桃果实中MiR167a与其靶基因的鉴定及对IAA的响应分析
2021-03-10张彦苹朱旭东刘照坤李庆魁
张彦苹,朱旭东,王 晨,刘照坤,李庆魁
(1 苏州农业职业技术学院, 江苏苏州 215008;2 南京农业大学 园艺学院, 南京 210095;3 苏州市农业科学院, 江苏苏州 215000)
植物的性状由基因控制,基因的表达被多种调节因子高度调节,以确保各组织的正常发育和功能以及适应外界的变化。MicroRNA(miRNA)是一类长度为19~24个核苷酸的内源非编码小RNA分子[1],已被证明通过参与转录后翻译在植物的生长、发育、代谢、信号转导等多方面起调控作用。miRNA具有在转录后水平与靶基因互补配对,通过剪切或翻译抑制的方式调控靶基因表达的功能。在植物中,目前的研究发现许多生长素相关表型出现在miRNA-靶基因互作体系中,说明植物miRNA直接或者间接地参与了生长素信号通路的调控,在植物的生长发育过程中起到关键调控作用。
生长素信号途径中,生长素和ARF(auxin response factor)转录因子均为植物生长素响应的关键调节因子[2]。ARFs转录因子可以与生长素响应基因启动子区域中生长素响应元件(AuxREs)TGTCTC/GAGACA发生特异性结合,激活或抑制下游基因的表达[3]。目前研究发现,植物中miR160和miR167通过靶向ARFs基因参与生长素信号转导调控,进而影响植物的生长发育。如在拟南芥中,miR167通过调控ARF6和ARF8的表达影响拟南芥雌蕊、雄蕊以及花瓣的发育[4]。miR160能够介导ARF17的转录后调控,从而影响植株对生长素的敏感性,进而调控植物体的发育[5]。此外,在其他植物中也已证实miR160及miR167家族的靶基因为ARFs,如番茄miR167下调ARF6和ARF8导致花的发育异常和雌性不育[6]。miR160a抑制slARF10影响番茄叶片的长出及早期果实的发育[7]。水稻中超量表达miR167导致水稻分蘖角度变大和植株矮小,表明miR167在调节植株株型上起作用[8]。在低温状态下,草莓miR167通过调节靶基因ARF8来调控激素的生物合成从而延缓草莓果实的衰老[9]。miR167靶向ARF8控制细胞的伸长参与大花牵牛(Ipomoeanil)幼苗茎尖,叶原基和雌蕊等器官的发育,并可能间接调控气孔的开放[10]。在杂柑品种‘清见’的果实发育过程中,伴随着IAA含量的变化,miR167和靶基因ARF8的表达量呈现出此消彼长的趋势[11]。综上,miR167靶向ARFs基因参与调控植物生长发育及生物胁迫等多个方面,表明二者在植物生长发育过程中具有十分重要的作用。
目前miR167的研究集中于拟南芥、水稻、番茄等模式植物中,且大多研究的重点为植株的生长发育调控,有关miR167参与生长素信号通路调控果实生长发育的研究还鲜有报道。本项目组前期从桃的基因组中预测到了miR167a及其对应的3个ARFs靶基因[12],本研究中以桃品种‘小白凤’不同发育时期的果实为实验材料,验证了该品种果实中ppe-miR167a的精确序列,克隆了预测的3个ARFs靶基因的ORF并进行了序列进化和保守性分析;qRT-PCR分析了miR167a及其靶基因在果实不同发育时期的空间表达情况;降解组测序确定了桃ppe-miR167a与ARF8间的裂解作用模式并明确了裂解位点;研究了ppe-miR167a、ARF8及其3′ 端裂解产物对外源IAA处理的响应,为今后进一步研究miRNA参与生长素信号途径调控桃果实发育的分子机制提供了理论支持。
1 材料和方法
1.1 植物材料
以苏州农业职业技术学院合作试验基地的‘小白凤’水蜜桃(Prunuspersica)成年树上不同发育时期的果实作为实验材料。于花后20 d (果实第一次迅速生长期)、50 d(果实硬核期)、75 d(果实第二次迅速生长期)和90 d(果实成熟期)分别取样。
生长素处理于2019年6月10日(成熟前30 d)用0.2 mmol·L-1外源IAA(加1% 吐温80)进行活体浸果处理1 min,清水(加1% 吐温80)为对照,处理后30 d取样。采集的所有样品带回实验室液氮速冻并放置在-80 ℃冰箱保存。
1.2 引物合成及生化试剂
实验所用引物由生工生物工程(上海)股份有限公司合成,具体序列见表1。大肠杆菌(Escherichiacoli)菌株DH5α、DNase酶Ⅰ、Power Script Ⅱ TM反转录酶、Taq酶、dNTPs、pMD19-T载体、DNA Marker、荧光定量染料SYBR Green Ⅰ均购自TaKaRa公司,DNA回收试剂盒购自北京天根生化科技有限公司。

表1 引物序列及用途
1.3 方 法
1.3.1 RNA提取及miRNA分离采用改良CTAB法提取不同发育阶段果实中的总RNA[12],DNase Ⅰ酶(RNase free)消化及氯仿抽提去除基因组DNA。在总RNA基础上,用4 mol·L-1LiCl分离出低分子量RNA(LMWRNA)与mRNA[12]。
1.3.2 cDNA合成mRNA与LMWRNA cDNA合成参考Zhang等[12-13]的方法。
1.3.3 桃miR167a精确序列验证以1.3.2中合成的LMWRNA的cDNA为模板,用表1中的引物进行扩增,miR-RACE方法参照Sun等进行[14]。
1.3.4 预测靶基因的克隆及序列分析根据从桃基因组中预测miR167a的3个靶基因序列[12],以所得总RNA反转录的cDNA为模板进行PCR扩增,获得‘小白凤’桃PpARF6、PpARF8 及PpARF6-like的ORF序列。获得的序列在NCBI中进行Blast(http/www.ncbi.nlm.nih.gov/BLAST/)比对。利用保守结构数据库(CDD, https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml)进行保守结构域的分析。系统进化树的构建使用MEGA 5.0软件。
1.3.5 降解组测序依据‘小白凤’桃确定的4个发育时期,选取花后20、50、75 和90 d果实的果肉部分进行降解组测序。总RNA 用Trizol试剂盒 (Invitrogen, CA, USA)提取,构建4个降解组文库,样品送联川生物公司(杭州)进行单末端(50 bp)测序,测序平台为Illumina Hiseq 2500(Illumina Inc., San Diego, CA, USA)。
1.3.6 实时荧光定量RT-PCR分别以mRNA、miRNA以及加5′端接头和3′-ploy(A)尾巴纯化的mRNA反转录合成的cDNA为模板,以桃RPII基因[15]和U6 rRNA为内参进行qRT-PCR扩增[16]。实验重复3次,实验数据用LinRegPCR和Excel 2010软件分析。目的基因的引物见表 1。
2 结果与分析
2.1 ppe-miR167a 精确序列验证及在不同发育时期桃果实中的表达
依据前期在桃基因组中预测的桃ppe-miR167a序列,利用miR-RACE技术验证ppe-miR167a在桃品种‘小白凤’中的精确序列[12]。实验结果表明,验证的ppe-miR167 序列与桃基因组中预测的序列在5′ 和3′ 端均存在一定的碱基差异,即验证序列在5′ 端第1个碱基处多了碱基U,而在3′ 端最后1个碱基处少了碱基G(表2)。
为进一步了解ppe-miR167a在桃果实发育过程中的表达情况,本试验利用qRT-PCR对其在‘小白凤’桃果实发育过程中4个关键时期的时空表达进行了分析[17]。结果表明,ppe-miR167a在花后20 d时表达量最高,随着果实进入硬核期,ppe-miR167a表达量显著下降,自硬核期(花后50 d)开始,ppe-miR167a的表达量逐渐上升(图1)。
2.2 ppe-miR167a预测靶基因的克隆及序列分析
依据前期利用psRNA target web server(http://plantgrn.noble.org/psRNATarget/)在基因组中预测的ppe-miR167a的靶基因结果(表3),为分析预测的靶基因,以mRNA反转录合成的cDNA为模板,利用在桃基因组数据库(https://www.rosaceae.org/species/prunus_persica/genome_v1.0)中比对到的序列设计引物,克隆得到PpARF6(XM_020562547)、PpARF8(XM_020560148)和PpARF6-like(XM_007217628)的完整ORF,大小分别为2 664、1 767 和 2 760 bp(图2)。保守结构域分析显示3个ARFs基因均含有高度保守的生长素响应DNA结合域(图3)。
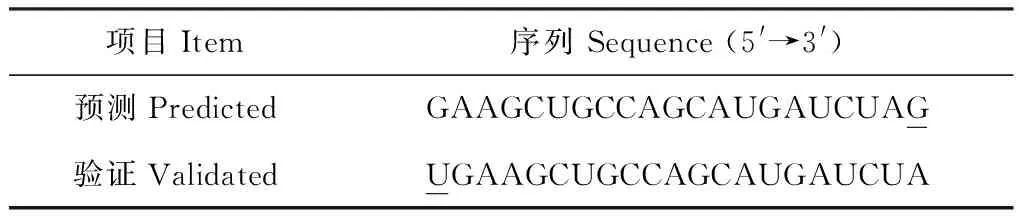
表2 桃miR167a预测序列与验证序列比较

不同小写字母代表显著差异(P<0.05),下同图1 桃果实不同发育时期ppe-miR167a的表达Different normal letters represent significant differences (P<0.05), the same as belowFig.1 Relative expression of ppe-miR167a in different stages of peach fruits
2.3 ppe-miR167a预测靶基因的系统发育
通过MEGA 5.0 软件构建了ppe-miR167a 3个预测靶基因的系统进化树(图4),结果显示,桃中PpARF6、PpARF8 及PpARF6-like均与巴旦杏的亲缘关系较近,其次为梅和甜樱桃。PpARF6和PpARF6-like与杨树的亲缘关系较远,而PpARF8与核桃的亲缘关系较远(图4)。
2.4 桃果实发育过程中ppe-miR167a预测靶基因的表达
分析ppe-miR167a预测靶基因在果实不同发育阶段中的表达结果表明,PpARF6在花后20和75 d果实中的表达基本一致,在硬核期(花后50 d)的表达相对较高,花后90 d果实中表达量最高(图5)。PpARF8的表达量在果实第一次快速膨大期(花后20 d)最高,至硬核期(花后50 d)有所下降,在随后的两个时期中无显著变化。PpARF6-like的表达量在4个发育时期无显著变化(图5)。分析ppe-miR167a与其预测靶基因的表达模式表明,PpARF8与ppe-miR167a的表达趋势基本一致,PpARF6和PpARF6-like的表达与其无明显相关性。因此,预测的3个靶基因及其与ppe-miR167a间的作用方式均有待进一步深入研究。
2.5 ppe-miR167a与其靶基因间的作用方式
为验证预测靶基因的真实性及ppe-miR167a与靶基因间的作用模式,本研究通过降解组测序进行了鉴定分析。结果表明,ppe-miR167a以剪切方式作用的靶基因有2个,其一为ARFs家族基因,即PpARF8,另一个为HPR-A基因(生物信息学未预测到,非ARF基因家族成员),其与miR167a间的作用频度较低,本研究中不进行详细分析。降解组未检测到ppe-miR167a与PpARF6和PpARF6-like之间的作用,说明信息学预测结果存在一定的误差,有必要进行深入的验证分析。降解组数据显示ppe-miR167a 作用于靶基因PpARF8的裂解位点位于miR167a 5′端的第9和10位碱基之间,即PpARF8 的2 441位碱基处(图6)。另外,在桃果实发育的4个时期,miR167a对PpARF8的裂解频度存在显著差异(表4),其中二者间的裂解频度在花后20和90 d时较低。在花后75 d时作用强度最大,表明miR167a可能在桃果实发育的第二次迅速生长期(花后75 d)起主要调节作用。

表3 预测的桃基因组中miR167a的靶基因

图2 桃PpARF6、PpARF8 及PpARF6-like的ORF扩增(M. DL5000)Fig.2 PCR products of PpARF6, PpARF8 and PpARF6-like genes ORF

图3 桃PpARF6、PpARF8 及PpARF6-like序列的保守结构域分布Fig.3 The distribution of conserved domains in PpARF6, PpARF8 and PpARF6-like genes
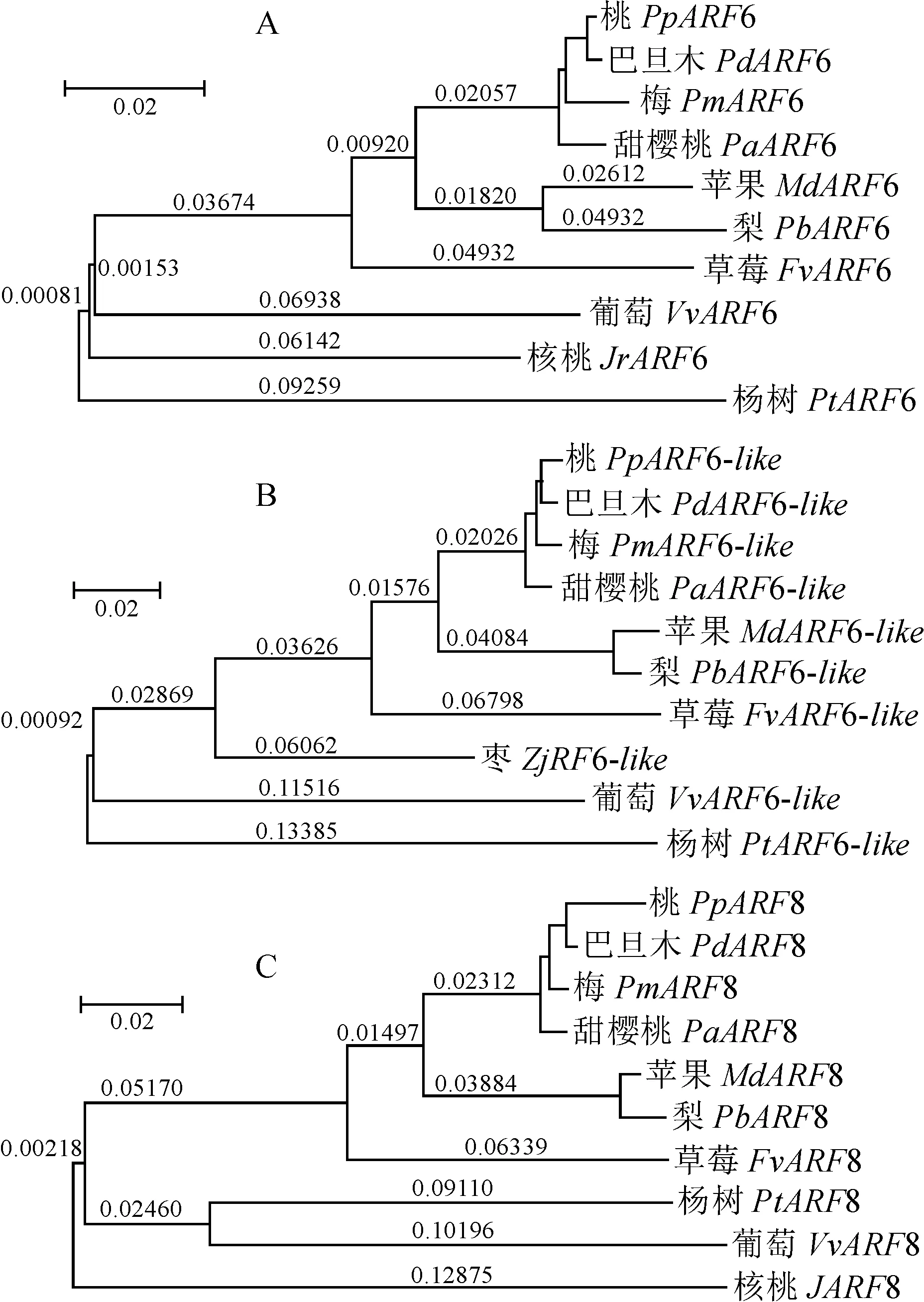
A. PpARF6;B. PpARF6-like;C. PpARF8图4 PpARF6、PpARF8 及PpARF6-like预测靶基因的进化树分析Fig. 4 Phylogenetic analysis of predicted target genes PpARF6, PpARF8 and PpARF6-like
2.6 生长素处理对桃ppe-miR167a、靶基因PpARF8 及裂解产物表达的影响
上述降解组结果表明ppe-miR167a的主效靶基因为生长素应答因子PpARF8,已知生长素通过与应答因子(ARF)结合影响植物的生长发育,因此为解析ppe-miR167a参与生长素信号调控果实发育的功能,本研究在果实发育后期用外源IAA处理30 d,对果实中ppe-miR167a及其靶基因的空间表达进行了分析,同时也分析了二者间裂解产物表达量的变化。结果表明,果实发育后期生长素处理后,处理组中ppe-miR167a的表达量低于对照组,而其靶基因PpARF8以及3′ 端裂解产物的表达在处理组中均高于对照组(图7)。由以上结果可以看出,生长素处理能在一定程度上影响ppe-miR167a及其靶基因PpARF8 的表达以及二者间的作用频度。

不同小写字母代表3个基因在不同发育时期表达量的显著性差异(P<0.05)图5 果实不同发育阶段PpARF6、PpARF8 及PpARF6-like的相对表达Different normal letters represent the significant differences of the expression levels of three genes in different development periods (P<0.05)Fig.5 Relative expression of PpARF6, PpARF8 and PpARF6-like in different stages of peach fruits

图6 ppe-miR167a与PpARF8间的作用方式及作用位点鉴定Fig.6 The mode and site of action between ppe-miR167a and PpARF8 were identified by degradome sequencing
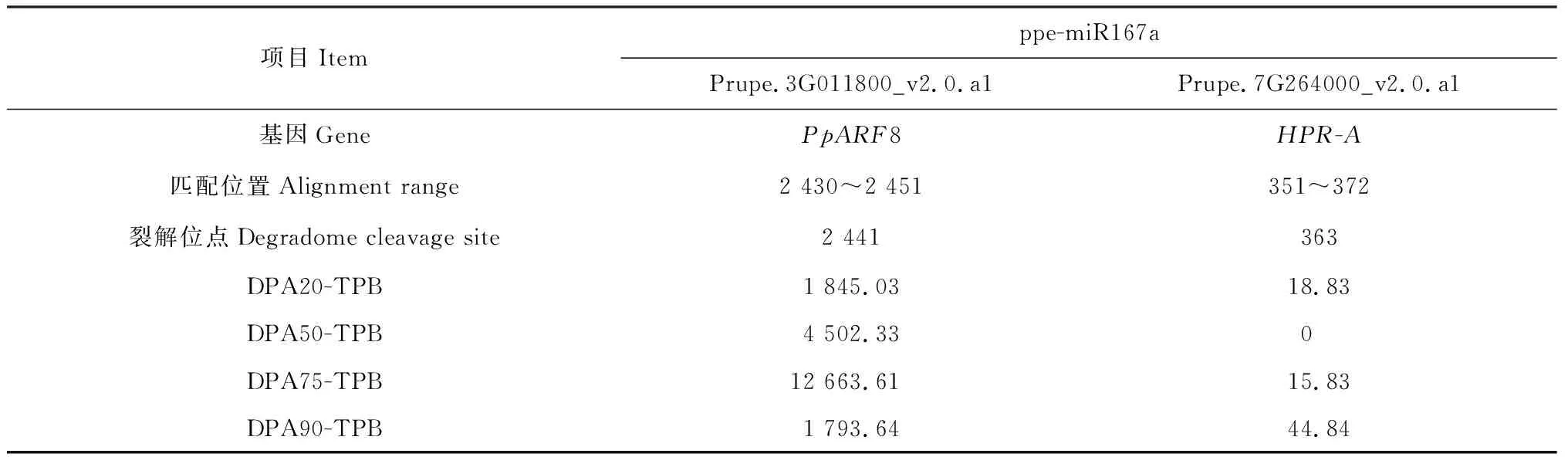
表4 降解组测序鉴定的ppe-miR167a的靶基因信息
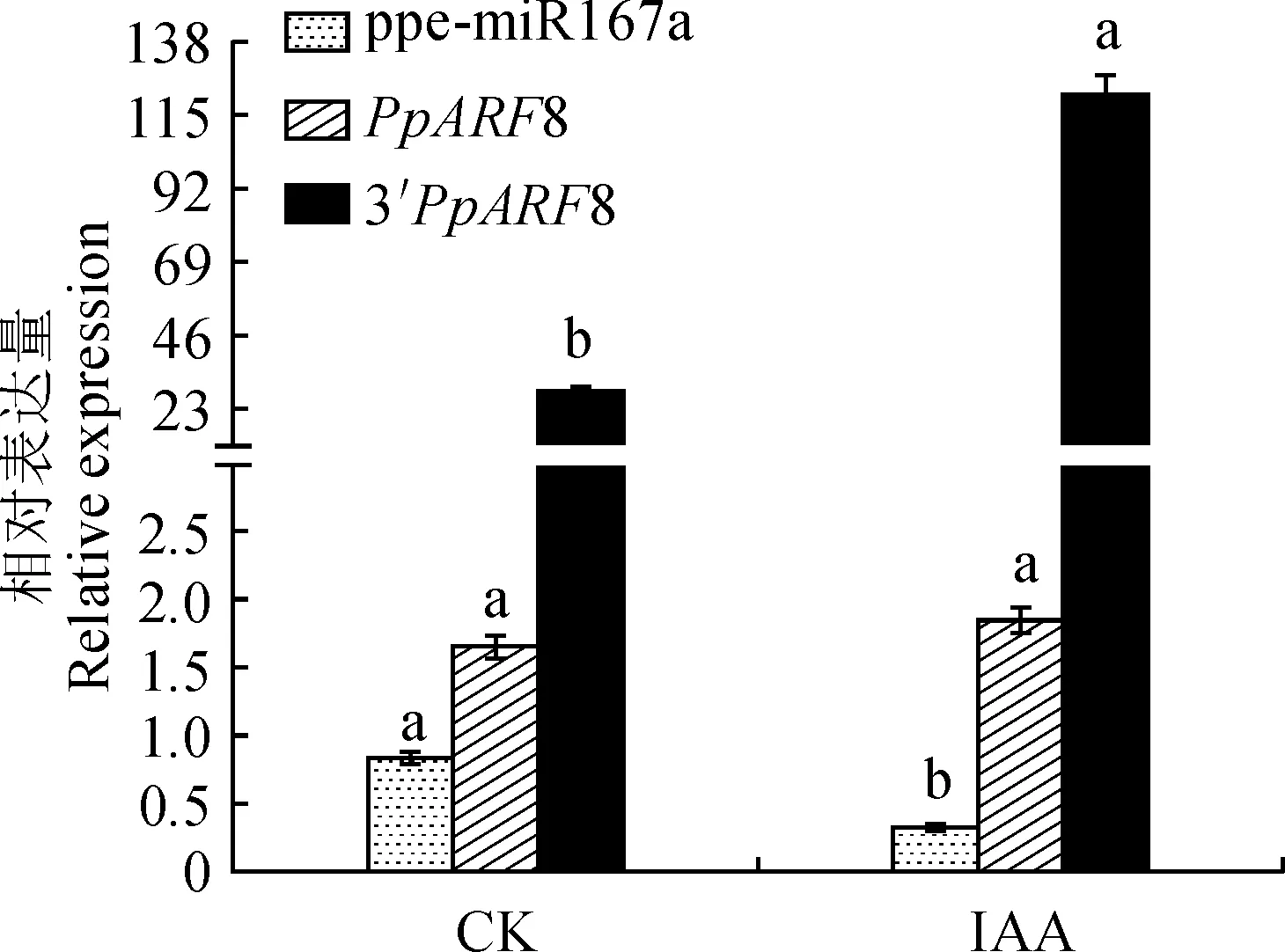
不同小写字母代表同一基因在处理前后的表达量的差异显著(P<0.05)图7 IAA处理后ppe-miR167a与其靶基因及靶基因3′ 端裂解产物的表达Different normal letters represent the significant differences of the expression levels of the same gene with treatments (P<0.05)Fig.7 Expression patterns of ppe-miR167a, ARF8 and cleavage product of peach ARF8 3′ end with IAA treatment and the control fruits
3 讨 论
目前的报道已经证明ARF转录因子为生长素信号通路中的关键因子,直接参与生长素信号,在植物果实成熟过程中发挥重要作用[18-21]。本项目组前期的研究发现ppe-miR167a的靶基因为生长素响应因子ARFs家族成员,为研究二者间的作用关系及它们在桃果实发育过程中可能存在的作用,我们首先验证了miR167a在‘小白凤’品种中的精确序列,结果表明,ppe-miR167a的序列在不同品种间存在一定的碱基差异[12],且差异碱基发生在序列的两端,这与早期其他植物中的研究结果一致[22]。分析ppe-miR167a在不同发育时期桃果实中的表达情况发现miR167a在果实快速膨大期和果实成熟期表达量较高,而在果实硬核期表达量较低,这与杂柑品种‘清见’中miR167a的表达量在果实发育前中期相对较高而后期较低有所不同[11]。3个ARFs基因的空间表达结果显示PpARF8与ppe-miR167a的表达趋势基本一致,PpARF6和PpARF6-like的表达与ppe-miR167a的表达无明显相关性,这与早期研究报道的miRNA与靶基因的表达一般呈负相关不同[23]。拟南芥中的研究发现,miR167与其靶基因AtARF6和AtARF8间存在反馈调节作用,AtARF6能激活miR167表达,AtARF8抑制miR167表达,它们之间通过彼此激活或抑制作用来调控不定根的形成[24]。在甜橙果实成熟过程中即花后170~250 d,CiARF8的表达量明显下调,在花后250 d的表达量保持在一个较低的水平[25],这与本研究中PpARF8在桃不同发育时期果实中的表达也存在一定差异。
为验证预测靶基因的真实性及ppe-miR167a与ARFs基因间的作用模式,我们通过降解组测序进行了分析,结果表明仅PpARF8上存在miR167a的裂解位点,说明生物信息学预测结果存在一定的误差,可能是预测时参数设置问题导致了一定的假阳性结果。降解组结果显示ppe-miR167a 作用于靶基因PpARF8的裂解位点位于miR167a 5′ 端的第9和10位碱基之间,这与早期报道的miRNA裂解靶基因的位点大多位于miRNA 5′ 端的第10和11个碱基之间有所不同[26-27],说明miRNA对其靶基因的裂解位点既有一定的保守性,又存在一定的多样性[28-30]。降解组结果显示3′端裂解产物的表达与对应ppe-miR167a的表达趋势不一致,这与之前研究者在桃及其他物种中推测的miRNA表达量可能与靶基因表达趋势相反而与3′端裂解产物的表达趋势一致的情况存在差异[13, 31],这可能是miRNA在转录和转录后水平参与了植物发育的多个过程,其不仅对靶向的mRNA起作用,同时也可以和其他的RNAs相互作用调控其自身或其他的竞争性内源RNA[32-33]。
已有的报道表明AtARF4,AtARF5,AtARF16,AtARF19,OsARF1 和OsARF23 在生长素处理后,转录物的表达量有所增加[34-37]。FvARF响应生长素处理[38],SlARF2 转录物积累量明显受生长素和赤霉素的正调控[39]。目前,研究最多的生长素是IAA[40],但miR167靶向ARFs基因对IAA处理的响应分析还未见报道。王少希等通过外源IAA处理[21],研究了不同浓度和不同IAA处理时间对FvARFs表达的影响,结果表明外源 IAA处理后所有FvARFs的表达均增加,并在处理6 h或12 h达到峰值,但其未对miR167的表达进行分析。本研究中外源IAA处理桃果实30 d后定量表达结果显示,miR167a的表达量在处理组中低于对照组,PpARF8及3′ 端裂解产物的表达在处理组中均高于对照组,PpARF8在IAA处理后的表达变化情况与王少希等[21]的研究结果一致。由此可知,外源生长素处理能在一定程度上影响了miR167a和PpARF8 的表达以及二者间的裂解作用频度,但未表现出之前研究者推测的miRNA表达量可能与靶基因表达趋势相反而与3′ 端裂解产物的表达趋势一致的现象,这一情况推测可能是miRNA和靶基因在植物的生长发育中不止在一个代谢过程中起作用,它们可能参与了植物生长发育的多个方面,受多种因素调控。综上,桃果实中ppe-miR167a可以通过介导其靶基因PpARF8的裂解影响生长素信号途径参与果实发育的调控,但其调控机理及作用通路还有待进一步深入研究。
