草-溞生物组合对农田周边富营养化水体中浮游植物的影响
2021-03-09张春雪魏孝承王晗阳郑向群
张春雪 ,魏孝承 ,王晗阳 ,2,郑向群 ,杨 波
(1.农业农村部环境保护科研监测所,天津 300191;2.天津城建大学,天津 300384)
近年来,中国水环境污染加剧,水体富营养化现象已成为一个不容忽视的环境问题之一[1],由此引发的藻类水华频发等现象是水体富营养化的主要表现形式[2]。其中,蓝藻水华在影响水生生态系统协调与稳定性的同时,其次生代谢产物藻毒素对其他水生植物、动物甚至人类的健康产生巨大影响[3]。生物修复富营养化水体的方法因其经济性和环境友好性成为当前国内外研究的热点。在以往的研究中,生物操纵对蓝藻水华的抑制均取得了一定成效,但在一些试验和应用中并未得到预期效果[4,5]。究其原因,一方面,水生动物排泄会使营养盐上升,且其对藻毒素的耐性仍存在争议;另一方面,部分水生动物会吞食浮游动物,对控制藻类生长产生负面影响。因此,进一步开发联合修复技术,降低负面影响,成为现阶段亟需解决的问题。
近年来,在生态修复实践中,采用2 种或2 种以上水生生物联合修复富营养化水体的技术受到广泛关注。Zhou 等[6]采用菹草(Potamogeton crispus)进行模拟修复试验,结果表明,在种植菹草的试验水体中总氮(TN)、总磷(TP)、叶绿素含量等指标均发生大幅度下降,并且在试验结束时菹草可改变浮游植物的群落结构,使之由绿藻为主,变为蓝藻、绿藻以及底栖藻类共存的良好状态。Zhao 等[7]在实验室可控条件下,提取金鱼藻的化感物质,通过提取液和普通小球藻(Chlorella vulgaris)的共培养试验证明,金鱼藻提取液不仅能够显著抑制小球藻的生长,并且对小球藻的形态也有影响,加入金鱼藻提取液的培养组内小球藻细胞呈现松散、破裂的情况。Zhang 等[8]选用苦草(Vallisneria natans)、金鱼藻(Ceratophyllum demersum)与绿藻门的刚毛藻(Cladophora)进行共培养试验,结果表明,苦草和金鱼藻这2 种沉水植物对刚毛藻的生长具有显著抑制效果。李启升等[9]采用室内可控试验,研究不同氮负荷对苦草和狐尾藻(Myriophyllum aquaticum)生长的影响,每日将含N 25 μg/L 和 500 μg/L 的营养液输入试验水体,以此来模拟高、低N 负荷,结果表明,在高、低N 负荷下,苦草和狐尾藻的存在均可以明显降低水中氨氮、硝氮浓度,并且高N 负荷对于狐尾藻的株高、生长率等指标均无显著影响,表明二者在较高的N 负荷下依旧可以发挥净水作用,并且狐尾藻表现出较好的N 耐性,说明其可以应用在较高N 浓度的水体修复中。邢春玉等[10]选用荇菜(Nymphoides peltatum)、苦草、黑藻(Hydrilla verticillata)和穗花狐尾藻(Myriophyl⁃lum spicatum)4 种天津市较为常见的水生植物对处于蓝藻暴发期的海河水体进行处理,结果表明,4 种植物均对蓝藻有很好的抑制效果,且其对水体pH、溶解氧以及温度变化具有良好的调节作用。现有研究中对生物操纵已有一定的成果[11-13],但对于生物操纵是否能够有效抑制产毒蓝藻,滤食性动物在修复水体时的添加密度是否会影响其对水华的抑制作用,水生植物修复和生物操纵能否完全相辅相成,此类生物修复过程中对水体浮游细菌群落结构会造成的影响等问题,仍需进一步讨论和研究。本研究基于生物操纵理论和水生植物修复理论,结合目标水体现状,采用大型溞作为滤食性动物的代表,狐尾藻和金鱼藻作为沉水植物的代表,在实验室可控条件下,模拟春夏水华易发生时期的自然状态,设计9 个试验处理,研究单一生物及其组合对富营养化水体的修复作用,并对其修复机理进行初步分析。
1 材料与方法
1.1 材料来源
供试大型溞购自中国科学院沈阳应用生态研究所;供试植物为狐尾藻和金鱼藻,购自天津市某花鸟鱼虫市场;富营养化水体取自天津市宁河区某农田周边的池塘。
水体初始水质为pH 8.77、溶解氧(DO)9.28 mg/L、透明度17.20 cm、TN 7.38 mg/L、TP 0.31 mg/L、NH4+-N 0.21 mg/L、硝氮(NO3--N)6.59 mg/L、叶绿素 a 68.59μg/L、浮游植物密度 2.80×107cells/L、蓝藻门密度2.52×107cells/L、微囊藻毒素2.77 μg/L。
1.2 试验设计
试验设9个处理,分别为对照(CK);高(100 ind/L)、低(25 ind/L)2 种大型溞密度处理,研究不同密度大型溞对试验水体的影响;2 种沉水植物金鱼藻、狐尾藻处理,研究不同沉水植物对试验水体的影响;2 种沉水植物和2 种密度大型溞分别组合处理,研究草-溞联合对试验水体的影响,详见表1。每处理均设3个重复,容器体积为20 L,试验时间为20 d。
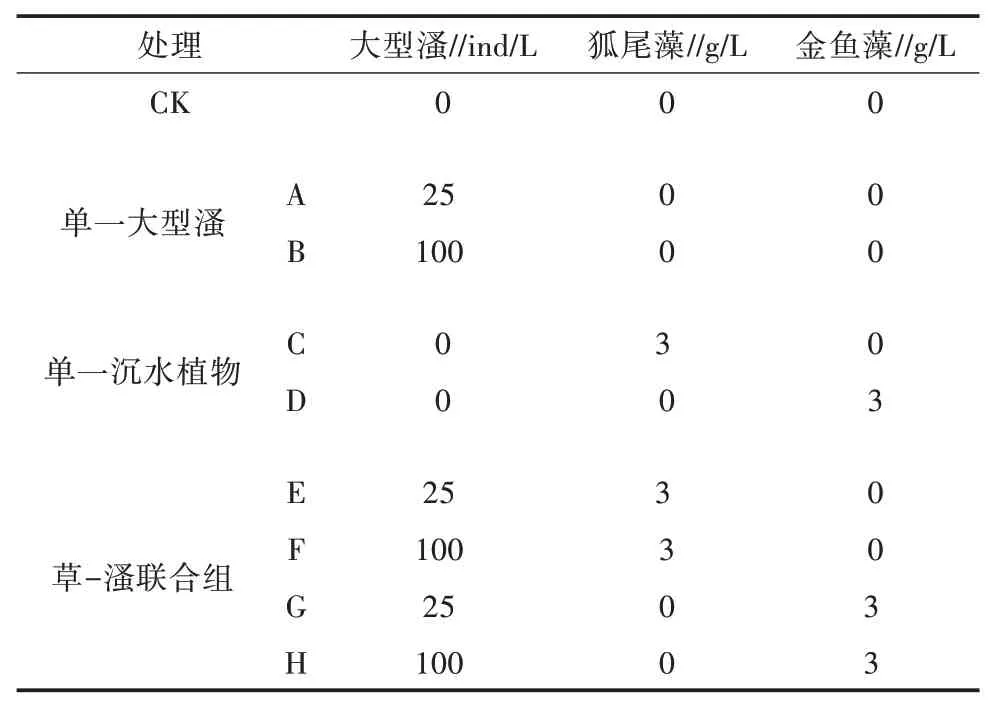
表1 试验设置
1.3 样品的采集
在试验2、5、10、15、20 d 时进行水样采集,采样时间为上午8:00—9:00。为减少试验误差,在每次取样后记录水位,在下次取样前采用去离子水补足蒸发量[14]。
1.4 样品的测定
1)透明度。采用铅字法测定。
2)浮游植物密度。移液枪吸取浓缩后的浮游植物样品1 mL,采用0.1 mL 浮游植物计数框于奥林巴斯显微镜400×下定量计数,每个样品计数3 片后取平均值,浮游植物种类鉴定参考《中国淡水藻类:系统、分类及生态》[15],浮游植物的密度参考章宗涉等[16]的研究方法,具体计算公式如下。

式中,N为每升样品中的浮游植物数量,V为采样体积,Vs为原水样浓缩后的体积,n为镜检计数所得的浮游植物数目,Va为计数框容量。
3)叶绿素 a 含量。采用丙酮法萃取[17]测定水体中的叶绿素a(chla),计算公式如下。
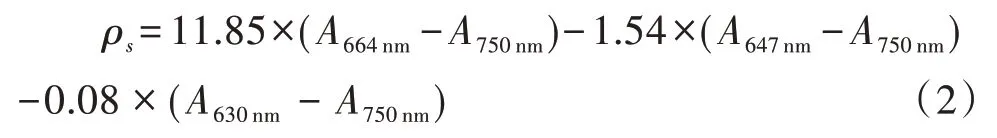
式中,ρs为样品中 chla 的质量浓度,A为吸光度。

式中,ρw为试验水体中叶绿素a 的质量浓度,ρs为样品中叶绿素a 的质量浓度,V1为离心管中溶液的体积,V为水样的体积。
4)微囊藻毒素。采用酶联免疫法(ELISA)检测[18],同一样本测定3 个平行样后取平均值。
5)大型溞密度。使用玻璃棒搅拌混匀水体,取1 L 水样采用3%的甲醛溶液固定并浓缩至50 mL,采用1 mL 计数框于显微镜100×下定量计数,每个样品均计数3 次后取平均值。
6)植物湿重。将各试验桶中的水生植物取出,用滤纸吸干表面的水分后,采用电子天平称重,每个样品称3 次取平均值。
1.5 数据统计分析
各项指标采用Origin 9.0制图,数据均以3个平行试验桶的平均值±标准差形式表示。采用SPSS 24.0进行统计分析,通过单因素方差分析(one-way/ANOVA)检验指标的差异显著性,图基检验(HSDtest)用于单因素方差分析后的多重比较,显著水平为0.05。
2 结果与分析
2.1 不同密度大型溞对水体透明度、浮游植物的影响
试验期间,不同密度大型溞对水体透明度的影响见图1。结果显示,试验2 d 时,各处理透明度差异较小;5~20 d 时,各试验处理水体透明度表现为CK
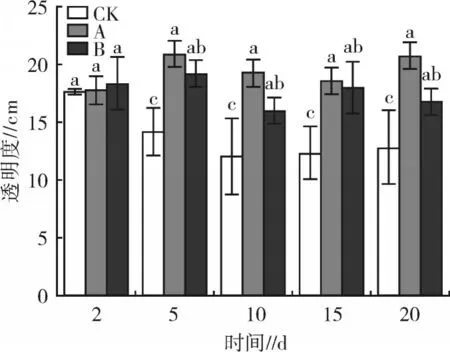
图1 不同密度大型溞对水体透明度的影响
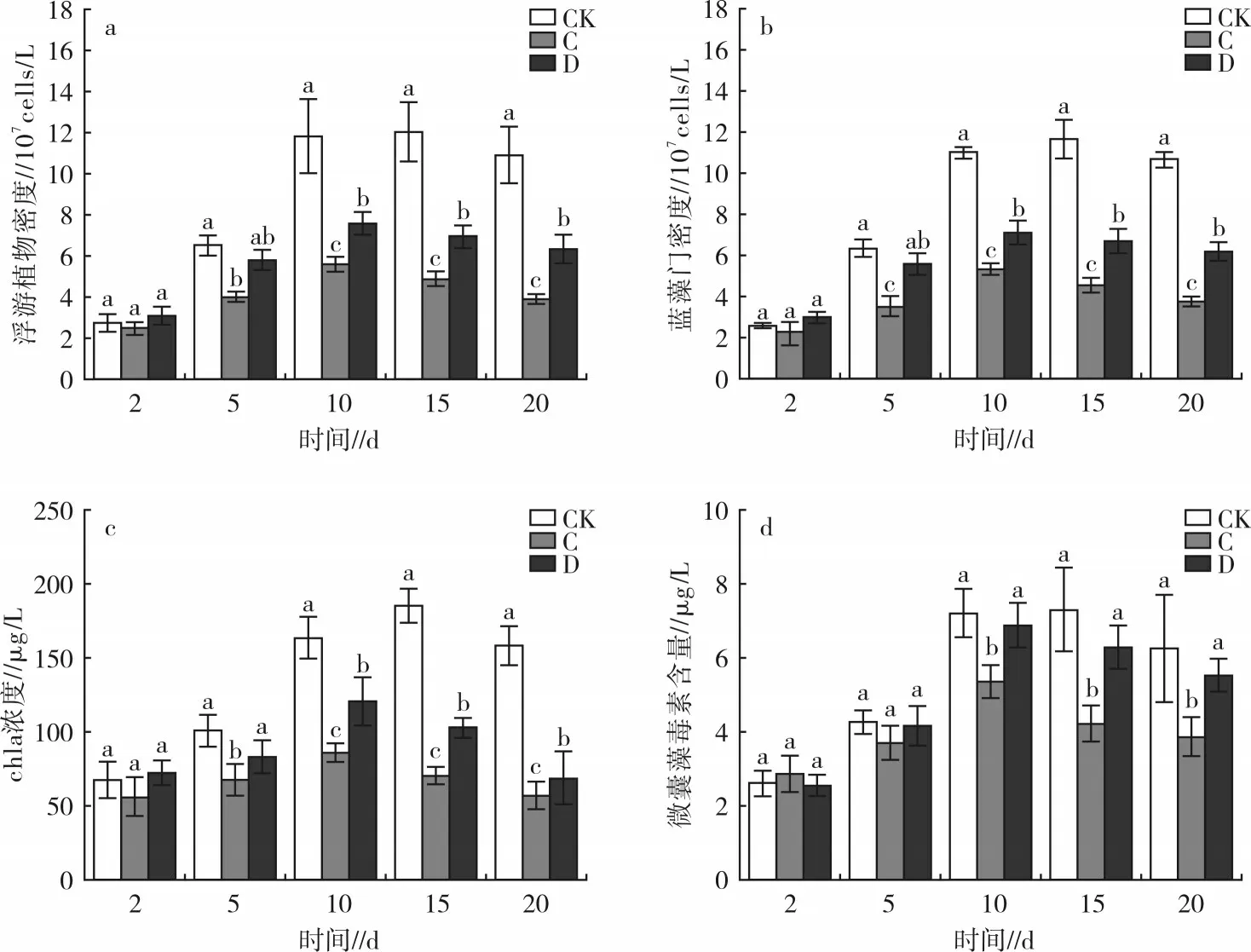
试验期间,各处理中的浮游植物密度、蓝藻门密度、chla 浓度和微囊藻毒素含量的变化趋势见图2。结果显示,在试验 5~20 d 后,处理 A、B 中的浮游植物密度、蓝藻门密度、chla 浓度均显著低于CK(P<0.05);在 10 d 时,处理 A 中的这 3 项指标均显著低于处理B(P<0.05);随着时间推移,3 项指标在处理A、B 之间的差异缩小,试验结束(20 d)时,处理 A 中的3 项指标分别为 3.24×107cells/L、3.10×107cells/L 和42.51 μg/L,处理 B 中的 3 项指标分别为 3.81×107cells/L、3.67×107cells/L 和 54.92 μg/L,均较 15 d 时的数据有所反升,此时处理A、B 间无显著差异(P>0.05)。微囊藻毒素含量在CK 中呈先升高后降低的趋势,在处理A、B 中则呈先升高后降低再升高的趋势。试验5~15 d 时,处理B 中的微囊藻毒素含量显著低于CK(P<0.05),但随时间推移,处理B 中的微囊藻毒素含量逐渐上升,试验结束(20 d)时,处理B中的微囊藻毒素含量升至5.56 μg/L,CK 中的含量为6.27 μg/L,此时处理 B 与 CK 无显著差异(P>0.05);试验15~20 d 时,处理A 中的微囊藻毒素含量虽出现反升,但一直显著低于CK(P<0.05)。
2.2 2 种沉水植物对水体透明度、浮游植物的影响
试验期间,2 种沉水植物对水体透明度的影响见图3。处理C、D 的透明度均高于CK,但除了试验20 d 时,处理C 的水体透明度显著高于CK 以外(P<0.05),其余时间点差异均不显著(P>0.05)。研究表明,沉水植物可以通过抑制浮游植物生长,提高水中透明度,但沉水植物的生长一般需要较长的时间[20],本试验周期较短,因而2 种沉水植物对透明度的提升作用并不显著。
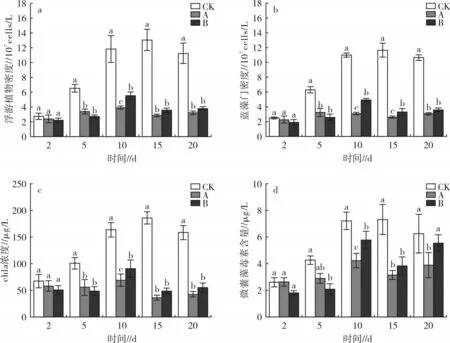
图2 不同密度大型溞对浮游植物的影响
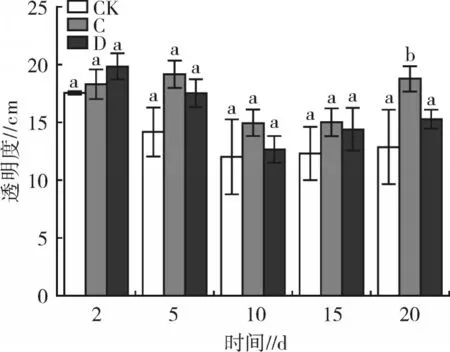
图3 2 种沉水植物对水体透明度的影响
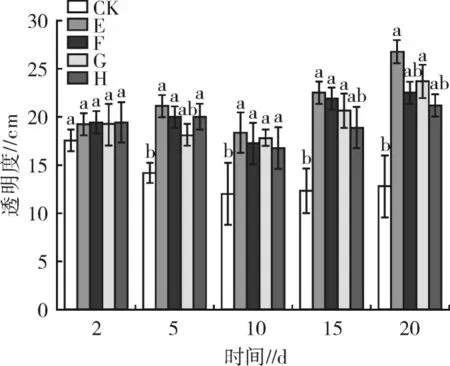
浮游植物密度、蓝藻门密度、chla 浓度在各处理中的变化趋同(图4),在试验5 d 后,处理C、D 中上述指标均显著低于 CK 中的含量(P<0.05),5 d 时,处理C、D 之间浮游植物密度差异不明显,5 d 之后处理C 中浮游植物密度、蓝藻门密度、chla 浓度显著低于处理D 和CK(P<0.05)。试验结束(20 d)时,处理C、D 和CK 中浮游植物密度、蓝藻门密度、chla浓度分别表现为 C(3.93×107cells/L) 微囊藻毒素含量在处理C、D 和CK 中的含量均呈先增高后下降的趋势。试验2~5 d 时各处理微囊藻毒素含量差异不显著(P>0.05),5 d 之后,微囊藻毒素在处理C 中的含量显著低于处理D 和CK(P<0.05),处理 D 中的含量则与 CK 无显著差异(P>0.05)。试验结束(20 d)时,微囊藻毒素在处理C、D和CK 中的平均含量由小到大依次为 C(3.88 μg/L)、D(5.55 μg/L)、CK(6.27 μg/L)。这说明狐尾藻能显著抑制微囊藻毒素含量的上升,而金鱼藻对其无显著抑制作用。 试验期间,草-溞联合对水体透明度的影响见图5。试验2 d 时,各处理水体的透明度差异均不显著(P>0.05)。试验 5~20 d 时,各草-溞联合处理水体的透明度处理E 最高,且4 个草-溞联合处理均高于CK。这是由于金鱼藻、狐尾藻对浮游植物有抑制作用,大型溞对浮游植物有摄食作用,在二者的协同作用下,使水体透明度显著提升。 浮游植物密度、蓝藻门密度、chla 浓度在各草-溞联合处理中的变化趋同,见图6。试验2 d 时,3项指标在各处理中差异不显著(P>0.05)。在试验5~20 d,各处理的浮游植物密度、蓝藻门密度、chla 浓度均显著低于CK(P<0.05)。各处理中3 项指标均呈先升高后降低的趋势,试验结束(20 d)时,各草-溞联合处理中浮游植物密度、蓝藻门密度、chla 浓度分别为 E(0.92×107cells/L) 图4 2 种沉水植物对浮游植物的影响 图5 草-溞联合对水体透明度的影响 试验期间,各草-溞联合处理和CK 中的微囊藻毒素含量变化趋同,均呈先升高后下降的趋势。在试验2~5 d 时,各处理中微囊藻毒素含量差异不显著(P>0.05)。随着时间的推移,在试验10 d 时,各处理中微囊藻毒素含量均显著低于CK(P<0.05),且处理E 中的微囊藻毒素含量显著低于其他处理(P<0.05)。15~20 d 时,处理 E、F、G 中的微囊藻毒素含量显著低于处理H(P<0.05)。试验结束(20 d)时,各草-溞联合处理微囊藻毒素含量表现为E(0.78 μg/L) 分别对比处理E 和G、处理F 和H,可以发现狐尾藻与大型溞联合对浮游植物相关指标的抑制作用均优于金鱼藻。在单一沉水植物试验(处理C、D)结果中,发现狐尾藻对浮游植物的抑制效果优于金鱼藻,这在草-溞联合修复的试验结果中也得到了进一步的印证,说明在草-溞联合修复时,植物的类别会影响其对浮游植物的抑制作用。在之前的结果中,单一使用金鱼藻对微囊藻毒素几乎没有抑制作用,但将其与大型溞联合使用时,微囊藻毒素均被显著抑制;对比单一使用狐尾藻的试验结果,发现大型溞与其联合处理对浮游植物的抑制效果也得到进一步提升。这一结果与姜小玉等[21]的研究一致,当沉水植物与大型溞联合修复水体时,对浮游植物的抑制效果均优于单一使用沉水植物。这是由于大型溞的摄食作用可以迅速提升水体的透明度,使水体恢复成清水态,改善光照条件,加速沉水植物的生长。试验结束后,处理E、F 中狐尾藻的平均湿重为(4.23±0.37)g/L;处理G、H 中金鱼藻的平均湿重为(3.54±0.62)g/L,均较之前单一使用时有所增加,这也证实了大型溞对沉水植物的生长促进作用。 大型溞属于植食性甲壳类动物,可以大量摄食水中浮游植物,从而降低水中浮游植物的含量;另一方面,其排泄作用加速营养盐的循环,促进浮游植物的生长[22]。在试验初期(2~5 d),高密度处理摄食量大,因而对浮游植物抑制作用较强,但在试验5~20 d时,高密度处理的排泄作用更大,加速了营养盐的循环作用,浮游植物相关指标较低密度处理略高。试验后期(15~20 d),大型溞排泄作用高于摄食作用,致使20 d 时处理A、B 中的浮游植物密度较15 d 反升。微囊藻毒素作为蓝藻的代谢产物,其含量与水体中的蓝藻门密度有关,试验后期,高、低密度处理的微囊藻毒素含量随蓝藻门密度的升高而升高。研究表明,微囊藻毒素一般可以在水生动物体内累积[23],但大型溞机体较小,耐性较差。此外,蓝藻毒素会对大型溞的生长和繁殖产生负面影响,且毒性的强弱与藻细胞密度呈正相关[24],本试验蓝藻门密度较高(>107cells/L),因而对大型溞的负面影响较大,试验期间处理A、B 水体中均观察到死亡现象。试验结束(20 d)时,处理A、B 的大型溞密度分别为(12.7±5.5)ind/L 和(41.8±13.7)ind/L,较添加密度25 ind/L 和100 ind/L 均发生大幅度下降,且高密度处理因受到较大种群竞争作用影响,下降更为明显。大型溞死亡时,体内储存的微囊藻毒素会再次被释放到水体中,因而试验后期大型溞对微囊藻毒素的抑制作用减弱,致使其含量升高。 沉水植物可以通过吸收水中营养盐来抑制浮游植物的生长,狐尾藻和金鱼藻均可显著降低水中营养盐含量,因此二者对于浮游植物也有一定的抑制作用,狐尾藻组的营养盐下降较金鱼藻更为明显,这也是其对浮游植物的抑制作用更强的原因之一。除了吸收营养盐以外,沉水植物还可通过竞争光照、生存空间、释放化感物质等方式抑制浮游植物的生长[25],试验结束时狐尾藻长势较好,因此其对浮游植物的抑制作用更强。在本试验结果中,金鱼藻对微囊藻毒素无显著抑制作用,说明金鱼藻对微囊藻毒素的富集能力较差。一方面,金鱼藻在试验过程中对浮游植物以及蓝藻门密度抑制能力较狐尾藻弱,另一方面,蓝藻细胞破碎、死亡时也会释放大量的微囊藻毒素[26]。张之浩[27]的研究表明,金鱼藻可以释放α-甲基苯乙醇、己二酸二酯等化感物质,此类化感物质会破坏藻细胞膜使之破碎,破碎的藻细胞继续释放微囊藻毒素,但本试验中金鱼藻生长较慢,对微囊藻毒素富集能力差,这可能是金鱼藻在抑制蓝藻门密度的同时却对微囊藻毒素的抑制作用较差的原因。 草-溞联合均能够显著抑制水中浮游植物密度、蓝藻门密度、chla 浓度和微囊藻毒素含量的上升,其中以低密度大型溞+狐尾藻组合较优,试验结束时相比于对照,可分别降低91.58%、92.05%、91.82%、87.56%。低密度大型溞与沉水植物的联合对浮游植物的抑制作用均优于高密度大型溞与沉水植物的联合,这与单一使用大型溞的试验结果一致。试验结束后,处理E、G 的大型溞平均密度分别为(56.3±18.2)ind/L 和(38.3±7.9)ind/L,均较投加密度(25 ind/L)有所增加。分析其原因认为沉水植物可以为大型溞提供庇护空间,减轻其种间竞争作用,有利于其生长和繁殖,进而提升其对浮游植物的摄食作用。此外,低密度大型溞种间竞争较弱,为多营孤雌生殖,这种状态的大型溞繁殖速度快、摄食率高,因而对浮游植物的抑制作用更强。相反,试验结束后,处理F、H 中大型溞的平均密度分别为(63.5±21.7)ind/L和(53.8±27.5)ind/L,均较投加密度(100 ind/L)有所下降。这是由于大型溞密度过高时种间竞争作用过大,大型溞进入两性繁殖状态,试验期间在处理E、F水体中均观察到大量带有冬卵的大型溞个体,这种状态的大型溞摄食率极低,对浮游植物的抑制作用被削弱,此外高密度大型溞排泄作用高,加速N、P 营养盐的循环作用,促进了浮游植物的生长。因而在草-溞联合修复时,低密度大型溞+沉水植物的抑制作用更优。结合单一修复中的数据进行分析,试验结束时,在相同溞密度和植物类型的条件下,联合修复相比单一使用溞或植物对浮游植物的抑制效果更优,在此也可与透明度的变化趋势相印证。 近年来,水体富营养化现象日趋严重,已成为亟需解决的环境问题之一。本研究基于生物操纵理论和水生植物修复理论,结合目标水体现状,采用大型溞作为滤食性动物的代表,狐尾藻和金鱼藻作为沉水植物的代表,在实验室可控条件下,模拟春夏水华易发生时期的光照和温度条件,考察单一生物及其组合对富营养化水体的修复效果,为生物操纵和水生植物修复富营养化水体的应用提供数据支持。本研究结论如下。 1)大型溞对浮游植物密度、蓝藻门密度、chla 浓度及微囊藻毒素抑制显著,且低密度(25 ind/L)抑制作用更强。试验后期(15~20 d)抑制作用减弱,试验结束(20 d)时,高密度(100 ind/L)下微囊藻毒素含量与对照无显著差异。 2)狐尾藻和金鱼藻均能抑制浮游植物密度、蓝藻门密度和chla 浓度的上升,且狐尾藻抑制效果更优,而金鱼藻对微囊藻毒素含量无显著抑制作用。 3)试验结束时,在相同溞密度和植物种类条件下,草-溞联合较单一大型溞和单一植物抑制效果更强,且以低密度大型溞(25 ind/L)+狐尾藻最优,草-溞联合相比单一溞处理,可有效改善其在处理中产生的微囊藻毒素后期反升现象。2.3 草-溞联合对水体透明度、浮游植物的影响
3 讨论
4 结论
