吸速率和自养呼吸速率的均值(变化范围)分别为2.70(0.87~4.60)、1.86(0.53~3.86)和0.84(0.09~2.43)μmol·m-2s-1(图1)。方差分析表明,长期施氮显著降低了土壤呼吸速率和异养呼吸速率(P<0.001)(表2;图2)。相比对照,LN、MN和HN处理的土壤呼吸速率分别下降了21.9%、27.3%和29.1%,异养呼吸速率分别降低了21.8%、36.6%和31.4%(图2)。但是,氮添加并未显著改变自养呼吸速率(P>0.05)(表2,图2)。同时,月份与氮处理的交互作用对土壤呼吸速率、异养呼吸速率和自养呼吸速率影响显著(P<0.001)(表2)。
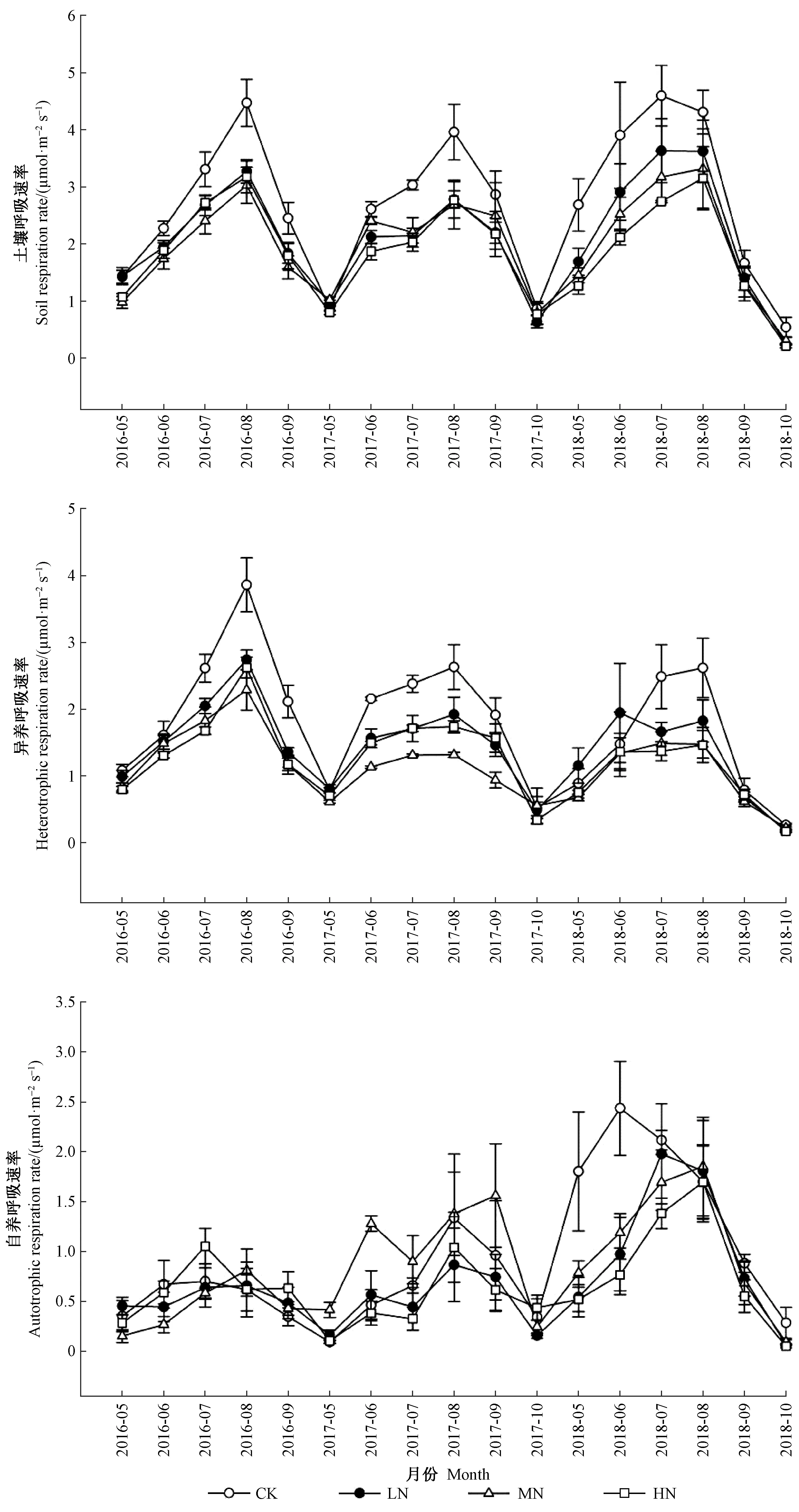
图1 2016—2018年生长季不同氮处理下土壤呼吸速率、异养呼吸速率及自养呼吸速率变化
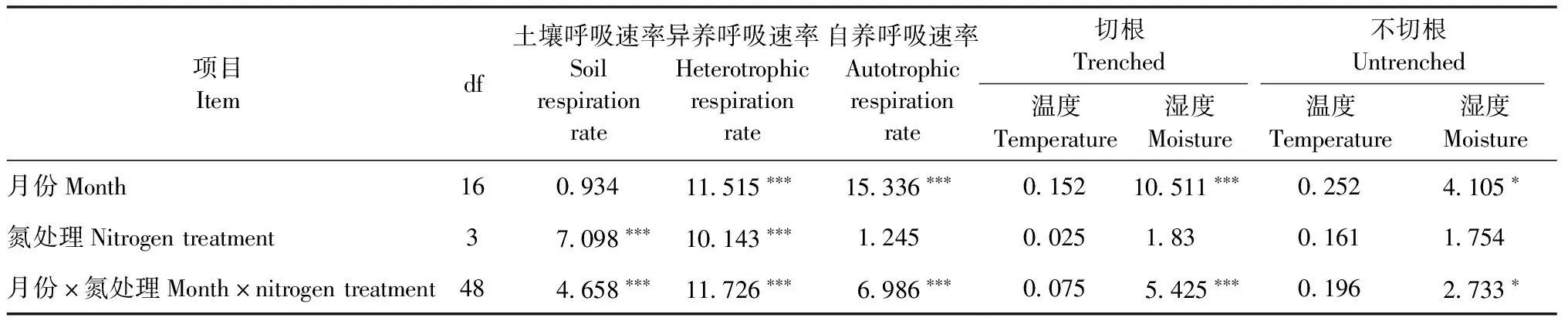
表2 土壤呼吸速率、异养呼吸速率、自养呼吸速率和土壤温湿度的重复测量方差分析F值①
3.2 自养呼吸速率和异养呼吸速率与土壤温湿度的关系
自养呼吸速率、异养呼吸速率与土壤温度均呈显著指数正相关(图3)。土壤温度分别解释自养呼吸速率的19.7%~39.6%和异养呼吸速率的63.0%~76.7%(P<0.001)(表3)。自养呼吸速率和异养呼吸速率(HN处理除外)与土壤湿度均无显著相关性(表3)。异养呼吸速率、自养呼吸速率与土壤温湿度的双变量模型表明,土壤温度和湿度共同解释了异养呼吸速率的64.9%~78.1%和自养呼吸速率的28.7%~42.0%(表 3)。与对照相比,长期施氮增加了自养呼吸的温度敏感性指数Q10,但降低了异养呼吸的温度敏感性指数Q10。
3.3 长期施氮对土壤化学性质、细根生物量、凋落物氮磷含量和细根氮磷含量的影响
表4表明,长期施氮显著降低了土壤pH值(表4),LN、MN和HN处理比对照分别降低了0.07、0.37和0.78个单位,并使土壤微生物生物量碳含量显著降低(P<0.05),LN、MN、HN处理比对照分别降低了11.3%、14.5%和14.7%,但长期施氮未显著改变土壤有机碳、全氮和全磷含量,也未显著改变细根生物量以及土壤、凋落物和细根的氮磷含量。

表4 长期施氮对油松林土壤化学性质、细根生物量、凋落物氮磷含量和细根氮磷含量的影响①
相关分析表明(表5):土壤呼吸速率与土壤微生物生物量碳极显著正相关(P<0.01),与凋落物氮含量显著正相关(P<0.05);异养呼吸速率与土壤微生物生物量含量极显著正相关(P<0.01);自养呼吸速率与细根生物量显著正相关(P<0.05)。
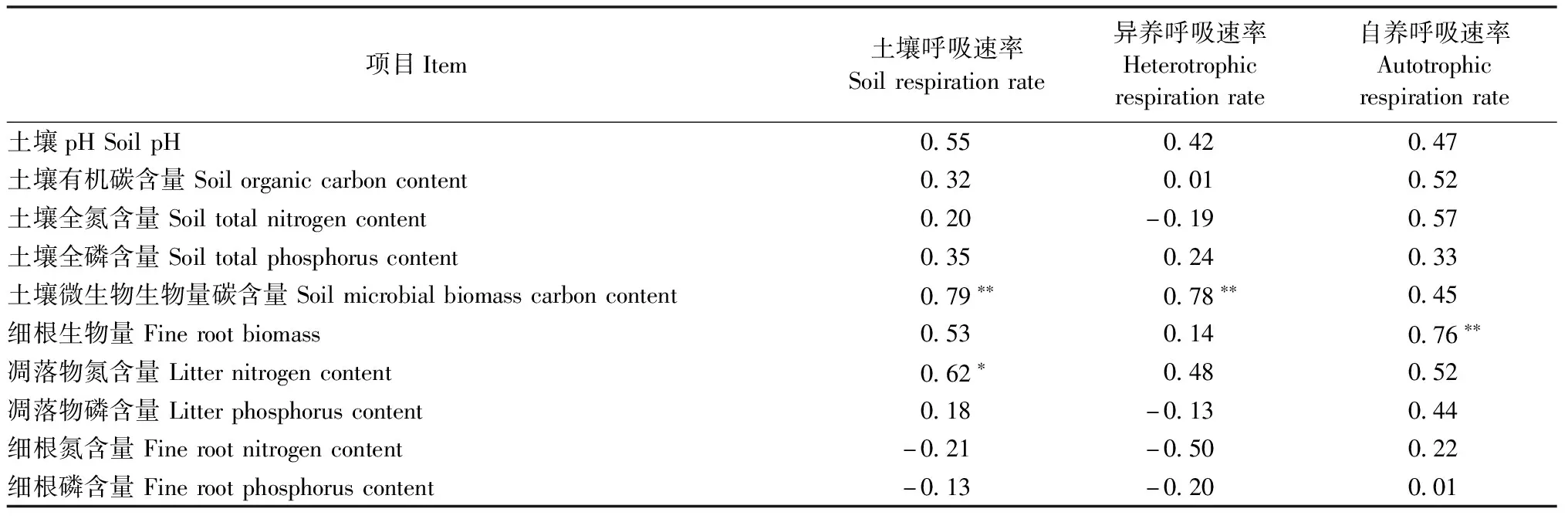
表5 长期施氮后土壤呼吸及其组分与土壤化学性质、细根生物量、凋落物氮磷含量和细根氮磷含量的Pearson相关分析①
4 讨论
4.1 长期施氮对土壤呼吸及其组分的影响
本研究发现,相比对照,10年氮添加导致土壤呼吸速率显著降低了26.1%(图2)。前人研究亦表明氮沉降对温带森林土壤呼吸具有抑制作用(Bowdenetal., 2004; Janssensetal., 2010; Sunetal., 2014; Yanetal., 2018)。本研究结果高于中国北方温带华北落叶松(Larixprincipis-rupprechtii)林的结果,其研究发现,4和7年氮添加(50 kg N·hm-2a-1)试验分别使土壤呼吸速率降低了14%(Sunetal., 2014)和11.2%(Yanetal., 2018)。但与美国温带红松(Pinuskoraiensis)林氮添加早期研究结果基本一致,低氮(50 kg N·hm-2a-1)和高氮(150 kg N· hm-2a-1)处理的第2年分别使土壤呼吸速率降低了21.7%和30.1%(Bowdenetal., 2004);而13年低氮和高氮处理分别使土壤呼吸速率降低了34%和41%(Bowdenetal., 2004)。本研究样地氮添加早期(第3~7年)研究发现,外源氮输入未显著改变土壤呼吸速率(Zhaoetal.,2018),这主要是由于本研究样地在施氮开始前土壤氮含量较低(Wangetal., 2015),7年氮添加导致土壤氮含量增加(Zhaoetal.,2018),这一方面促进根系生长而增加自养呼吸速率,另一方面氮添加导致土壤微生物生物量降低从而抑制异养呼吸速率,二者相互抵消导致土壤呼吸速率未显著变化(Zhaoetal., 2018)。而10年施氮后土壤呼吸速率明显降低,主要是因长期施氮显著降低了异养呼吸速率29.9%,对自养呼吸速率无显著影响(图2)。
本研究中长期施氮后的异养呼吸速率降低主要与土壤微生物生物量碳含量显著下降(表4)有关,这与最新的全球整合分析结果一致(Wangetal., 2018; Zhangetal., 2018)。全球尺度上,氮添加导致土壤微生物生物量碳含量下降了11%(Wangetal., 2018),从而使异养呼吸速率下降了8.1%(Zhangetal., 2018)。异养呼吸速率降低表明土壤微生物对土壤有机质分解能力的下降,这将利于土壤固碳(Yanetal., 2018)。另外,长期施氮也会通过形成不易被微生物降解的化合物(Chenetal., 2017)来降低土壤微生物碳利用率(Sinsabaughetal., 2005; Lietal., 2018)和纤维素酶活性(刘星等, 2015; 张艺等, 2017; Chenetal., 2018; 范珍珍等, 2018; Zhaoetal., 2018)以及降低微生物多样性而降低异养呼吸(Ramirezetal., 2012; Zhengetal., 2018)。本研究的10年长期施氮未改变自养呼吸速率(P>0.05);而Zhao等(2018)发现本样地在施氮3~7年时自养呼吸速率显著增加,主要是通过增加根系生物量而增加自养呼吸速率。这可能与根系生长和活动随施氮年限表现出不同的敏感性有关(Nadelhoffer, 2000)。细根生物量变化与土壤氮含量有很大关系,本研究样地在施氮前期受土壤低氮限制(Wangetal., 2015),氮输入将刺激细根生物量增加进而增加自养呼吸,但10年氮添加使土壤出现氮饱和(表4),之后植物无法吸收更多的氮(Yanetal., 2018),同时过多氮输入也可能加快根系周转即缩短根系寿命(Nadelhoffer, 2000; Xiaetal., 2017),因此氮输入不会明显改变细根生物量(Bowdenetal., 2004)。
4.2 不同施氮处理下土壤呼吸及其组分与土壤温湿度的关系
本研究表明,土壤温度是自养呼吸速率和异养呼吸速率的主要控制因素(图3),这与国内外研究结果一致(常建国等, 2006; Laganièreetal., 2012; 汪金松等, 2012; 李化山等, 2014; 陈平等,2018; Yanetal., 2018; Zhaoetal., 2018)。同时,土壤温度对异养呼吸速率的解释度高于对自养呼吸速率的解释度,表明土壤微生物受土壤温度影响更大(Yanetal., 2018),而根系活性同时受多因素调控(Booneetal., 1998; Sunetal., 2014)。
自养呼吸速率和异养呼吸与土壤湿度未呈现显著线性关系,这可能与当地气候有关,本地四季明显,生长季内降水充足,因此较少受土壤湿度限制,这也与本研究样地之前研究结果一致(Zhaoetal., 2018)。同时,土壤温度和土壤湿度的双因子模型对自养呼吸速率和异养呼吸速率解释度均大于单因子模型(表3),表明土壤微生物代谢活动及根系活性均受土壤温湿度的共同调控(汪金松等, 2013)。
本研究中对照处理的自养呼吸和异养呼吸的Q10值分别为2.19和2.72(表3),这与温带森林的Q10值(2.7±1.7)相近(Chenetal., 2005)。长期施氮使自养呼吸的Q10值增加(Dengetal., 2010),这是因为在养分匮乏地区(如北美的温带森林)长期施氮会增加土壤有机质含量进而增加底物供应(Sinsabaughetal., 2005)。而这与短期氮沉降使自养呼吸Q10值降低不同(Sunetal., 2014),短期氮添加通过增加地表凋落物进而减少光合产物对根系的分配(Savageetal., 2013),根系呼吸底物基质降低是自养呼吸Q10值降低的一个主要原因。长期施氮使异养呼吸的Q10值降低,这与短期氮沉降对异养呼吸Q10值的影响一致(Sunetal., 2014; Zhaoetal., 2018),主要是因施氮降低了凋落物和土壤中的微生物酶活性(Maetal., 2013)进而降低分解作用对温度的敏感性(Zhaoetal., 2018),施氮还能改变呼吸底物组分,使其产生更多的顽固性有机质,其与氮结合会产生难降解的酚类化合物和微生物酶化合物(Chenetal., 2018),使异养呼吸底物的可利用性降低,进而降低异养呼吸的温度敏感性;同时,顽固性有机质的增加也将使土壤有机碳含量增加(Janssensetal., 2010)。总体上,Q10对施氮的响应受土壤养分含量、环境因子和生态系统类型等多因素影响(Zhengetal., 2009),要准确判断氮沉降对Q10的影响需综合考虑多因素作用。
5 结论
长期施氮(10年)使山西省太岳山油松林的土壤呼吸速率和异养呼吸速率分别降低26.1%和29.9%,而未明显改变土壤自养呼吸速率,这与早期施氮(3~7年)增加自养呼吸不一致。长期施氮提高了自养呼吸的Q10值,这与短期施氮减少光合产物对根系的分配进而降低自养呼吸Q10不一致;长期施氮降低了异养呼吸Q10值,这与短期施氮对异养呼吸Q10值的影响一致。长期施氮与短期氮添加对自养呼吸及异养呼吸Q10的影响不一致,在未来氮沉降持续增加的背景下,亟需开展土壤呼吸对氮添加响应的更长期观测和机理研究。
