甜高粱对镉胁迫的生理生化响应及镉富集研究
2021-03-12郝正刚赵会君魏玉清曾周琦王志恒
郝正刚, 赵会君, 魏玉清, 曾周琦, 王志恒
(北方民族大学生物科学与工程学院, 银川 750021)
土壤重金属污染是全球面临的重大环境污染之一。近年来,由于工业“三废”的不科学排放,使得土壤重金属含量显著增加[1]。目前,我国镉(Cd)污染情况比较严重,2014年《全国土壤污染状况调查公报》指出,我国耕地土壤点位超标率达到了19.4%,其中重金属镉污染点位超标率占了7.0%,位居无机污染物超标率第一位[2]。刘凤莲等[3]调查分析指出,宁夏农村土壤镉含量平均值为0.187 mg·kg-1,高于《中国土壤元素背景值》中宁夏地区的0.112 mg·kg-1,镉含量超标倍数为1.67,说明宁夏农村土壤镉元素也有富集现象。
重金属镉不是植物生长所必需的元素,但是被吸收后会在植物体内积累,最终有可能通过食物链进入人体,对人类的健康造成危害[4-5]。20世纪40年代,日本富士山县神通川流域发生的“痛痛病”事件就是人类食用了含镉的大米所致[6]。因此,为了人类健康,修复被重金属污染的土壤显得尤为重要,但是常规的物理、化学等方法对土壤破坏力度大、成本高、费用昂贵。相比之下,生物修复以对土壤破坏力度小、安全性高、成本低的优点越来越受到学术界的关注[7]。近年来,关于蜈蚣草(Pterisvittata)、向日葵(Helianthusannuus)、芥菜(Brassicajuncea)等植物修复重金属污染的研究较多,但有关甜高粱(SorgghumbicolorL. Moench)的报道甚少[8]。甜高粱具有生物产量高、适应性强、耐盐碱等特性,其作为一种新兴的生物质能源作物受到广泛重视[1]。相关研究主要报道了镉胁迫对甜高粱种子萌发、幼苗生长、抗氧化酶活性、光合参数、超微结构及微量元素吸收的影响[9]。在前人研究基础上,本研究以甜高粱为对象,通过盆栽水培,探讨了镉胁迫对甜高粱生长、生理生化的影响及甜高粱对重金属吸收效应,旨在为利用边际性土地和重金属污染土壤发展能源作物甜高粱提供参考依据。
1 材料与方法
1.1 试验材料
试验材料为甜高粱杂交品种辽甜1号,由辽宁省农业科学院提供。
1.2 试验方法
试验采用盆栽试验方法,于2018年在北方民族大学植物逆境生理室进行。选取大小均匀、健康饱满的种子,自来水反复冲洗后,用10%次氯酸钠消毒10 min。常温下,用灭菌水冲洗并浸泡8 h,摆放在铺有2层滤纸的培养皿中,置于27 ℃ 恒温生化培养箱内催芽。将催芽后的种子移至装有等量石英砂(直径约为0.3 cm)的塑料花盆(11.7 cm×7.7 cm×10.7 cm)中,每盆8粒种子,置于28 ℃、光照周期为13 h/11 h(昼/夜)的光照培养箱内进行光照培养。待苗高为3叶1心时,每盆定苗6株,并转至自然光条件下培养。培养过程中,每周更换1次Hoagland营养液。培养4周后,将幼苗随机分成4组,每组6盆即重复6次。以镉溶液(CdCl2·2.5H20)模拟重金属胁迫环境,共设置5个梯度水平:CK(0 μmol·L-1)、T1(50 μmol·L-1)、T2(100 μmol·L-1)、T3(200 μmol·L-1)、T4(300 μmol·L-1),每组加处理液2 L,胁迫过程中,根据墒情补加等量的营养液和水。分别在处理的第3、7、11 d采样,测定叶片的保护酶活性,并对处理后第3 d的叶片采用垂直板聚丙烯酰胺凝胶电泳(PAGE)进行同工酶分析;于第30 d测定叶绿素含量、光合参数、叶绿素荧光参数和净光合速率(net photosynthetic rate,Pn)对光强的响应曲线;第31 d测定生物量,并对地上部和地下部进行前处理,用于镉含量的测定。
1.3 测定项目及方法
1.3.1保护酶活性的测定及同工酶分析 称取0.5 g鲜叶,在冰浴条件下加酶提取液0.1 mol·L-1Tris-HCl缓冲液[每100 mL含半胱氨酸0.073 g、抗坏血酸0.105 g、乙二胺四乙酸二钠(Na2-EDTA) 0.075 g、甘油10 mL、1mol·L-1HCl 5.84 mL,pH 8.0]2 mL,快速研磨至匀浆。然后在4 ℃下,于低温冷冻离心机中13 000 r·min-1离心20 min,上清液即为酶提取液。超氧化物歧化酶(superoxide dismutase,SOD)活性测定采用邵从本等[10]方法;过氧化物酶(peroxidase,POD)活性测定采用李合生[11]方法;过氧化氢酶(catalase,CAT)活性测定采用李仕飞等[12]方法;抗坏血酸过氧化物酶(ascorbate peroxidase,APX)活性测定参照孙云[13]方法。
同工酶分析:分离胶浓度CAT为7.5%,POD、SOD、APX为10%;浓缩胶浓度为3%。SOD同工酶染色采用四氮哇蓝NBT法[14];POD同工酶染色采用联苯胺法[14];CAT同工酶染色采用淀粉法[15];APX同工酶染色参考文献[16-17]方法。
迁移率(Band RF)=固定染色后凝胶中酶蛋白区带的迁移距离/固定染色中指示剂的迁移距离。
1.3.2叶绿素及光合参数的测定 叶绿素含量的测定:参照陈福明等[18]的混合液提取法。
光合参数及响应曲线的测定:选择生长健康、完全展开的甜高粱叶片,于上午09:00—11:00采用LI-6400XT便携式光合作用测定系统(美国LI-COR公司)进行测定,每处理取3次重复。光合参数净光合速率(Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentrations,Ci)、蒸腾速率(net photosynthetic rate,Tr)在控制条件下进行,设定光强(photosynthetic active radiation intensity,PAR)为1 000 μmol·m-2·s-1固定红蓝光源,Flow为500 μmol·s-1,CO2R为400 μmol·mol-1,Temp为25 ℃,等待数据稳定后开始记录数据。测定光响应曲线时,采用自动测量程序,使用CO2注入系统浓度控制为(CO2R)400 μmol·mol-1,光强由强到弱依次设定PAR为2 000、1 500、1 200、1 000、750、500、250、150、100、60、20、0 μmol·m-2·s-1,Temp为20 ℃,测定胁迫下叶片的Pn,测定前用800 μmol·m-2·s-1冷光源诱导20 min,在每个光强下平衡120~300 s后开始测定。
荧光参数的测定:采用LI-6400XT便携式光合作用测定系统的荧光叶室进行测定,选择生长健康、完全展开的甜高粱叶片,全部做标记后,暗适应一个晚上,测定原初光能转换效率(Fv/Fm),然后在自然光下活化,测定Fs、Fm′和Fo′,计算光化学猝灭系数(qP)、光合电子传递效率(φPSⅡ)、非光化学猝灭系数(NPQ)。
1.3.3生物量及镉元素含量的测定 将甜高粱苗从花盆中移除,用剪刀使地上部与地下部分离,量取地上部长度即株高(cm),用电子天平称取地上部鲜重(g)。将地上部与地下部用20 mmol·L-1Na2-EDTA 浸泡 1 h,除去根表面粘附的金属离子,然后用自来水冲洗3遍,再用蒸馏水冲洗2遍,随后在105 ℃杀青 30 min,在80 ℃下烘干至恒重,称取地上部干重(g)与地下部干重(g)。
将烘干至恒重的地上部与地下部分别用粉碎机粉碎并过60目筛,称取0.5 g样品置于微波消解仪(MARS5,美国CEM公司)衬罐内,按顺序滴加3 mL超纯水、5 mL优级纯浓硝酸、2~3滴H2O2,加盖后置于SEM微波消解炉里,消解结束经放气后取出内衬罐,将消解液用定量滤纸过滤并定容至50 mL摇匀,最后用全谱直读等离子体发射光谱仪(iCAP 6300,美国Thermo Fisher SCIENTIFIC公司)测定分析镉元素的含量。
1.4 数据统计
数据采用3个重复的平均值±标准误差,利用 Microsoft Excel 2003和GraphPad Prism 5.0进行数据处理和方差分析,采用SPSS Statistics 21拟合光响应曲线,采用经典的Farquhar模型来拟合[19],具体操作参照陆燕元等[20]的方法,该模型的理论公式如下。
式中,Pn为净光合速率,I为光合有效辐射强度,φ为表观量子效率,Pmax为最大净光合速率,Rd为暗呼吸速率,θ为非直角双曲线的凸度(曲角)。
在低光强下,Pn随I的增加呈线性增高,通过对200 μmol·m-2·s-1以下所采集的数据进行直线回归分析,得出的线性方程与X轴的交点数值即为光补偿点(light compensation point,LCP),而与Pmax的交点在X轴上的数值则为光饱和点(light saturation point,LSP)。
2 结果与分析
2.1 镉胁迫对甜高粱生长的影响
表1显示,甜高粱幼苗在不同浓度的镉处理下,地上部鲜重、株高、地上部干重、地下部干重有不同的响应。总体表现为各指标随着镉浓度的增加呈降低的趋势。各处理组较对照组均有显著差异(P<0.05),T4处理下生物量降至最低,地上部鲜重、株高、地上部干重、地下部干重较对照分别降低了55.41%、24.66%、56.50%、71.57%。
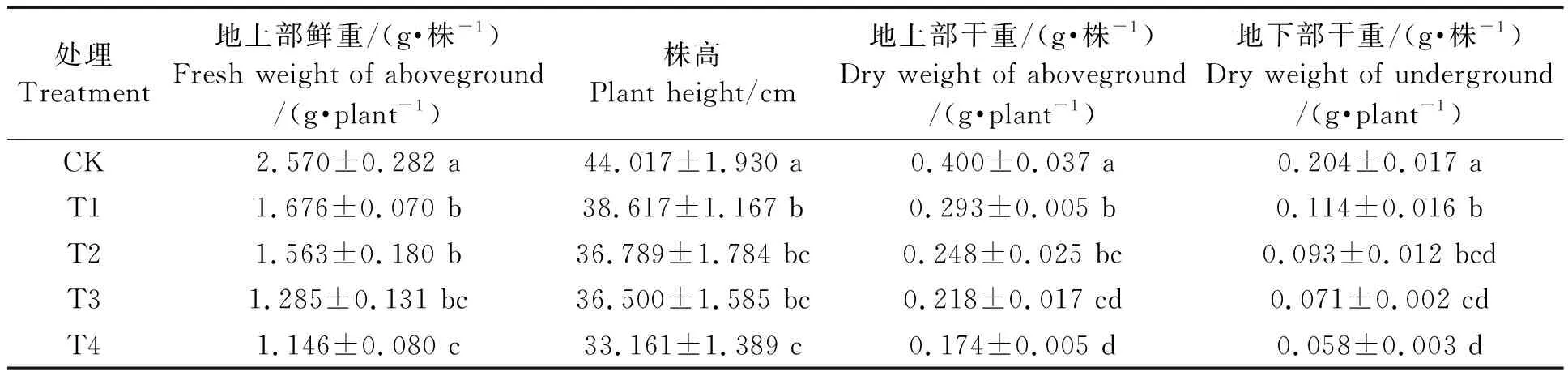
表1 镉胁迫对甜高粱生长的影响Table 1 Effects of cadmium sress on the growth of sweet sorghum
2.2 镉胁迫对甜高粱抗氧化酶的影响
2.2.1不同处理下抗氧化酶活性 由图1可知,镉胁迫下,甜高粱幼苗超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、过氧化物酶(抗坏血酸APX)活性的响应不同。SOD活性随镉浓度升高呈先上升后下降的趋势,且随着胁迫时间的延长,处理组显著高于对照(P<0.05)。POD活性在处理第3、11 d随镉浓度升高呈上升的趋势,而在第7 d呈先上升后下降的趋势;胁迫期间,T1处理与对照差异不显著,而T2、T3、T4处理与对照差异显著(P<0.05)。CAT活性随镉浓度升高呈下降的趋势,且伴随胁迫时间的延长,处理组显著低于对照(P<0.05)。APX活性在胁迫第3 d,随着镉浓度升高呈先下降后上升再下降的趋势,处理组与对照无显著性差异;在胁迫第7 d,随着镉浓度升高呈先下降后上升的趋势,T1和T2处理较对照差异显著(P<0.05);在胁迫的第11 d,随着镉浓度升高呈下降的趋势,各处理组与对照组均有显著差异(P<0.05)。
2.2.2胁迫早期过氧化物同工酶电泳结果 对胁迫第3 d的甜高粱叶片进行抗氧化同工酶分析,结果见图2。在SOD同工酶方面,处理组的S5、S4、S3条带较对照组色度加深;T2、T3处理下出现了新的S2条带,且随着处理浓度的增加,色度先加深后消失;而S1则随着处理浓度的增加色度先变浅后加深,其中T1处理色度最浅,这与胁迫下SOD酶活性的测定结果一致。在POD同工酶方面,处理组的P9、P8、P7、P6、P5条带较对照亮度加深;P4、P3、P1条带均随着处理浓度的增加亮度先加深后变浅,其中P4、P3在T3处理下亮度最深,P1条带在T1处理下最深;而P2条带在CK、T4处理下无表达,在T3处理下亮度最深,这也进一步证实了胁迫下POD酶活性呈上升的趋势。在APX同工酶方面,A6、A5、A4、A3、A2条带均随着处理浓度的增加,亮度先加深后变浅再加深,其中于T1处理亮度最深;而A1条带则随着处理浓度的增加亮度先变浅后加深再变浅,其中在T2处理下亮度最深。在CAT同工酶方面,C2、C1条带处理组较对照组亮度均变浅,其中C2条带在T2处理下亮度最浅,而C2在T3处理下亮度最浅,这与胁迫下CAT酶活性呈下降趋势的结果一致。
2.3 镉胁迫对甜高粱光合作用的影响
2.3.1对甜高粱叶绿素含量的影响 由图3可知,镉胁迫下,甜高粱叶片叶绿素a、b及a+b的含量随着镉浓度的增加而降低,处理组与对照组相比差异均显著(P<0.05),其中T4处理下降至最低,较对照分别降低了63.21%、67.61%、64.34%。叶绿素a/b随着镉浓度的增加呈先上升后下降的趋势,处理组与对照组相比差异均显著(P<0.05),其中在T3处理下最高,较对照升高了17.24%。
2.3.2对甜高粱叶片光合参数的影响 由图4可知,镉胁迫会严重影响甜高粱叶片光合作用的进行。净光合速率(Pn)随着镉浓度的增加而呈降低的趋势,其中T1处理与对照无显著性差异,T2、T3、T4处理较对照差异均显著(P<0.05),分别降低了31.05%、45.29%、61.24%。气孔导度(Gs)、蒸腾速率(Tr)随着镉浓度的增加也呈降低的趋势,处理组与对照组相比差异均显著(P<0.05)。随着镉浓度的增加,胞间CO2浓度(Ci)呈上升的趋势,T1处理与对照无显著性差异,其他处理较对照差异均显著(P<0.05)。
2.3.3对甜高粱叶片荧光参数的影响 叶绿素荧光参数是被用来描述植物光合系统生理状况和叶绿素结构状态的指标。由图5可知,叶片原初光能转化效率(Fv/Fm)、光化学淬灭系数(qP)随着镉浓度的增加呈下降的趋势,其中T3、T4处理较对照均有显著性差异(P<0.05)。光合电子传递效率(φPSⅡ)随着镉浓度的增加也呈降低的趋势,其中T2、T3、T4处理较对照分别降低了18.82%、34.71%、61.35%,且均有显著差异(P<0.05)。而非光化学淬灭系数(NPQ)则随着镉浓度的增加呈上升的趋势,其中T1、T2处理较对照无显著差异,T3、T4较对照分别上升了84.41%、90.32%,且均有显著差异(P<0.05)。
2.3.4对甜高粱幼苗叶片光合响应曲线变化及相关参数的影响 图6显示,光强PAR在0~500 μmol·m-2·s-1左右时,净光合速率Pn快速上升,当光强达到500 μmol·m-2·s-1左右时上升的幅度较弱。PAR在200 μmol·m-2·s-1以下时,各处理下光响应曲线都呈线性增长,其中直线方程的斜率为表观量子效率(φ),主要反映甜高粱叶片对弱光的利用能力。对各处理下的叶片光响应曲线的变化趋势进一步分析发现,T1、T2处理较对照无显著差异,而T3、T4较对照具有显著差异(P<0.05)。
由表2可以看出,镉胁迫下甜高粱叶片最大净光合速率(Pmax)、表观量子效率(φ)、暗呼吸速率(Rd)、光饱和点(LSP)均随着镉浓度的增加呈降低的趋势。其中,各处理组的Pmax较对照均有显著性差异(P<0.05),分别降低了31.05%、38.13%、70.83%、84.15%。T3、T4处理下φ、Rd较对照均有显著性差异(P<0.05),φ分别降低了50.00%、83.33%,Rd分别降低了37.31%、55.22%。T2、T3、T4处理下LSP较对照均有显著性差异(P<0.05),分别降低了25.71%、43.32%、51.81%。而镉胁迫下甜高粱叶片光补偿点(LCP)随着镉浓度的增加呈上升的趋势,其中T4处理较对照升高了89.29%,且具有显著性差异。

表2 镉胁迫对甜高粱叶片光合响应曲线相关参数的影响Table 2 Effects of cadmium stress on related parameters of photosynthetic response curves of sweet sorghum leaf
2.4 甜高粱对镉的富集特征
富集量(地上部、地下部生物量(干重)与其镉浓度的乘积)和富集率(地上部、地下部富集量占处理液镉含量的百分比)均可以反映甜高粱在镉胁迫下对镉的富集能力。由表3可以看出,同一处理下,甜高粱地下部镉浓度均高于地上部。甜高粱植株地上部、地下部的镉浓度均随着处理梯度的增加而呈上升的趋势,其中,地上部的镉浓度在T4处理下与T3、T2、T1差异显著(P<0.05);地下部的镉浓度T1处理与T4、T3、T2相比差异均显著(P<0.05)。地上部富集量随着处理梯度的增加而呈下降的趋势,而地下部富集量则随着处理梯度的增加而呈先上升再下降的趋势,但处理组之间均无显著性差异(P>0.05);地上部富集率、地下部富集率、单株富集率均随着处理浓度的增加而降低,处理组之间差异均显著(P<0.05)。

表3 甜高粱对镉的富集特征Table 3 Enrichment characteristics of cadmium by sweet sorghum
3 讨论
植物体在其生命活动中不断与环境进行物质、能量及信息的交换,其生长发育受到环境制约[21]。植物通过不断调节自身的生理状态以适应逆境条件带来的胁迫,保护酶活性调节是植物在逆境条件下防御自由基氧化损伤的重要机制之一,其中,超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)、抗坏血酸过氧化物酶(APX)是植物内源的活性氧清除剂,在逆境中只有保持相对较高的活性才能缓解外界对自身膜结构的伤害[22]。镉进入植物体后,破坏了植物正常的生理生化过程,活性氧产生。当胁迫发生后,植物会采取各种措施提高抗性以适应不良环境,但当胁迫发生超过植物忍受的极限时,其防御措施也就相应减弱,乃至死亡。因此,通过分析保护酶的活性可以初步了解植物对胁迫的适应性[23]。本研究结果显示,甜高粱SOD活性呈现低浓度增加、高浓度下降的趋势,POD活性随着处理浓度的增加呈升高的趋势,这与李冬琴等[24]的结论一致。而CAT活性则随着处理浓度的增加而降低,这与Romero-puertas等[25]的结论一致;APX活性随着处理浓度的增加在不同时期呈现不同的趋势,这可能与APX活性和基因表达不受镉的影响有关[26]。这说明,甜高粱在镉胁迫下主要是SOD、POD发挥抗氧化功能,并伴随着CAT活性的降低和APX的增加或减少,抗氧化酶类之间保持良好的平衡将胁迫产生的活性氧维持在平衡的状态。同工酶是基因表达的直接产物,稳定性较高,在植物体的合成和表达主要受遗传因子和环境二者的影响[27-28]。因此,通过同工酶的分析,可以了解甜高粱对胁迫的适应及基因的表达情况,同时也能进一步印证各抗氧化酶活性对镉的响应。本研究显示,镉胁迫下甜高粱SOD酶谱多条带的表达不尽相同,在T2、T3处理下出现了新的S2条带;POD的很多条带随处理浓度的增加而亮度加深,在T1、T2、T3出现了新的P2条带;而CAT、APX酶谱没出现新的条带,但在各组处理中各个条带的表达量均不相同。这说明,镉胁迫会打破胁迫前保护酶之间的平衡,甜高粱通过增加或抑制自身一些同工酶的合成与表达、甚至被诱导某个基因的表达产生新的酶谱条带来抵御镉胁迫,酶带的加深或变浅、新增或缺失与甜高粱抵御镉毒害有关,是甜高粱对不同程度镉胁迫的适应机制。
植物作为生态系统中的生产者,依靠光合作用不仅可以获得能量,同时也合成营养物质。植物叶片中存在一定量的叶绿素蛋白复合物,在光能的吸收、传递、转换中起着重要的作用,因此,叶绿素含量的高低和植物光合作用的强弱密切相关[29-30]。本研究表明,镉胁迫使得甜高粱叶片叶绿素a、b及总含量、净光合速率(Pn),气孔导度(Gs)和蒸腾速率(Tr)均下降,而胞间CO2浓度(Ci)则增加,且随着胁迫时间的延长变化更显著,这与前人利用草莓[5]、烟草[31]得出的研究结果相似。相关研究表明,影响植物光合作用的主要因素包括气孔因素和非气孔因素,当植物叶片Pn、Gs、Ci同时降低,则说明Pn的下降主要是气孔因素导致的;当Pn、Gs下降,Ci升高,则表明Pn下降的主要原因是非气孔因素的限制[19]。本镉胁迫下,Pn、Gs降低而Ci升高,这说明镉胁迫下甜高粱幼苗光合作用的降低不是气孔导度下降使得CO2供应减少导致的,可能是叶绿素含量降低,使得叶肉细胞光化学活性受到限制而影响了CO2同化利用造成的。镉胁迫使得叶片叶绿素含量降低的原因可能是镉被植物吸收后进入叶绿体,取代了叶绿体中的Fe2+、Zn2+和Mg2+,使得叶绿体的亚显微结构发生改变,主要表现在基粒垛叠结构的解体和片层系统相关的基质减少[32-33]。
叶绿素荧光和光合作用的关系十分密切,当强光持续照射植物时,为了避免叶绿体吸收光能超过光合作用过程中光化学反应的消耗能力以及过量的光能灼伤损害光合机构,荧光起到了重要的保护作用,主要表现为一部分光能以荧光的形式被耗散掉[34]。叶绿素荧光参数可以反映叶片PSⅡ反应中心的状态,与“表观性”的气体交换指标相比,叶绿素荧光参数具有反映“内在性”特点[35],其中Fv/Fm是植物暗适应下PSⅡ反应中心完全开放时的最大光化学效率,它反映了PSⅡ反应中心最大光能转化率;φPSⅡ是实际光化学效率,反映植物在照光条件下PSⅡ反应中心部分关闭的情况下的实际光化学效率,表示光化学反应消耗的能量占叶片吸收光能的比例[36];光化学淬灭系数(qP)反映PSⅡ反应中心的开放程度,反映叶片中捕光激发能用于光化学反应的程度[37];而非光化学淬灭系数(NPQ)则代表植物热耗散的能力。本研究结果显示,镉胁迫下甜高粱叶片Fv/Fm、φPSⅡ、qP均减小,而NPQ则增加,这与李伶[38]、曹玲等[39]得出的结果相同,说明镉胁迫下甜高粱叶片PSⅡ原初光化学活性受到影响,PSⅡ活性中心被损伤或破坏,使得反应中心捕获激发能下降,PSⅡ功能受到抑制。因此,通过热耗散途径来耗掉过剩的光能来尽量保护光合机构免受伤害,使得用于光化学反应的比例减少,导致甜高粱叶片CO2同化能力下降。
植物对光强的响应曲线变化也是研究植物光合作用的手段之一,同一植物的光合响应相关参数不是固定的数值,会随着外界环境的变化而变化[40-41]。其中,叶片Pmax反映植物最大净光合能力,其大小受Rusbico活性和电子传递效率的影响[42-43]。表观量子效率φ的大小反映了叶片在弱光下吸收转换及利用光能的能力,该值越大说明叶片利用弱光的能力越强,光能转化率较高[44-45]。而LCP、LSP则反映植物叶片对光的利用能力,是植物需光特性的体现,相关研究表明,较低的LCP、LSP使植物在有限的光照条件下以最大能力利用低光量子密度,这与低光强下单位面积叶绿素含量升高和暗呼吸速率降低有关[46]。本研究中,镉胁迫降低了甜高粱叶片Pmax、φ、Rd、LSP,使得LCP升高,这说明镉胁迫降低了甜高粱叶片Rusbico活性和电子传递效率,使得叶片对光的吸收、传递、转化能力下降,从而降低甜高粱叶片对光的利用能力。
甜高粱作为能源材料被用于修复土壤镉污染,其生物量的高低以及对镉的富集量是评价修复能力的重要指标。本试验条件下,甜高粱的生物量随胁迫浓度的增加显著降低,而地上部与地下部的镉浓度则显著升高,且地上部低于地下部,这与前人的研究结果一致[47-48]。本研究中,镉胁迫造成了甜高粱光合作用能力的下降是生物量降低的主要原因之一。对地上部与地下部的镉浓度并结合生物量进一步分析得出,地上部富集量与地下部富集量伴随胁迫浓度的增加而降低,同时地上部、地下部及单株富集率也降低;低浓度(50 μmol·L-1)胁迫下地上部富集量、富集率高于地下部,同时单株富集率最高达6.05%,而高浓度胁迫下则富集量、富集率趋于相同。这主要是由于镉胁迫下甜高粱地上部、地下部的生物量均降低,虽然地上部、地下部的镉浓度均随着处理梯度的增加而上升,但是由于其生物量降低,从而导致了富集能力下降。
综上所述,在镉胁迫下,甜高粱通过调控SOD、POD、CAT、APX相关基因的表达来提高SOD、POD酶活性,降低CAT酶活性,以此维持抗氧化酶类之间的动态平衡,从而减轻胁迫带来的活性氧对植株造成的伤害,保证植株细胞的正常代谢。镉胁迫是通过降低叶片中叶绿素的含量,使得PSⅡ原初光能转化效率降低,电子传递受到抑制,使得叶片对CO2的吸收、固定和同化效率下降,以此来降低甜高粱幼苗的光合能力,甜高粱叶绿体为了避免过多、剩余的光能灼伤光合机构,因此增加了热耗散来适应镉胁迫。镉胁迫降低了甜高粱地上部与地下部的生物量,使得地上部与地下部的镉浓度升高;低浓度胁迫下富集能力较强,单株富集率伴随处理浓度的增加而降低,这主要跟胁迫后生物量的降低有关。
本研究以甜高粱幼苗为研究对象,采用水培模拟胁迫环境,探讨和分析了镉胁迫对甜高粱生理生化的影响,在此基础上评价了对镉的吸收效应。胁迫时间较短,没有考虑成熟期的甜高粱对镉的富集能力和镉胁迫对其汁液生产乙醇的影响。鉴于甜高粱作为能源材料,对重金属的富集效率与其生物量有关,因此,土壤镉胁迫下成熟期甜高粱的镉富集能力、汁液发酵生产乙醇的得率和生产乙醇后含镉的废弃秸秆的处理等问题是后续的研究方向。
